Fragmentation forestière
La fragmentation des forêts désigne toutes les formes de fragmentation (physique et écologique) des habitats naturels forestiers (et d'écosystèmes associés le cas échéant).
Le morcellement anthropique des forêts augmente depuis plusieurs siècles. Il s'est fortement accru depuis quelques décennies, notamment dans les régions à forte démographie[1], en grande partie en raison de la croissance du maillage routier[2]. Il est devenu l'un des premiers facteurs de dégradation de la biodiversité[3], et en particulier de la biodiversité forestière. C'est l'une des premières menaces pour les grands écosystèmes forestiers, dont en forêt tropicale[4], en Amazonie particulièrement[5],[6],[7]. Les zones boréales[8], fraiches ou tempérées sont également très touchées[9], dont notamment les forêts des États-Unis[10]. Des facteurs climatiques peuvent l'aggraver[11],[12] ce qui est préoccupant pour les puits de carbone forestiers.





Cet article traite du morcellement physique et fonctionnel des forêts, qui explique certaines caractéristiques écologiques de milieux fragmentés, forestiers ou semi-forestiers (voir forêts urbaines, forêt périurbaine, grands parcs urbains boisés, forêts récréatives, bosquets et certains jardins…). Il traite aussi de ses causes[13] et de ses effets, ainsi que des moyens de les mesurer ou d'en réparer certains impacts.
Éléments de définition
Histoire de la notion
Le morcellement des forêts est une préoccupation récente[14], comme le sont les études sur ce thème. Leur nombre démontre un intérêt croissant pour le sujet (par exemple, quelque 2 500 études scientifiques ont été publiées sur le sujet en 2007 et concernant des forêts de presque tous les pays[15]). La fragmentation écosystémique est un vaste champ de recherche, qui aborde diverses échelles spatiales et temporelles, et un grand nombre de taxons.
Plusieurs raisons expliquent cet intérêt :
- s'agissant des surfaces, l'aggravation rapide du morcellement et de la destruction de la plupart des forêts tropicales aux XXe et XXIe siècles et la pression toujours exercée sur les autres surfaces forestières ;
- s'agissant de la biodiversité, les effets de la fragmentation ajoutés à ceux de la destruction des forêts primaires ou des secondaires[16] ;
- depuis le Sommet de la Terre de Rio (1992), la fragmentation (physique ou immatérielle) des habitats est reconnue comme une des grandes menaces pour l'ensemble de la biodiversité ;
- la fragmentation nuit et peut aggraver les questions relatives à l'adaptation des espèces et des écosystèmes au réchauffement climatique,
- certains effets n'avaient pas été anticipés comme les impacts différés des routes forestières sur la biodiversité.
Les études concernent les causes et effets de cette fragmentation[13] et parfois sur la modélisation du phénomène et/ou sur la mesure du morcellement, ou encore sur les moyens d'en atténuer, réparer ou compenser certains effets négatifs.
Définitions et vocabulaire
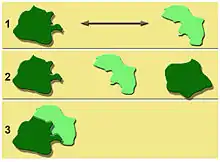
La longueur de la flèche symbolise la distance maximale que peut parcourir une espèce particulière.
1 : colonisation impossible
2 : colonisation possible à partir de deux massifs anciens
3 : colonisation par contact ; agrandissement de la forêt intérieure (le territoire où l'espèce est présente sera plus réduit que dans la situation 2)
La qualité de la matrice (en jaune) a une grande importante pour la dispersion d'une tache à l'autre[17]


La fragmentation forestière désigne le phénomène de morcellement qui transforme de grands massifs boisés en ensembles de parcelles plus petites et plus ou moins isolées, ce qui modifie le fonctionnement de leurs communautés microbiennes, fongiques, animales et végétales.
- Les parcelles sont appelées fragments ou patchs ou vestiges de forêt. La matrice paysagère (matrice écopaysagère) est l'espace non forestier qui entoure et isole les parcelles encore arborées ; il peut s'agir d'espaces naturels ouverts (savanes, prairies, pelouses, zones humides…) ou dans le cas de fragmentation d'origine anthropique (créée par l'homme) de terres agricoles, de zones urbanisées (zones d'activité commerciales, infrastructures de transport, etc.).
- La forêt intérieure ou forêt profonde est la partie de la forêt où les effets de bordure ou de lisière ne se font pas ou peu sentir. C'est une notion consensuelle[18] dont les critères de définition fondés sur l'importance de l'effet-lisière, sont encore en débat[19].
Par exemple, dans la littérature scientifique, au Sud Québec[20], en considérant les oiseaux forestiers, on ne peut parler de forêt continue qu'à partir de 1 000 hectares de forêt profonde d'un seul tenant, alors que Porneluzi et Faaborg (1999)[21] considèrent qu'il n'y a plus de forêt intérieure en dessous de 2 000 ha d'un seul tenant, Kupfer et al. (2004) jugent nécessaire de conserver des patchs d'au moins 2 300 ha non fragmentés que Faaborg et al[22]. portent à 3 000 ha au minimum, sans nier l'importance des petites réserves. Faaborg et al. (1993)[22] estiment même qu'en zone tempérée, la taille de l’habitat importe plus que son hétérogénéité, son degré d'insularisation et sa structure végétale. Pour conserver durablement son potentiel faunistique et constituer un Paysage forestier intact, la superficie boisée devrait atteindre au moins (500 km2 (soit 50 000 hectares), d'un seul tenant[23].
Deux processus majeurs caractérisent la fragmentation forestière : la perte d’habitats viables pour les espèces typiquement forestières, et l'isolement des habitats restants. Les conséquences pour les espèces peuvent être :
- positives pour les espèces de lisières et de clairières. La création de clairières maintient l'ouverture du paysage propice à ces espèces là où les grands herbivores ont disparu ;
- négatives pour la biodiversité forestière à la suite de la dégradation de leurs conditions environnementales (invasions d’espèces exotiques, perte et dégradation d'habitats et de microclimats forestiers, perte de ressource alimentaire ; disparition des espèces les plus vulnérables, déstructuration des réseaux trophiques).
La réponse des populations des sites fragmentés dépend de l’organisation écopaysagère, de la fréquence et de l’intensité des perturbations et de la durée de l’isolement.
Les Impacts correspondent à la fragmentation et à la destruction d'une partie des habitats naturels forestiers, et sont aggravés par[24] :
- l'incapacité des petits fragments forestiers à abriter durablement des populations viables d'espèces forestières (de grands vertébrés notamment), ce qui induit un appauvrissement génétique et au-delà d'un certain seuil un risque de goulot d'étranglement génétique, qui peut être un prélude à la disparition d'espèces. La dégradation de l'intégrité écologique d'un massif boisé peut y perturber les flux de gènes intra-population et/ou au sein des métapopulations, avec des effets secondaires sur la diversité génétique, voire un effet de « piège écologique » ;
- l'extinction locale d'espèces ne disposant plus d'au moins un fragment capable de supporter une population viable ;
- un « effet de bord » (ou effet-lisière), correspondant à une dégradation des conditions de vie des espèces de cœur de forêt près des lisières (Une base de données spéciale a été créée pour mieux suivre l'évolution des lisières dans le monde ; elle a été dénommée BIOFRAG[25]).
La flore, la faune et la fonge typiquement forestières, c'est-à-dire inféodées aux cœurs de forêts sont les plus touchées. Elles le sont d'autant plus que le patch est petit et isolé (l'isolement dépend de l'accessibilité des patchs similaires proches, ainsi que du contraste « écopaysager » avec les zones environnantes[3] (Les écologues parlent à ce sujet de « perméabilité de la matrice ») ; ainsi, si l'environnement du fragment est un chablis en cours de régénération naturelle ou un milieu bocager fortement maillé, ce fragment sera moins isolé que dans un contexte d'openfield intensivement cultivé. La diversité structurelle de la végétation du contexte paysager (matrice) permet de réduire l'isolement des fragments de forêt. Lorsqu'une grande partie des sols boisés est convertie de façon permanente en pâturages, champs ou par l'urbanisation, les fragments de forêt relictuelle et le biote[26] qu'ils abritent seront souvent très isolés et moins résilients[3]. Les îlots boisés les plus petits et/ou les plus isolés perdent leurs espèces plus rapidement que ceux qui sont plus grands et/ou moins isolés[3]. Un grand nombre de petits "îlots boisés" ne peuvent abriter la même biodiversité que l'équivalent en surface non fragmentée, même sur une surface totale cumulée plus grande.
Biogéographie insulaire
Selon la biogéographie insulaire (écologie du paysage), pour les espèces typiquement forestières, les îlots boisés relictuels fonctionnent comme des îles dans le paysage (où les pâturages, champs et, les zones imperméabilisées ou construites sont comme une mer qui isole les boisements et qui empêchent ou limitent les espèces forestières de circuler d'une tache boisée à l'autre[27]). Les espèces, guildes et communautés inféodées à la forêt profonde (ou aux cœurs de fragments de massifs forestiers) sont les plus concernées. Strictement forestières, elles vivent exclusivement en milieux forestiers et par la forêt. Elles y exploitent souvent préférentiellement une strate ou un habitat particulier. Ces espèces peuvent être dépendantes de massifs riches en arbres anciens ou d'habitats typiquement forestiers ; elles peuvent dépendre d'une essence particulière, d'une hauteur d'arbre ou de l'âge de l'arbre (Cf. bois sénescents, bois sur-âgé, bois-mort). D'autres espèces sont inféodées à l'humus forestier qu'elles contribuent à créer comme les champignons ou bactéries et peuvent être également dépendantes d'une ou plusieurs espèces symbiotes (ex : symbiose arbre-champignon, insecte saproxylophage[28]-champignon-bactérie, etc.) ou hôte. Ces espèces semblent dépendre du microclimat forestier (plus tempéré en forêt que sur les lisières ou en milieu déboisé à latitude et altitude comparables). Elles sont aussi sensibles à l'ambiance forestière : en forêt, le rayonnement solaire, UV notamment, est filtré par la canopée et la lumière est tamisée avant d'arriver au sol, toute l'année en zone tropicale humide, et environ sept mois par an en régions tempérées. Toute rupture linéaire ou de grande taille de la continuité du couvert forestier est source de morcellement écologique ; plus ou moins grave ou gênant, selon son ampleur ou selon le degré de dépendance des espèces aux conditions microclimatiques forestières.
La fragmentation forestière englobe la perte nette d'habitats, le morcellement physique et le morcellement fonctionnel (fragmentation écologique) des forêts[29]. En biologie de la conservation, la notion de fragmentation de l'habitat inclut la destruction de l'habitat, sa réduction de superficie et pas seulement la partition induite par les infrastructures[30]. Différentier les effets de ces trois facteurs reste difficile car ils interagissent entre eux et avec d'autres[31].
Évaluation de la fragmentation

L'évaluation de la fragmentation réelle d'un habitat est rendue difficile par le fait que les effets (sanitaires, génétiques et de dynamique des populations) d'une insularisation ne touchent pas toutes les espèces au même rythme ni avec les mêmes conséquences[32]. L'importance et la brutalité ou lenteur des effets varient beaucoup, notamment selon la durée de vie de ces espèces, leur diversité génétique initiale, leur cycle de reproduction et leurs capacités à se déplacer et à efficacement échanger leurs gènes[32].
Évaluation « spatiale » de la fragmentation physique des forêts
Il est utile voire nécessaire (pour l'écocertification, ou dans le cadre du réchauffement climatique et de la lutte contre le développement des gaz à effet de serre, cf. REDD+ par exemple) de pouvoir quantifier la fragmentation spatiale des forêts[33], notamment en régions tropicales où la fragmentation est rapide et intense et où, routes et pistes ne figurent pas toujours sur les cartes et dépendent de la qualité des images satellitales.
Le résultat d'une évaluation de la fragmentation forestière dépend bien évidemment de l'échelle d'analyse[34], et également de la définition retenue d'une forêt. Plus cette définition retient un seuil important de densité d'arbres, plus les facteurs de fragmentation retenus sont petits et, plus la forêt apparait fragmentée et moins, les catégories de bordure et de transition ou de patch ont de poids.
La mesure faite à partir d'images aériennes ou satellitales est également sensible au degré de précision de l'image (nombre de pixels rapportés à la surface analysée) et aux facteurs climatiques. L'amélioration de l'imagerie satellitale devrait permettre celle de la mesure de la fragmentation[35],[36],[37]: la fragmentation apparente augmente fortement en se fondant sur des cartes à "grain plus fin" en forêt tropicale humide, là où les images satellitales donnent l'impression d'une forêt pas ou peu fragmentée.
Il est important de distinguer entre la fragmentation naturelle et celle d'origine anthropique, la seconde a souvent beaucoup d'importance en termes d'impacts[38] et donc pour la gestion restauratoire. Ceci implique de connaitre l'écopotentialité du territoire (les cartes de la végétation potentielle sont qualitativement intéressantes mais plus grossières, de deux ordres de grandeur que les cartes de couverture terrestre[39]. Cela demande des cartes assez précises pour ne pas avoir trop d'incertitudes sur le statut forestier et naturel, ce qui n'est pas toujours possible.
Évaluation des effets génétiques
Une enquête génétique devrait théoriquement permettre d'évaluer le degré réel de « fragmentation démographique », mais dans un milieu très anthropisé, les introductions artificielles d'espèces ou de gènes (éventuellement non appropriées sur le plan de l'adaptation) par l'homme ou ses voies de transport perturbent ce type d'analyse, de même que les analyses de dépression de consanguinité et/ou de perte de variabilité génétique au sein d'une population ou d'un habitat ou îlots d'habitat[32].
De plus, jusqu'à un certain seuil, même si le nombre d'individu diminue, la variation génétique d'une espèce dont la population est fragmentée ne disparait pas entièrement[32]. Et dans quelques cas, cette population fragmentée peut perdre moins de diversité génétique qu'une population panmictique de taille égale soumise à une forte et nouvelle pression[32]. Par conséquent, tant que le « taux d'extinction locale » reste faible, ou tant qu'il est contrecarré par un programme de réintroduction approprié (gènes diversifiés et adaptés au contexte), une population fragmentée peut conserver la quasi-totalité de sa richesse génétique au niveau mondial durant une longue période[32].
Les types de fragmentation
Deux grands types de fragmentation, naturelle et artificielle peuvent être distinguées, dont les enjeux et conséquences diffèrent beaucoup.
 Les grands cours d'eau (Blue Earth river, Minnesota) : compartimentage naturel des forêts mais également corridors (vecteurs d'arbres et bois flottants comme de propagules). Les gués permettent la traversée des espèces terrestres en période de basses eaux
Les grands cours d'eau (Blue Earth river, Minnesota) : compartimentage naturel des forêts mais également corridors (vecteurs d'arbres et bois flottants comme de propagules). Les gués permettent la traversée des espèces terrestres en période de basses eaux Forêt nationale du Mont Baker-Snoqualmie (États-Unis) : la topographie montagnarde (couloirs d'avalanches, pentes fortes, orientations) et le climat d'altitude constituent les facteurs naturels de distribution de la forêt d'altitude. La limite supra-forestière se présente naturellement sous la forme de digitations plus que de morcellement (voir lisière)
Forêt nationale du Mont Baker-Snoqualmie (États-Unis) : la topographie montagnarde (couloirs d'avalanches, pentes fortes, orientations) et le climat d'altitude constituent les facteurs naturels de distribution de la forêt d'altitude. La limite supra-forestière se présente naturellement sous la forme de digitations plus que de morcellement (voir lisière) Il est parfois difficile de distinguer les facteurs humains et naturels de fragmentation forestière. La géomorphologie, les microclimats interfèrent ici avec les effets du pâturage et de l'exploitation des arbres (Vue depuis le sommet du Monte Grappa vers les Alpes et les Dolomites)
Il est parfois difficile de distinguer les facteurs humains et naturels de fragmentation forestière. La géomorphologie, les microclimats interfèrent ici avec les effets du pâturage et de l'exploitation des arbres (Vue depuis le sommet du Monte Grappa vers les Alpes et les Dolomites) Boisement insularisé en fond de vallon (Cwm yr Annell, Royaume-Uni) : l'hydromorphie, la raideur des pentes peut participer au maintien de parcelles forestières
Boisement insularisé en fond de vallon (Cwm yr Annell, Royaume-Uni) : l'hydromorphie, la raideur des pentes peut participer au maintien de parcelles forestières Trzy Korony vu d'Okruglica, 2007 : hors zones protégées, la forêt est moins fragmentée d'altitude mais avec des sols plus pauvres et avec moins d'essences d'arbres ; dans ce paysage semi-naturel, la fragmentation croît vers les fonds de vallée
Trzy Korony vu d'Okruglica, 2007 : hors zones protégées, la forêt est moins fragmentée d'altitude mais avec des sols plus pauvres et avec moins d'essences d'arbres ; dans ce paysage semi-naturel, la fragmentation croît vers les fonds de vallée
Aux rythmes des temps géologiques
- La lente formation tectonique des chaines de montagnes (altitude), de détroits ou de bras de mer, de grands fleuves, de variations du niveau marin, de configurations océans-continents a fragmenté de vastes régions forestières. C'est le cas de biomes séparés de milliers de kilomètres par la dérive des continents comme les forêts tropicales d'Afrique de l'Ouest et de l'Amazonie réunies sur le supercontinent de la Pangée qui s'est ensuite morcelé.
- À échelle continentale et à celle des modifications paléoclimatiques, on peut évoquer l'exemple des forêts tropicales humides du Carbonifère (-300 millions d'années environ) de l'Euramérique se sont scindées en sous-ensembles lors d'une réchauffement et un assèchement du climat qui ont entraîné la diminution de la diversité des amphibiens mais ont stimulé la diversification des reptiles [27]. Ces changements lents se sont déroulés sur des dizaines voire des centaines de millions d'années, ils sont source de l'évolution et de la diversification du Vivant. Ces échelles de temps ne sont absolument pas comparables à la vitesse, rythmes de la fragmentation ou de la destruction des forêts par l'Homme, rapportés à échelle géologique, ces bouleversements sont brutaux (Cf. la Sixième extinction)[27].
- En régions circumpolaires, à un rythme moins lent, les cycles glaciaires ont plusieurs fois détruit ou fragmenté de vastes paysages forestiers, avec constitution de nouvelles répartitions des biomes ; de même que les montées et baisses du niveau marin qui ont accompagné ces cycles glaciaires
Lors des dernières périodes glaciaires, des refuges ont permis aux espèces de la forêt une migration vers les néosols qui se formaient là où l'océan reculait, sur les actuels plateaux continentaux.
Ces variations biogéographiques ont été d'autant plus marquées que proches des pôles, en altitude ou en régions péri-désertiques. Elles semblent avoir été lentes et plutôt propices à la diversification génétique des espèces forestières et semi-forestières. En zone circumpolaire, la taïga est plus riche en grandes clairières tourbeuses ou entretenues par les herbivores se transformant au Nord en taches boisées éparpillées sur de vastes étendues couvertes de lichens (écotone).
Les dynamiques naturelles de fragmentation
- Une fragmentation généralement plus momentanée et locale peut être induite par des couloirs d'avalanche, des glissements de terrains, des incendies naturels de forêts[27]. Lorsque ces incendies sont causés par l'homme et répétés (comme en Australie), ils peuvent être cause de désertification et de disparition de nombreuses espèces.
- en zone tempérée ; les oiseaux sont le plus souvent migrateurs ; certains sont aussi plus ou moins inféodés au cœur de forêt le temps de leur reproduction. Les autres espèces forestières sont plus strictement liées à la forêt, voire à son seul « cœur ».
La forêt tempérée était autrefois habitée par de nombreux grands herbivores (bisons, élans, aurochs, etc.) qui y entretenaient de vastes clairières et probablement des corridors plus ouverts entre ces clairières. Les grands mammifères jouent encore ce rôle en Afrique. Il n’existe pas d’indices laissant penser que ces corridors aient pu avoir un réel rôle de fragmentation ; - en zone aride, la forêt est remplacée par la savane, ou la toundra où le feu, les grands herbivores entretiennent naturellement des ilots boisés et des arbres épars, là où l'eau est suffisamment présente ;
- dans les plaines et petites montagnes des zones tropicales humides, la forêt forme généralement un continuum physiquement continu et apparemment homogène. En zone tropicale humide, sous la canopée la température est fortement tamponnée et dans les terriers elle est quasiment stable. Elle intègre pourtant des sous-ensemble écologiquement différentiés, avec en particulier la mangrove près de la mer, la forêt galerie au-dessus des cours d'eau, des ripysilves très denses aux abords des grands fleuves, des végétations associés au relief ou contextes édaphiques particuliers (ex : inselbergs guyanais) ;
- dans tous les cas, la forêt se raréfie naturellement en haute altitude (écotone), en présentant des phénomènes naturels d'insularisation écologique de sous-populations d'arbres (sur des parois mieux exposées par exemple).
- aux échelles locales et paysagères, la fragmentation forestière la plus visible est celle produite par les grands cours d'eau (quand la canopée est disjointe et les berges assez éloignées pour que les troncs tombés ne puissent plus faire « passerelle » entre les deux rives).
Néanmoins, une connectivité discrète existe parfois : Ainsi, lors de fortes et parfois fréquentes inondations, certains animaux sont obligés de quitter leurs îles à la nage ou sur des radeaux de végétation[40] par exemple constitués d'hélophytes (Ptéridophytes, Poacées et Cypéracées en Guyane à la suite de la création du barrage EDF de Petit-Saut[40]) et d'hydrophytes. De même, des branches et troncs d'arbres couverts d'épiphytes ou contenant du bois mort partent à la dérive peuvent jouer le rôle de « radeaux » transportant diverses espèces sur de grandes distances et jusqu'en mer parfois. De plus, certains animaux comme le renne, le cerf, le chevreuil ou le caribou des forêts (R. tarandus caribou ou des bois, sont d'excellents nageurs ; ils peuvent traverser des fleuves ou lacs de grande taille, à condition que les berges ne soient pas artificielles et verticales. Enfin, en climat froid, certains grands mammifères carnivores ou herbivores peuvent traverser les fleuves gelés (comme à l'hiver 1956 pour certaines rivières et fleuves européens) ; et les cours d'eau non rectifiés et non aménagés possédaient souvent des gués naturels (estivaux ou de saison sèche) permettant une traversée plus facile par certaines espèces. Enfin, dans un bassin versant les cours d'eau ont une forme globalement arborescente, qui permet à certaines des espèces qui fuient le milieu liquide (ex : fourmis) de peu à peu contourner le cours d'eau.
Les fragmentations anthropiques
Ce sont des déterminants à intégrer dans la réflexion sur les mesures de gestion restauratoire et de protection des habitats et de la biodiversité.
 Goleniów, (Pologne) : fragmentation majeure d'un massif forestier par une autoroute
Goleniów, (Pologne) : fragmentation majeure d'un massif forestier par une autoroute L'agriculture moderne contribue au mitage de la forêt mais certaines pratiques agro-sylvo-pastorales, selon les traditions et le mode de propriété foncière, maintiennent un bon niveau de biodiversité. De l'Antiquité à nos jours, des formes d'agrosylviculture et de forêts paysannes ont intimement mêlé arbre et culture.
L'agriculture moderne contribue au mitage de la forêt mais certaines pratiques agro-sylvo-pastorales, selon les traditions et le mode de propriété foncière, maintiennent un bon niveau de biodiversité. De l'Antiquité à nos jours, des formes d'agrosylviculture et de forêts paysannes ont intimement mêlé arbre et culture. Fragmentation par une remontée mécanique : impact sous évalué et a priori moindre que celui d'une route. Ce remonte-pente touristique du château du Rietburg (de), à Rohdt (Allemagne) ne fonctionne que de jour. Si la tranchée est assez large et bien exposée au vent ou au soleil, elle peut tout ou partie de l'année entretenir un microclimat non forestier sur toute sa longueur
Fragmentation par une remontée mécanique : impact sous évalué et a priori moindre que celui d'une route. Ce remonte-pente touristique du château du Rietburg (de), à Rohdt (Allemagne) ne fonctionne que de jour. Si la tranchée est assez large et bien exposée au vent ou au soleil, elle peut tout ou partie de l'année entretenir un microclimat non forestier sur toute sa longueur Coupes de bois, Central highlands, Victoria : à 50 km au nord-est de Melbourne, où quelques spécimens de Gymnobelideus leadbeateri, espèce d'opossum supposée disparue depuis 1900, redécouverte en 1960[41]. La surexploitation permise par les routes, peut aussi dégrader les capacités de puits de carbone et stock de carbone du sol forestier.
Coupes de bois, Central highlands, Victoria : à 50 km au nord-est de Melbourne, où quelques spécimens de Gymnobelideus leadbeateri, espèce d'opossum supposée disparue depuis 1900, redécouverte en 1960[41]. La surexploitation permise par les routes, peut aussi dégrader les capacités de puits de carbone et stock de carbone du sol forestier.

(carte de Cassini)
Les facteurs de cette fragmentation artificielle (ou anthropique) sont par exemple :
- les infrastructures de transport (routes, voies rapides, autoroutes, canaux, voies ferrées…). On a expérimentalement démontré (in situ et en laboratoire) que le bruit routier est à lui seul une source de fragmentation, notamment pour les oiseaux chanteurs migrateurs en halte migratoire.xxxxxxxxx La fragmentation est aggravée lorsqu'ils sont clôturés ou dans les cas des canaux lorsque les berges verticales (souvent des palplanches) et ne permettent pas aux animaux de descendre et sortir de l'eau, et quand l'infrastructure n'est pas compensée par un nombre suffisant d’écoducs.
La pollution lumineuse est rare en forêt mais elle peut toucher ses lisières ; elle est souvent atténuée par l'épaisseur du feuillage mais peut être localement importante au bord de l'eau, en bordure de clairière ou de zones humides, sur les îles tropicales… dont en raison de l'attirance parfois fatale de certaines espèces forestières pour la lumière (à ce sujet voir les articles Pollution lumineuse et environnement nocturne, piège écologique) ;
Quelques espèces (souvent de grande taille, ou protégées par un venin (serpents, crapauds), une carapace (tortues), des piquants (hérissons, porcs-épics…) traversent plus facilement les routes mais s'y font facilement écraser (voir article Roadkill à ce sujet). Les routes en forêt affectent négativement (via le roadkill mais aussi par le salage) de nombreuses populations d'amphibiens, ou limitent leurs déplacements[42]. - les voies de circulation internes de la forêt (routes forestières) : Ce sont des pistes damées et parfois désherbées ou empierrées voire bitumées dont la répartition est déterminée par le foncier et les objectifs d'exploitation[43]. Ces voies sont des facteurs de fragmentation pour de nombreuses espèces[44],[45], plus encore que les coupes à blanc dans les Rocky mountains[45] et de manière aggravée sur les zones de pente[46].
Elles contribuent souvent au drainage et donc aux crues[47] et localement à l'érosion du sol forestier[48] et à la dégradation des cours d'eau[49],[50].En zone tempérée, dans la chaîne des Cascades (Ouest de l'Oregon) il a été montré que l'association de coupes rases et de routes double presque le débit maximum de crue des cours d'eau forestiers[49], privant la forêt d'une partie de son eau).
par radio-tracking[51] et avant cela de « Capture-marquage-recapture », une thèse[52] en Écologie du paysage a montré en 1991 que même des routes forestières non fréquentées constituaient un obstacle majeur aux déplacements de carabidés forestiers comme Abax ater (coléoptère fréquent des lisières forestières et du bocage), qui bien que sachant parfaitement voler, «refuse» de traverser une route coupant son habitat.
Le linéaire des voies intraforestières a beaucoup augmenté depuis quelques décennies[53].
Leur impact dépend aussi de la densité et des horaires (diurnes/nocturnes) ou de la saisonnalité des flux de véhicules, mais il existe même avec peu de véhicules. Ainsi en Amazonie même d'étroits chemins de terre étroits accueillant moins de circulation cinq véhicules/jour peuvent significativement réduire l'abondance locale (17) et les passages[54] de certaines espèces d'oiseaux de sous-bois ; et de manière plus spectaculaire, des coupes linéaires de moins de 200 m de large (typiques des grands axes routiers) suffisent à complètement bloquer les mouvements d'espèces inféodées à la forêt, empêchant par exemple des oiseaux ayant été expérimentalement translocalisés de l'autre côté de la route de retourner dans leurs territoires vers leurs compagnons à vie[55]. - L'entretien routier aggrave parfois l'effet fragmentant, par exemple avec des fauchages annuels, avec le salage, par l'usage de pesticides sur les accotement, via l'entretien d'un drainage par un réseau de fossés, à cause d'un éclairage nocturne ou simplement du bruit et de diverses formes de dérangement. Quand ces voies sont des pénétrantes de "cœurs d'habitats", les impacts peuvent être exacerbés en touchant des espèces plus rares et vulnérables :
certains éléments artificialisants de l'exploitation forestière (cloisonnements, aménagements sylvicoles, coupes rases), l'usage répété du feu et/ou de pesticides peuvent fragmenter l'espace au détriment des taxons forestiers et au profit d'espèces pionnières de milieux ouverts. Le tassement de sols limoneux et tourbeux vulnérables au poids des engins peut aussi être en cause. L'importance de ces impacts est encore mal mesurée et parfois discutée ; - le labour de parcelles intraforestières ou traversant un massif de part en part peut aussi être fragmentant pour les espèces strictement dépendantes du milieu forestier ;
- de nombreuses espèces forestières pour des raisons biologiques fuient les espaces artificialisés (ou simplement ouverts et minéralisés). Elles peuvent se retrouver « enfermées » en petites populations génétiquement isolées entre trois ou quatre infrastructures (voies ferrées, canaux, etc., (Cf. phénomène de fragmentation écopaysagère).
Ces causes et effets se cumulent au détriment de la biodiversité.

Évolution et géographie de la fragmentation


La fragmentation, l'artificialisation et la conversion des sols ont leur propre géographie, qui répond à des facteurs humains (sociaux, industriels, économiques…), d'accessibilité et de biogéographie ; avec des impacts sur la génétique du paysage.
- De long temps, l'anthropisation des milieux se concentre autour des villes et villages, principalement dans les vallées (proximité de l'eau) et dans les régions riches en ressources faciles à exploiter ou à parcourir (parcours d'altitude). Peu à peu les hommes exploitent ou réduisent la forêt ancienne et les lambeaux de forêts anciennes (pas ou peu modifiées par l'homme depuis quelques siècles) sont souvent « mités » et fragmentées, dont par les routes forestières ;
- La révolution industrielle a entraîné l'augmentation des régions de grandes cultures et d'exploitation du bois. Au XXe siècle, l'augmentation exponentielle de la démographie mondiale et le modèle consumériste ont permis l'accroissement des villes et des industries. Les besoins toujours croissants et de plus en plus délocalisés en ressources (minérales, organiques, alimentaires, hydriques, énergétiques, bois et fibre…) ont eu des impacts forts sur les milieux forestiers. Les ressources sont en partie forestières (gibier, bois, ressources minières). Elles sont transportées via des réseaux de plus en plus denses et interconnectés, passant d'une structure en arborescence (moins morcelante) à de véritables quadrillages du territoire.
- Les forêts de plaines (plus accessibles à l'ancien maillage routier), ont été les premières touchées ; le développement des transports pour les traverser (canaux, autoroutes, voies ferrées) ou les exploiter (voire surexploiter comme en région tropicale où le mitage puis la déforestation commencent presque toujours le long des routes).
- Là où la démographie et la densité de population sont élevées, et là où la route a facilité l'accès au cœur de la forêt, la pression sur les cœurs d'habitats forestiers augmente. Inversement les forêts boréales ou tropicales pauvres et d'accès difficile contiennent un pourcentage plus élevé de forêt intérieure[56].
Les forêts de la zone intertropicale
La déforestation des régions intertropicales s’est considérablement accrue dans le courant du XXe siècle, à la faveur de changements politiques et économiques. Elle est devenue une des préoccupations majeures de la communauté scientifique et du grand public.
Les forêts tropicales ont longtemps conservé leur intégrité bien qu'habitées par des millions de personnes mais l'arrivée des routes, associée à la double pression démographique et de la pauvreté ont au XXe siècle amorcé une fragmentation accélérée de ces forêts. Les forêts tropicales (moins de 7 % des terres émergées) hébergent plus de 50 % de la biodiversité. Outre le rythme accéléré de la perte de diversité biologique, s'ajoute le rôle dans les changements climatiques globaux (voir cycle du carbone).
Les causes de la déforestation sont :
- l'exploitation des ressources forestières :
- la forte croissance démographique entraînant un besoin accru en surfaces de cultures vivrières ;
- le développement des cultures de rente (industrielles) comme le café, le cacao, l'hévéa, les bananes, l'huile de palme, etc.
Par exemple en Côte d'Ivoire, plus de 80 % des surfaces forestières ont été converties en cultures entre 1969 et 1993 ; - le front de déforestation en Afrique tropicale se présente sous la forme d'un morcellement paysager de type mosaïque de forêts-cultures-jachères : une forêt continue se transforme en une multitude d'îlots;
- le front de déforestation amazonien est plus continu et de plus en plus fragmentant[57]. Les lignes à haute tension, les barrages, les pipe-lines s'étendent jusque dans des secteurs très reculés de l'Amazonie notamment[56] (dont au coeur de la Guyane avec le barrage de Petit-Saut comme le CNRS et d'autres l'ont montré à partir de la station scientifique de Saint-Eugène[58],[59], à partir de plus de 10 ans d'études[60],[61],[62],[63],[64],[65],[66],[67],[68].
Au Brésil voisin, dans la région de Rondonia, l'imagerie satellitaires montre clairement que les routes et pistes forestières expliquent le caractère maintenant très fragmenté de la forêt résiduelle[69], mais ailleurs en Amazonie, les grands fleuves et des accès par le littoral ou certains plateaux ont aussi permis le recul et la fragmentation des forêts.
Les forêts d'Amérique du Nord
Aux États-Unis, l'intégrité forestière n'a été conservée que dans quelques parcs nationaux et sur les reliefs marqués, certains bords de cours d'eau torrentiels[56]. La forêt publique exploitée n'a pas échappé à la fragmentation routière.
- Les forêts bordées par des usages anthropiques risquent-elles plus la dégradation que celles qui partagent une forte proportion de leurs écotones avec un milieu non forestier, mais naturel comme les zones humides ?
En 2003, une analyse satellitale (via radiomètre Avancé à Très Haute Résolution (AVHRR) a cherché à différencier les causes naturelles et anthropiques du morcellement : à échelle satellitale, plus de 50 % de la forêt tempérée feuillue et mixte et, près de 25 % de la forêt tropicale sont au début des années 2000 effectivement morcelés ou détruits par l'Homme, contre seulement 4 % de la forêt boréale. L'Europe est la plus touchée par la fragmentation anthropique et l'Amérique du Sud, la plus épargnée [70]. Les hot spots de forêt perforées ont été cartographiées, par exemple pour l'Est des États-Unis[71].
En Europe

1)un incendie de forêt en Espagne (effet provisoire sauf s'il y a répétition des incendies dans une même zone),
2) la construction d'une autoroute trans-européenne
3) d'un défrichement de ripisylve et des restes d'une forêt alluviale (d'après la cartographie des forêts du CCR 2000 et 2006)
- En 2013, selon les données réunies par l'« European Forest Data Centre » (EFDAC) [73] mis en place par la commission européenne, et un récent rapport sur le paysage forestier européen (« Forest landscape in Europe:Pattern, fragmentation and connectivity »[72]), 70 % du territoire de l'UE a des forêts trop fragmentées pour permettre la survie de la biodiversité, à cause des routes, de l'agriculture, des établissements urbains et des feux de forêt. Une légère et régulière augmentation surfacique de la forêt européenne (+0,4 %/an) n'a pas servi à améliorer la connectivité entre les fragments forestiers ; au contraire, le morcellement forestier progresse encore[72], en contradiction avec les objectifs de la conservation de la nature[74], et avec ceux des Directives Oiseaux (79/409/EEC)[75] et Natura 2000 (92/43/EEC)[75] ;
- L'Europe du Nord a longtemps été épargnée mais le réseau de routes forestières s'y est fortement développé à la fin du XXe siècle, en particulier en Finlande où les routes et pistes forestières occupaient en 1998 environ 1 300 km2 (surface en croissance). En Finlande, le secteur forestier consommait en 1998 environ 25 % des carburants fossiles utilisés dans le pays[76], avec des changements importants dans la structure du paysage (tendance à la fragmentation des années 1940 aux années 1970 et à une certaine fermeture des paysages à partir des années 1970 aux années 1990[77] (avec néanmoins une perte de forêts anciennes et naturelles vitales pour beaucoup d'espèces menacées[77]). L'évolution a été comparable en forêts publique et privée[77].
- L'Europe de l'Ouest a connu une longue période de fragmentation par les routes et autres infrastructures linéaires. Depuis quelques décennies, une tendance à la reconnexion physique par reboisements spontanés ou artificiels est constatée en montagne et en secteurs de déprise agricole. Selon les données de Corine Land Cover, en considérant à la fois les changements de structure spatiale de la forêt et de perméabilité moyenne de la matrice paysagère, la connectivité inter-fragments semble avoir peu évolué de 1990 à 2000. Une légère augmentation de la connectivité moyenne est constatée essentiellement due à quelques pays à faible couvert forestier où les changements ont été spectaculaires ;
- Les effets de la fragmentation sont aggravés par le manque de zones-tampon : en 2015, environ 40 % des lisières des forêts européennes jouxtent ou sont à moins de 100 m de terres non forestières principalement utilisés pour l'agriculture intensive. Et près de 15 % des forêts en Europe sont fortement fragmentées par des utilisations intensives des terres, ce qui diminue leur capacité d'accueil et de circulation des espèces forestières et de lisières entre éléments boisés du paysage. Ceci renforce aussi la probabilité d'exposer les lisières et bordures intérieures des forêts à des espèces envahissantes, des ravageurs et des maladies.
- La commission européenne a démontré en 2013[72] la faisabilité d'une évaluation harmonisée de la fragmentation paysages forestiers d'Europe via un ensemble reproductible d'indicateurs de fragmentation[78] et d'indices établis par le CCR. Ces indices permettent de mesurer les progrès faits ou à faire dans l'atténuation de la fragmentation et pourraient être appliqués à d'autres écosystèmes[72]. Les résultats du rapport de 2013 ont été utilisés par l'Europe dans ses rapports déclaratifs statistiques et qualitatifs sur la Forêt et la protection des forêts en Europe à la CEE-ONU et à la FAO, alors que note la Commission, les données structurées, comparables et officielles sur les tendances des forêts n'existent pas encore aux niveaux nationaux[72].

B. profil national de structure du paysage forestier (données 2006). 1) éléments boisés parfois naturels, 2) mixte naturel-artificiel , 3° forêt à dominante naturelle, 4) Cœur de forêt naturel,
C. Profil national des écotones de lisières de forêt(données 2006). 1) lisières artificielles, 2) lisières naturelles
En France

« En Île-de-France, 7 200 kilomètres d’infrastructures de transport (routes et toutes voies carrossables forestières) sillonnent les 269 000 hectares (selon l'IFN) de forêt (22 % du territoire francilien) »[79]

La France est concernée par la fragmentation forestière : l'IRSTEA considérait (en 2012) que « si la forêt progresse en France, elle est de plus en plus morcelée par toutes sortes d'infrastructures : cette fragmentation est de nature à affecter la dynamique de la biodiversité dans le temps et l'espace »[80].
- Les données archéologiques et paléobotaniques montrent que les peuples celtiques ont déforesté et fragmenté une partie des forêts dès les âges du fer. Le phénomène s'est accentué avec la colonisation romaine (voies romaines, installation de grandes villae) et s'est poursuivi de l'époque carolingienne à la fin du Moyen Âge par les grands défrichements médiévaux et la construction de nouvelles routes. L'impact du réseau de routes et de chemins antique et moderne n'est évidemment pas comparable concernant la limitation à la circulation des espèces que ne l'est l'actuelle densité et imperméabilité (clôtures) des structures et de fréquentation.
- Pour sauvegarder pour le Royaume de France des réserves vitales de bois d'œuvre, J.-B. Colbert met un frein à la déforestation mais le morcellement des forêts par les routes s'est poursuivi (de la Renaissance à la Révolution française) ; les cartes de Cassini montrent des forêts royales à la fin du XVIIIe siècle entièrement quadrillées de voies forestières. La grande forêt royale de Compiègne conquise au Moyen Âge sur les marais et cultures de la période gallo-romaine apparaît sur les cartographies de l'Ancien régime découpée par le réseau de routes et chemins.
Cependant, en forêt, ces axes (de terre battue et/ou empierrés, parfois sols végétalisés) avaient un rôle de barrière plus faible que celui des voies fortement damées, drainées et/ou goudronnées construites XXe siècle. - La densité routière : le réseau français des routes secondaires (traversant des forêts ou non) a peu évolué depuis les années 1980 mais était déjà l'un des plus denses du monde, constituant une grande partie du réseau français total (28 515 km en 1980).
Au début des années 2000, en forêt publique métropolitaine, avec 30 160 km de routes forestières au total, on comptait de 1,8 km de routes pour 100 ha (goudronnées pour 16 %, empierrées pour 43 % et sur sol plus ou moins naturel pour les 41 % restants[81].
La forêt de Fontainebleau à la fin des années 1990[82] était traversée par deux voies ferrées, l’A6, plusieurs nationales à grand gabarit (N6, N7, N37, N152), des départementales, des routes forestières parfois goudronnées, soit : 144 km de routes, favorisant une pression touristique forte et diffuse (13 millions de visiteurs par an) « qui provoquent des coupures physiques, écologiques et sonores qui rompent l’unité du massif et perturbent la faune comme la quiétude des promeneurs »[82]. - La densité en TGV et autoroutes: les axes TGV se sont multipliés comme les autoroutes (passant - de 1960 à 1997 - de 170 km d'autoroute à 8 940, dont près de 6 500 km concédés). Ces axes ont aggravé les discontinuités spatiales forestières et les difficultés de restauration d'une trame verte et bleue y compris dans des régions riches en forêts comme en Lorraine où - selon les données disponibles - « malgré l’augmentation des surfaces forestières, la connectivité fonctionnelle entre les fragments forestiers n’est pas effective sur le plan de la diversité, démontrant ainsi l’importance de la présence de corridors forestiers pour faciliter la migration des espèces végétales »[83].
À la fin des années 1990, on trouvait par km2 de forêt et en moyenne : 590 m de route ; 160 m de grandes infrastructures ; et un certain linéaire, mal connu, de routes forestières et chemins plus ou moins carrossables ou enherbés[84]. - Les indices de fragmentation forestière : avant 1994, les données IFN n'étaient disponibles que pour 59 départements sur 101 (soit 11,2 millions d'hectares boisés), en France métropolitaine. L'IFN sous-estimait fortement le phénomène en excluant les boisements de moins de 4 ha, ne tenant pas compte ni de l'âge, ni de la qualité ou de l'écopotentialité des boisements ; une coupure n'étant en outre considérée comme fragmentante d'un massif qu’au-delà de 200 m de largeur. Sur cette base, on dénombre 32 500 ensembles forestiers de moins de 100 ha (représentant un peu moins de 6 % des forêts et bois de plus de 4 ha).
Cette approche ne permet pas la juste mesure de l’intégrité écopaysagère des massifs forestiers (un massif non fragmenté doit offrir des zones importantes hors lisières et ruptures anthropiques, même autres que d’origine sylvicole).
Selon ces critères, en 1994, seize ensembles forestiers étaient jugés moins fragmentés et formant des ensembles de plus de 50 000 hectares ;- Massif central (2 812 000 ha), en huit sous-ensembles parfois reliés par des corridors ténus),
- Alpes du Sud et Préalpes de Provence (1 338 000 ha),
- Massif landais (1 070 000 ha),
- Pyrénées et les Corbières (573 000 ha),
- Jura (361 000 ha),
- Corse centrale (259 000 ha),
- Vosges du Nord (226 000 ha),
- Massifs picards (130 000 ha),
- Ceinture verte Sud de Paris (121 000 ha),
- Plateaux bourguignons (111 000 ha),
- Massif des Maures (86 000 ha),
- Forêt d'Orléans (73 000 ha),
- Vallées du Rouergue (72 000 ha),
- Double saintongeaise (60 000 ha),
- Vosges du Sud (56 000 ha),
- Sologne (53 000 ha).
- Selon l'IFEN sur la base de l'IGN/BD Route 500, France : 0,76 km de route/km2 de territoire enforesté, dont 0,16 km de grandes infrastructures.
Et 230 000 hectares de zones boisées comportent de grandes infrastructures avec une densité dépassant 1 km par km2.
- Une sous-estimation de la fragmentation : en tenant compte de l'indice de compacité[85] et de la fragmentation réelle par les routes, canaux, TGV et facteurs de ruptures physiques de moins de 200 m de large, la forêt française apparaît bien plus fragmentée. Les calculs de l'IFN considèrent qu'une rupture de 200 m n'interrompt ni l'intégrité (continuité) forestière, ni le déplacement de nombreuses espèces forestières.
Or, les espèces strictement forestières, de cœur de forêt comme le cerf, l'ours, la martre et de nombreux coléoptères saproxylophages ou des prédateurs comme Notiophilus rufipes, ont besoin de tranquillité et de vastes ensembles non fragmentés, contrairement au chevreuil ou à certains rapaces qui acceptent ou recherchent des territoires hétérogènes, en mosaïque, au moins à certaines périodes de leur cycle de vie, comme les lisières et les petits massifs forestiers.
D’autre part, les infrastructures de transport modifient le microclimat forestier, tuent de nombreuses espèces et/ou réduisent leurs possibilités de déplacements vitaux, menaçant certaines populations animales et peut-être végétales et fongiques (Cf. populations en danger ou vulnérables). Inversement, l’ouverture de clairières sur un modèle naturel (chablis) dans les forêts artificielles serait bénéfique à certains taxons.
Les enjeux de conservation
 En Asie du Sud-Est, la destruction et fragmentation des forêts est la première cause de disparition de l'orang-outan
En Asie du Sud-Est, la destruction et fragmentation des forêts est la première cause de disparition de l'orang-outan Pour certaines espèces comme Hélice des bois : Arianta arbustorum), une route peu fréquentée peut jouer un rôle de barrière
Pour certaines espèces comme Hélice des bois : Arianta arbustorum), une route peu fréquentée peut jouer un rôle de barrière Suivi par capture-marquage-recapture, l'Hélice des bois refuse de traverser un chemin non imperméabilisé de 3 m de large[86], un sentier de 30 cm envahi par la végétation n'affecte pas ses déplacements[86]
Suivi par capture-marquage-recapture, l'Hélice des bois refuse de traverser un chemin non imperméabilisé de 3 m de large[86], un sentier de 30 cm envahi par la végétation n'affecte pas ses déplacements[86] Les routes forestières peuvent affecter la répartition, les déplacements voire la survie de nombreux amphibiens[42], dont la rainette européenne Hyla arborea [87]
Les routes forestières peuvent affecter la répartition, les déplacements voire la survie de nombreux amphibiens[42], dont la rainette européenne Hyla arborea [87] En Amérique du Nord, l'Ours noir, une des nombreuses espèces vulnérables aux collisions routières
En Amérique du Nord, l'Ours noir, une des nombreuses espèces vulnérables aux collisions routières En Guyane : des espèces à carapace et à piquants s'aventurent volontiers en espaces ouverts avec le risque d'être tués par des véhicules
En Guyane : des espèces à carapace et à piquants s'aventurent volontiers en espaces ouverts avec le risque d'être tués par des véhicules

Des écoducs spécifiques aux écureuils (les écureuilloducs) leur permettent de traverser un axe de transport sans toucher le sol, sur des cordes tressés ou mini-ponts suspendus

Ce sont des enjeux de connaissance, de conservation et développement durable, mais aussi de coûts. Les forêts et leurs sols sont concernés par les deux grands enjeux du Sommet de Rio de 1992 (biodiversité et climat)[88], car la forêt est la fois le principal réservoir terrestre (émergé) de biodiversité et l'un des puits de carbone majeurs (avec les prairies et les tourbières). Or, depuis au moins 40 000 ans[89] ; dès le néolithique[90], via l'invention du feu[89] et l'intensification de l'élevage, l'humanité a déforesté une part significative de la planète. L'Homme a ainsi commencé à fragmenter les forêts ; avec localement des effets de désertification ou d'érosion, même par des populations relativement modestes en nombre, comme les Mayas. Ceux-ci, en convertissant des sols forestiers en champs on causé l'érosion de jusqu'à environ 1 000 tonnes de sol par km2 et par an (qu'on retrouve sédimentées dans les lacs[91]). La déforestation est massive et ancienne en Chine et en Australie[89] et pour partie en Europe[92]. Il y a eu en Europe, corrélation entre la densité de population et l'intensité de la déforestation[92]. La fragmentation des forêts tropicales est plus récente ; intense et rapide à la fin du XIXe siècle, et encore amplifiée dans la seconde moitié du XXe siècle et ces dernières décennies. Les boisements et forêt tempérés - depuis 200 ans environ - regagnent du terrain, mais - jusque dans les années 2000 - sous des formes souvent artificialisées, et alors avec une faible biodiversité (par rapport à l'écopotentialité des territoires qu'ils occupent) ; Il ne reste presque plus de forêt ancienne ni à forte naturalité en zones tempérées de l'hémisphère nord. Après s'être adapté à 3 glaciations et deux périodes interglaciaires, les grands mammifères herbivores européens et de l'hémisphère Nord (bison d'Europe, cerf mégacéros, antilope saïga, auroch, cerf, élan, mammouth, bouquetin, chevaux sauvages…) y ont en grande partie disparu ou ne survivent qu'en populations ténues. De même, pour les grands carnivores (Lion des cavernes, ours des Pyrénées ours des cavernes, Glouton, loup, lynx, etc.).
Il existe un consensus scientifique[31] (explicité dans cet article) sur le fait que la fragmentation des milieux par l'Homme (des forêts notamment) est devenue :
- une des causes majeures de perte de biodiversité (via la perte de richesse spécifique et de diversité génétique notamment)[31] ;
- un frein au potentiel de recolonisation et d'auto-restauration des milieux (ce qui correspond à une moindre résilience écologique), car la fragmentation diminue le pouvoir de dispersion de la plupart des espèces forestières, et affecte les relations entre animaux et végétaux (dont l'alimentation et la zoochorie[93]) ;.
- une source de déséquilibre des relations d'interactions durables (ex : relations prédateurs/proies et relations parasites/hôtes). Ces déséquilibres pourraient interagir négativement avec les équilibres sylvocynégétiques que recherchent les forestiers et les chasseurs ;
- un obstacle à la surveillance et aux mesures visant à restreindre l'accès motorisé du public aux cours de forêts (plus le réseau routier s'étend, plus il est difficile à surveiller). Des infractions aux interdictions de circuler, et des violations d'accès réservés sont fréquentes. Une étude de cas a porté en 1997 sur une unité de gestion forestière (UGF de Temagami) où un plan de gestion prévoit que certaines zones restent ouvertes à l'exploitation en restant peu accessibles au public et interdites aux véhicules[94]. Cette étude a mis en évidence « des niveaux élevés de violation des accès contrôlés »[94].
L'intégrité des forêts, le maintien de paysages forestiers intacts, une naturalité suffisante, et une répartition spatiale correspondant à l'écopotentialité des milieux sont donc peu à peu devenus des enjeux importants pour la restauration et conservation de la biodiversité forestière et globale et pour une gestion dite « durable » des forêts. Dans le même temps, l'enjeu « puits de carbone » a pris du poids. Cependant, les évaluations, processus et conventions internationaux se sont focalisés sur l'ampleur de la perte ou (localement du gain) de surfaces boisées, délaissant d'autres facteurs tels que l'évaluation des sols, la représentativité biogéographique ou le degré réel de fragmentation des forêts.
Ceci pourrait être dû au manque d'évaluation de la fragmentation des forêts, à la difficulté de prévoir ou interpréter ses effets (étudiés à l'échelle des patch, alors qu'il faudrait pouvoir le faire à échelle des métapopulations[31]), ou au manque de consensus sur un « état de référence » pour la naturalité, et sur des échelles et indicateurs de fragmentation, ou des seuils de gravité. Ainsi, les États-Unis n'ont évalué le degré de fragmentation de leur forêt que dans les années 2000[95],[96],[97], avec publication en 2007 de données montrant qu'une faible partie des forêts peut encore être classée en "cœur d'habitat tant les forêts sont fragmentées[98]. L'intégrité écologique (non-fragmentation) est devenue un des critères de qualité et de gestion durable de la forêt aux États-Unis[99].
La plupart des arbres vivent en symbiose avec d’autres organismes (bactéries, champignons mycorhizateurs et pollinisateurs notamment). Ils croissent lentement. Ils n'assurent leur plein succès reproductif qu’après plusieurs décennies (voire siècles pour certaines espèces). Hormis pour quelques essences dont les graines sont entrainées par l’eau, ou transportées par des oiseaux, leur capacité naturelle à migrer pour s'adapter aux changements climatiques semble plus réduite que celle de nombre de leurs déprédateurs. Les arbres (et les guildes et communautés d'organismes forestiers en dépendant) sont donc a priori plus vulnérables que d'autres aux changements globaux et rapides (et à leurs conséquences secondaires telles qu'incendies, phytopathologies, épidémies, attaques parasitaires, etc.). De plus dans l'écosystème forestiers lui-même, certaines espèces (oiseaux par exemple) ont des capacités d'adaptation et surtout de mobilité bien plus rapide que les arbres. Dans un contexte associant dérèglement climatique et effondrement de la biodiversité, la capacité de la forêt à se régénérer et s'adapter devient un enjeu majeur pour la fourniture durable de bois, produits dérivés du bois, pâte à papier, et d'autres services écosystémiques… Elle dépend notamment de la conservation de l'intégrité écologique de vastes massifs et des corridors forestiers (intra-forestiers, interforestiers). La partie boisée de la trame verte et bleue cherche à répondre à cet enjeu, avec aussi en France un programme « AM-Tools » sur les « outils écologiques et légaux » pour la « migration assistée des forêts »[100].
En effet, même pour des espèces sachant parfaitement voler comme certains coléoptères, diptères et papillons qui semblent très affectés par la fragmentation des forêts, avec probablement déjà des effets d'effondrement de leur diversité génétique :
- une analyse de la distribution spatiale et de la structure génétique de populations belges du papillon Operophtera brumata (Geometridae) a montré que sa densité de population diminue avec l’isolement et la petitesse de la parcelle forestière, l’espèce disparaissant même de certaines parcelles. Selon les données génétiques récoltées, environ 25 % de la variation génétique intra-subpopulationnelle (Hexp) semble être perdue du fait de l'isolement des fragments forestiers, l'hétérogénéité génétique parmi les subpopulations est intermédiaire (Gst = 0,02), et le dendrogramme REML suit le patron géographique des parcelles d'étude, prouvant qu'un effet de distance géographique entre les subpopulations s'ajoute à l'effet d'isolement[101]
- des coléoptères coprophages forestiers jouent un rôle important en enterrant des excréments qui enrichissent le sol et contenant parfois des graines qui peuvent ainsi mieux germer. Certains se montrent sensibles à la fragmentation de leur milieu[102]
Enfin, aux États-Unis et au Canada, après avoir longtemps construit de plus en plus de routes en forêt, l'intérêt de ces dernières est maintenant mis en doute au regard de leurs impacts écologiques négatifs[103], des coûts (de construction et d'entretien) et, des avantages relatifs qu'elles génèrent (évidents pour la sylviculture intensive mais non pour le tourisme, en particulier, l'écotourisme qui génère des bénéfices croissants[104] et que les gouvernements veulent aussi développer[105], recherche des paysages forestiers non dégradés par les routes et coupes rases[106],[107]. Une accessibilité accrue est-elle souhaitable ? se demandent Bourgeois & al (2006)[108] Daniel Kneesha & al. ; « Il est indéniable qu’un réseau routier développé comporte de nombreux avantages : des régions, qui autrement seraient isolées, sont ainsi relativement accessibles au grand public. Néanmoins, il semble que l’expérience dans son ensemble (nature, tranquillité, atmosphère paisible et calme, etc.) est ce qui détermine le succès d’une visite d’écotourisme »[107].

Enjeu carbone
- La fragmentation des forêts par les routes facilitent la coupes et l'exportation du bois voire la destruction et conversion des forêts (tropicales notamment), avec des impacts sur le cycle du carbone. Le mémento aux décideurs [110] du GIEC rappelle que « Les stocks de carbone sont bien plus grands dans le sol que dans la végétation ». Tout changement d’affectation d’un sol forestier ou sa dégradation peut fortement modifier ses capacités de puits ou stock de carbone et d’émissions de carbone[110] ; Selon le GIEC, « Des quantités considérables de carbone ont été libérées en raison du déboisement opéré depuis plusieurs siècles aux latitudes moyennes et élevées et, dans la dernière partie du XXe siècle, dans les régions tropicales »[110].
Le « C.A.S. » (changement d'affectation des sols) est un des éléments à prendre en compte dans la comptabilisation du carbone, dont dans le cadre du protocole de Kyoto et de ses suites qui prévoient que « les Parties pourront prendre en considération, pour remplir leurs engagements en vertu de l’article 3, les incidences de leurs activités de boisement, de reboisement et de déboisement (BRD) ainsi que de leurs autres activités reconnues dans le secteur de l’utilisation des terres, du changement d’affectation des terres et de la foresterie ». - les routes occupent aussi de vastes surfaces autrefois enforestées ; selon Foltz et al., le retour des routes forestières et pistes et chemins forestiers inutiles à un état naturel, améliorerait l'infiltration et diminuerait l'érosion[111], et serait équivalent s'agissant des puits de carbone (39,5 - 48,5 millions de tonnes métriques) à la revégétalisation d'une superficie supérieure à celle de Rhode Island, ou autrement dit, une quantité de carbone supplémentaire stockée équivalente au retrait 7.2 à 8,8 millions de voitures utilisées durant un an. Ou, à l'arrêt de toute émission par les camions, autos et motos de tout le Colorado durant plus de deux ans, selon une analyse, par région, du potentiel de séquestration du carbone si les routes inutiles étaient déconstruites et réensemencées[112]. Supprimer les routes inutiles a des avantages écologiques[113] mais aussi socio-économiques[114],[115],[116], notamment en réduisant les frais d'entretien des routes inutiles[117].
- enfin, l'analyse des feux (source : 14 ans d'imagerie satellitale [118]) de l'Est de l'Amazonie a montré que plus la fragmentation est importante, plus le risque d'incendie est élevé. De plus, l'effet lisière fait disparaitre les grands arbres et peut faire perdre à la forêt une partie significative de son carbone[119]. Un calcul (modélisation sur la base des données disponibles vers 1995) a conclu que la fragmentation de l'Amazonie brésilienne était vers 1995 à elle seule (hors carbone émis par sa destruction) responsable de 3,0 à 15,6 millions de tonnes de carbone perdues par an, et de 22 à 149 millions de tonnes par an pour l'ensemble des forêts tropicales.
Enjeux sanitaires
La dégradation des forêts peut favoriser l'extension de maladies émergentes[120] comme par la propagation du virus Ebola[121], émergent depuis 1976, du virus Marburg depuis 1967[121], de la maladie de Lyme[122] ou de la malaria : « une étude réalisée au Pérou en 2006 révèle que le taux de piqûre par les moustiques porteurs de malaria est 278 fois moins élevé dans les forêts intactes que partout ailleurs. »[123].
Enjeux spécifiques aux zones tropicales



- En présence d'un climat et/ou d'un sol difficile, la fragmentation forestière aggrave les risques d'érosion et d'incendies[118], d'érosion des sols, avec comme à Madagascar des effets irréversibles (à l'échelle humaine).
- Hormis sur certaines îles depuis longtemps déforestées (comme l'Île de Pâques) ou en cours de déforestation, la fragmentation forestière est sous les tropiques souvent récente. Lorsque le sol et sa banque de graines ont été conservés, un potentiel de recolonisation (résilience) existe, au moins pour ce qui concerne la composition en essences pionnières et végétaux héliophiles, voire en essences sciaphiles (d'ombre) lorsque les fragments sont assez grands et nombreux (Cf. potentiel mesuré en Côte d'Ivoire dans une forêt en cours de fragmentation depuis une trentaine d'années[15]. la recolonisation par la faune ou la fonge peut être plus délicate ; elle nécessite que les espèces n'aient pas disparu, or plus une forêt est morcelée, plus elle est vulnérable aux épidémies, aux introductions d'espèces invasives ou seulement envahissantes et à la surexploitation du gibier et de la viande de brousse. Les grands animaux sont souvent ceux qui disparaissent les premiers. Le recul des populations de carnivores et prédateurs augmente le risque de déséquilibre des relations prédateurs-proies.
- En 2016 J. Barlow et al (2016), sur la base des données disponibles sur les modifications du couvert forestier mondial [124],[125], confirment qu'en forêt tropicale la biodiversité est mieux conservée dans les paysages peu fragmentés par l’homme, et là où le dérangement humain est moindre (le dérangement peut doubler la perte de biodiversité liée à la déforestation tropicale[126]).
En une étude confirme que le recul mondial de la forêt naturelle érode de manière « disproportionnée » la biodiversité ; les dernière forêts et paysages intacts devraient être protégés concluent les auteurs[127]. Là où la forêt recule les vertébrés risquent plus d'être classé menacé par l'UICN ou de basculer dans une catégorie de menace plus élevée et qu'elles présentant des populations en déclin augmentent « considérablement »[127]. Et ce risques est « disproportionné » dans des hot-spots de biodiversité que sont les immenses massifs forestiers tropicaux de Bornéo, d'Amazonie centrale et Forêt du bassin du Congo[127] ; Là, même une faible déforestation (routes, pistes forestières, aires de stockage, petite urbanisation…) a de graves conséquences pour la biodiversité des vertébrés (et donc secondairement donc pour des espèces qui en dépendent). Les données montrent que la perte de forêt y est aussi grave pour la biodiversité des vertébrés que dans les zones déjà fragmentés[127] ; A ce rythme, à Bornéo, en Amazonie centrale et dans le bassin du Congo, rien que pour les vertébrés, 121 à 219 espèces rejoindront la liste des espèces menacées dans les 30 prochaines années[127]. Et les effets du changement climatique pourraient aggraver les choses, de même que la dette d’extinction (voir Dette d'extinction) Or l’artificialisation du monde s’aggrave rapidement[128]. Or seules 17,9 % de ces trois zones sont actuellement formellement protégées et moins de la moitié (8,9 %) ont une protection stricte. De nouveaux efforts de conservation et de restauration de l’intégrité écologique des forêts sont urgemment à mettre en œuvre à grande échelle (mégaréserves naturelles, réellement protégées, déjà suggérées en 2005 par C Peres[129]) « pour éviter une nouvelle vague d'extinction globale »[127].
Les effets de la fragmentation
Les effets généraux
- la connectivité entre peuplements et populations au sein d'une métapopulation (animale ou végétale) augmente la viabilité des populations (Cf. études et modèles[130],[131] et travaux expérimentaux[132],[133], [134],[135].
La perte de connectivité due à la fragmentation forestière est (directement ou indirectement) source de régressions ou disparitions d'espèces (faune, fonge, microbes, flore forestière) ou de modification de comportement des espèces forestières, notamment des petits mammifères[136].
Elle dégrade le fonctionnement des écosystèmes forestiers, notamment en changeant les espèces forestières et leurs proportions[137].
Elle appauvrit la faune[138], altère le "recrutement"[139], la richesse[140] et la diversité des forêts, au profit d'espèces banales ou invasives, et au détriment d'espèces typiques des forêts anciennes et en particulier des vieux et grands arbres[141] qui disparaissent les premiers.
Certains changements sont rapides (visibles dès les premiers mois), et perdurent longtemps[142],[143].
En Forêt comme ailleurs, chaque espèce répond différemment à la fragmentation ; selon ses exigences[144], ses traits de vie[145] mais aussi selon l'âge et la taille du fragment[146].
Les effets synergiques : plusieurs phénomènes aggravent respectivement leurs effets négatifs.
- Perte directes d'habitats viables[142](plus ou moins importante selon les cas et selon le type de matrice écopaysagère du fragment[147]). Elle est souvent définitive à échelle humaine de temps, par exemple pour les surfaces converties en route, en zone urbanisée, industrielle ou d'agriculture intensive ou en grande retenue hydraulique.
Les qualités de l'habitat typique forestier se dégradent aux abords des lisières artificielles (alors que l'effet "écotone" des lisières naturelles de clairières, berges et milieux naturels "ouverts" sont au contraire positifs pour la biodiversité). Ces effets restent cependant modestes par rapport aux autres. Les espèces arboricoles sont affectées[148] mais elles ne sont pas les seules, notamment parce que d'autres espèces en dépendent au travers d'interactions durables (pollinisation et dispersion de graines par exemple). - Création d'"îlôts forestiers" (« insularisation ») où les ressources alimentaires peuvent manquer. Même les espèces volantes sont touchées. Dans ces îlots, beaucoup d'espèces animales produisent autant de petits[149], mais la prédation y augmente, ou change de nature en milieu fragmenté[150] ; ex. : en Amérique du Nord, les corvidés et d'autres prédateurs (rapaces) augmentent leur prédation dans les nids d'oiseaux en forêt fragmentée, alors que les écureuils Tamiasciurus hudsonicus (qui mangent des œufs ou oisillons) la diminuent[151]. La nourriture des espèces forestières peut manquer plus rapidement dans les fragments de forêts ; Des populations d'oiseaux ou d'insectes, pourtant capables de voler, peuvent ainsi s'effondrer dans les parcelles fragmentées[152].
De plus, au sein d'une métapopulation d'espèce forestière, la fragmentation stoppe ou freine les flux de gènes pour les espèces "insularisées". Et, également pour d'autres raisons, les îlots forestiers ainsi formées peuvent devenir des pièges écologiques et/ou des puits écologiques.
Le milieu "ouvert" ainsi créé ne peut pas remplacer ceux que créent les chablis ou clairières naturelles (sauf, d'un certain point de vue, rares cas particuliers de "no-man's-land").
Plus ces îlots boisés relictuels sont petits et isolés les uns des autres, plus grands sont les risques de consanguinité, de dérive génétique, de perte de patrimoine génétique et donc d'extinction d'espèces.
Selon les modèles disponibles, en zone tropicale, en dessous d'un seuil de 25 ha, la structure forestière et la biodiversité sont rapidement, profondément et durablement affectées : la forêt mature ne peut réapparaitre ; seuls les premiers stades du cycle sylvogénétique sont maintenus[153].
C'est en forêt de montagne, à cause des pentes impliquant des routes en lacets, que le linéaire de routes est proportionnellement le plus long. Or, c'est là aussi ; grâce à des microclimats variés induits par l'altitude et le relief (⇒ expositions variées) que la diversité biologique est la plus variée sur de courtes distances géographiques. La biodiversité pourrait y être plus vulnérable encore à la fragmentation ; avec des pertes considérables dans la forêt des nuages[154]. - le morcellement détruit ou limite l'intégrité écologique (d'une forêt, d'un massif…), au détriment de la résilience écologique de la forêt.
Les cœurs de tous les grands massifs boisés n'étaient autrefois accessible qu'au prix de beaucoup d'effort, en pirogue ou après de longs trajets en forêt). La même forêt aujourd’hui fragmentée par un nombre croissant de routes, pistes et layons est devenue aisément pénétrable ; Ceci y facilite la chasse et le braconnage tout en exacerbant leurs effets[155],[156]. La déforestation ou la surexploitation du bois et de ressources forestières autres que le bois (gibier, fruits, plantes, champignons…) sont également favorisées (y compris éventuellement par les populations autochtones[157]), précisément là où ces ressources étaient autrefois naturellement mieux protégées. La fragmentation freine également la restauration naturelle des populations de gibier ou plantes exploitées.
Les lisières et tous les petits "patchs" de forêt sont aussi plus vulnérables à l'introduction d'espèces invasives ou de certains pathogènes ainsi qu'à la déhsydratation et donc aux incendies.
- Ces trois facteurs sont sources de déséquilibres écologiques. Ils dégradent tout ou partie des services écosystémiques rendus par les forêts naturelles ou subnaturelles. Dans les cas les plus graves, ils aboutissent à la disparition même de la forêt.
Altération de l'ambiance forestière
La fragmentation augmente la pénétration de la lumière, des sons, du vent et modifie les températures, l'hygrométrie et les odeurs, surtout à proximité de la zone de "coupure" où l'effet-lisière y est maximal ;
Ces paramètres - au-delà de certains seuils - sont de discrets mais puissants facteurs limitants pour les espèces typiquement forestières. Leur altération crée un effet barrière ou de répulsion qui s'étend plus ou moins loin dans les marges intérieures des fragments de forêt (quelques mètres à quelques centaines de mètres).
Modification de la flore, du nombre d'essences et de leur répartition
Les modifications de la diversité floristique et la répartition sont les mieux étudiées. Dans les fragments forestiers, la lumière pénètre mieux, favorisant le passage du stade pousse au stade subadulte essentiellement pour les essences pionnières et de lumière (héliophiles) au détriment des espèces d'ombres (sciaphiles)[15](voir dryades). Lorsque la lisière se densifie (en quelques mois ou quelques années en présence de lianes à croissance rapide), le phénomène est freiné, en particulier lorsque la canopée reste fermée au-dessus du fragment (cas d'une piste forestière étroite par exemple).
La forme du fragment influe sur la longueur de son périmètre. S'il est digité et long, l'implantation d'héliophiles et d'éventuelles espèces invasives ou opportunistes est favorisé. La dynamique du peuplement de régénération est stimulée par la lumière et une moindre concurrence, pour partie comme sur les limites d'un chablis naturel[15]. Cet effet est marqué dans les fragments récemment constitués[15] et de forme digitée[15] (plus de lumière). La richesse spécifique et la diversité de la régénération peuvent apparaître plus importantes dans ces fragments[15] car il s'y ajoute des espèces de la matrice et des lisières. Les espèces de cœur d'habitat forestier, presque toujours sciaphiles, peuvent cependant disparaitre ou être fortement inhibées. « Une érosion progressive du potentiel de régénération des fragments apparaît avec la durée de l’isolation. Au sein des fragments de faible surface, cette érosion est accentuée »[15]. Les espèces pionnières sont d'autant plus présentes que des perturbations de la structure végétale (trouées, feux, arasements, terrassements, etc.) accompagnent le processus de fragmentation. Les espèces dominantes changent très rapidement ; celles dont la dispersion est aérienne (anémochores et ornithochores) prolifèrent, surtout en bordure des fragments au détriment de celles dont la dispersion ou la germination nécessitent la présence de mammifères [15]. En Côte d'Ivoire« l'étude de la composition floristique totale menée dans les fragments de moins de 4 hectares a mis en évidence une série d'espèces pour lesquelles la régénération est très compromise »[15].
Les lisières artificielles et l'accès facilité aux cœurs de forêt peuvent en outre faciliter l'introduction d'espèces invasives comme le cerisier tardif ou l'ailanthe en Franc. La biodiversité floristique du sous-bois est « nettement plus pauvre et composée d’espèces plus banales sous A. altissima que sous les autres espèces d’arbres et la composition floristique est sensiblement différente[158]. En outre, la densité de drageons d’A. altissima dans les aires d’inventaires est significativement négativement corrélée avec la richesse floristique ». Les impacts croisés de la compétition interspécifique et les propriétés allélopathiques d’A. altissima pourraient expliquer ce double phénomène[158].
La fragmentation et souvent associé à un rajeunissement de la forêt, par une forêt secondaire qui se traduit par une perte parfois très importante de biodiversité floristique[159], ainsi que sur les champignons, notamment lorsque la forêt tend à devenir monospécifique[160]. L'impact du passage de l'homme peut rester visible des siècles durant, voire des millénaires[161],[162].
Modification de la qualité et de la nature de la "pluie de spores et de graines", et mauvaise régénération
La "pluie" de spores et graines est une composante essentielle de la régénération forestière, dont pour les épiphytes. Certaines espèces ont une production irrégulière de graines (ex. les glandées des chênes).
On pourrait croire que près des lisières où le vent est plus actif et la lumière plus disponible, la dispersion des semence est plus active. Ceci n'est vrai que pour un faible pourcentage des espèces forestières (souvent des pionnières et ubiquistes, dont les graines sont légères et résistantes). Les études faites au moyen de "pièges à graines" montrent au contraire que la pluie de spores (de mousses, lichens[163], fougère champignons) et la pluie de graines sont pour la plupart des espèces forestières négativement affectées par la proximité des lisières (et donc par la fragmentation)[163].
- Il a ainsi été (en 2006) démontré[164] qu'en lisière artificielle et en bordure de parcelles fragmentées, la « pluie de graines » diffère fortement de ce qu'elle est dans la forêt continue, en ce qui concerne l'abondance de graines, la variété et la richesse en espèces, le mode de dispersion, et même la taille des graines ou encore la manipulation, le transport, le stockage, l'enfouissement, la digestion par des vertébrés. Par exemple, dans un massif de 3 500 ha de forêt atlantique du nord-est du Brésil, la pluie de graines a été étudiée sur un transect de 1,5 km (à partir de 76207 semences de 146 espèces recueillis sur 100 points dans différents habitats)[164] ; cette "pluie" s'est montrée fortement modifiée par la fragmentation : dans le noyau de forêt dense, les graines moyennes, grandes et très grandes étaient près de deux fois plus nombreuses (21,8 % du total) que près des lisières (13,5 % du total). De même le nombre d'espèces représentées (82,6 % des espèces) est bien plus élevé que près des lisières (57,5 % des espèces)[164]. De plus, les semences de grandes et très grande tailles (correspondant souvent aux grands arbres) étaient beaucoup plus rarement manipulées par des vertébrés près des lisières (13,04 % de toutes les espèces connues dans cette zone pour leur zoochorie par des vertébrés, versus 31,5 % en intérieur de forêts)[164]. Enfin, les semences et fruits « manipulés » par les vertébrés ne représentaient que 5,9 % de toutes les graines du bord de la forêt (versus 11 % à l'intérieur des forêts)[164]. La fragmentation en modifiant le taux de grosses graines et le taux de dispersion des graines aura des impacts à moyen et long terme, encore peu perceptibles pour les espèces longévives, mais probablement importants, et qui peuvent en partie expliquer certains changements floristiques observés près des lisières artificielles. Certaines espèces de vertébrés pourraient également en pâtir (quand leur nourriture dépend de grosses graines par exemple)
De plus le microclimat plus déshydratant des lisières artificielles empêche la fructification et/ou la germination de nombreuses épiphytes qui sont également des abris, des habitats, des moyens de circulation (dans le cas des lianes) et des sources de nourritures pour diverses espèces.
Une autre étude a montré que la régénération en est affectée[165] - Au Nord-Est du Brésil, en 2012[166], les forêts atlantiques secondaires sont comparées aux restes périphériques (3 500 ha) d’une forêt primaire atlantique encore épargnée et non fragmentée (dans laquelle elles sont incluses).
Les parcelles de forêt secondaire étaient fragmentées et âgées de 19 à 62 ans. Environ 400 000 semences de 180 morphospecies y ont été récoltées dans 60 pièges à graines de 1 m2 pour étudier la pluie de graine[166].
Résultats : à partir du 1er stade (régénération et pionnières) jusqu’au stade d’une soixantaine d’année, la pluie de graines diminue en densité mais augmente en nombre d’espèces. La pluie de graines se différencie spatialement lorsque la forêt devient plus mature[166]. Enfin, l’étude montre que dans cette forêt, les vertébrés (mammifères, oiseaux) jouent un rôle essentiel dans la régénération du couvert forestier par zoochorie : En effet, quel qu’était l'âge du peuplement secondaire étudié, les graines dispersées par des vertébrés prédominaient toujours largement (67 à 75 % de toutes les espèces enregistrées)[166]. Par contre, les grosses graines (typiques de la forêt ancienne) n’y représentaient que 5 à 8 % des espèces recensées dans la pluie de graines, soit 5 fois moins que dans les forêts anciennes périphériques (où elles sont 31 % des graines tombant sous la canopée)[166]. L’étude conclut que ces forêts secondaires, bien qu’incorporées dans les fragments les plus grands et les mieux conservés de la forêt atlantique très fragmentée du Nord-Est du Brésil, risquent de ne pas pouvoir atteindre les derniers stades de la succession, faute de grosses graines (celles de la forêt mature), et que la pire situation est celle des parcelles les plus isolées et les plus petites ; même protégées, elles pourraient perdre leur capacité à fournir la plupart des services écosystémiques fournis par la forêt originelle[166].
Déclin de la biomasse
La biomasse décline dans la forêt amazonienne fragmentée, avec parfois un véritable effondrement ("Collapse"[167]) ; Au centre de la forêt pluvieuse, la biomasse aérienne s'effondre, semble-t-il sans être compensée par le recrutement de nouveaux arbres ; Les "pertes" sont les plus importantes « dans les 100 mètres de bords de fragments, où la mortalité des arbres est fortement augmenté par les changements microclimatiques et les turbulences plus élevées du vent ». Les parcelles d'étude permanents dans la bande de 100 mètres où l'effet lisière est le plus marqué « ont perdu jusqu'à 36 % de leur biomasse dans les 10 à 17 ans qui ont suivi la fragmentation ». On constate une expansion des lianes ligneuses sur la lisière, mais « qui ne compense qu'une faible partie de la biomasse perdue par la mortalité accrue des arbres ». Le puits de carbone et le stock de carbone forestier en sont affectés. Les phénomènes d'érosion ou d'incendie peuvent suivre et encore aggraver le phénomène, mais la fragmentation est dans ce cas la première cause de perte de biomasse[168]. David Cowles signale[169] que la mesure classique de la biomasse sur pied par la surface terrière présente un léger biais ; elle peut conduire à sous-estimer l'importance des branches épicormiques et « gourmands » qui poussent vers la lumière en lisière et du recru en ne prenant pas en compte les pousses de moins de 10 cm de diamètre. Il est donc important d'étudier l'effet à long terme de la fragmentation, le temps que le recru (jeunes pousses) puisse s'exprimer (ce qui est plus long en zone tempérée et plus encore en zone froide qu'en zone tropicale). Une autre étude a comparé des fragments de 1 à 50 ha isolés de la forêt amazonienne depuis 10 à 19 ans à la forêt originelle. Cette étude a mesuré « la quasi-totalité du matériel végétal vivant ou mort (grands arbres, semis, plants, petits arbres, palmiers, lianes, vieux bois et bois mort, y compris abattu, chicots, litière). » et elle a confirmé ce phénomène. Cette étude a également montré que « étonnamment, le « turnover » annuel estimé de débris de bois augmente significativement près des lisières artificielles de forêts, ce qui suggère que la décomposition se produit plus rapidement dans les forêts fragmentées que dans les forêts continues », donnée nouvelle à prendre en compte dans le cycle du carbone et l'évaluation des puits de carbone forestiers et les émissions de CO2 et de méthane à partir des forêts[170],[171]. Il semble en être de même en forêt ancienne tempérée (ex : fortes densité de bois-mort, malgré une biomasse également plus élevée en Forêt de Białowieża).
Le déclin des arbres émergents de la canopée
Dans le cas de la forêt atlantique (mata atlântica) du Nord-Est du Brésil, une étude (2008[172]) a comparé l’état et le nombre des très grands arbres (large treesou arbres émergents), dans la forêt ancienne et dans 58 fragments (déjà anciens de 3,4 à 295,7 ha). Les espèces comptabilisées sous forme de surface terrière (toutes les tiges de diamètre ≥ 10 cm) montre que celle-ci est divisée environ par deux dans les lisières de forêt ancienne et dans les fragments[172].
Les individus très hauts et larges ont presque totalement disparu de la forêt fragmentée (0,24 ± 0,27 % de toutes les tiges échantillonnées) et totalement disparu des lisières. Alors que les très grands arbres dépassent toujours 1,5 % des arbres dans les cœurs intérieurs de la forêt la moins fragmentée (2,9 ± 2,8 %).
En outre, la fragmentation forestière modifie aussi l’architecture des grands arbres : pour des raisons mal comprises, leur surface terrière individuelle est en moyenne diminuée de 30 % dans les petits fragments et les lisières de forêt ancienne (ce qui en fait a priori des arbres plus fragiles)[172].
Enfin, une analyse par espèces bioindicatrices a mis en évidence un groupe écologique de douze espèces de grands arbres significativement associés aux cœurs de forêt mais totalement absentes des habitats concernés par un effet de bordure[172].
Les auteurs concluent à « un appauvrissement persistante et substantiel des grandes espèces émergeant de la canopée, incluant d’un effondrement structurel (structural collapse) de la strate sous-jacente émergente (forest emergent layer) », dans les parties fragmentées ou hyper-fragmentées de la forêt atlantique brésilienne[172].
Vulnérabilité accrue de la forêt fragmentée
Les effets des tempêtes, cyclones tropicaux et orages sont accrus en forêt fragmentées, notamment en zone tropicale[173].
Déclin fonctionnel des écosystèmes forestiers fragmentés

Un habitat forestier fragmenté voit son fonctionnement dégradé ; avec des effets indirects inévitables. Ces effets varient selon l'intensité de la perturbation et selon les "fonctions écologiques" remplies par les espèces les plus perturbées.
Sur toute la planète, les scientifiques constatent des phénomènes de synergies[175] qui font que la fragmentation des forêts, via notamment des "effets de bordure"[176] induit une dégradation des écosystèmes forestiers. Les espèces typiquement forestières régressent le plus mais la productivité globale de l'écosystème se dégrade aussi[177]. La banalisation de la flore des lisières artificielles diminue aussi l'offre alimentaire des animaux[178], conduisant parfois à leur disparition[175].
- Par exemple, au Brésil, neuf attributs du fonctionnement écologique de la forêt atlantique (Mata Atlântica) ont été décrits dans de quatre types d'habitats ; 1. au cœur d'un morceau de forêt primaire de 3 500 ha d'un seul tenant ; 2. dans des habitats de lisière ; 3. dans des parcelles de forêt secondaire et 4. dans de petits fragments relictuels de forêt primaire (de 3,4 à 79,6 ha chacun)[179].
Les résultats ont confirmé l'hypothèse que le fonctionnement de la forêt morcelée était « bien plus semblable » à celui d'un « boisement secondaire » de 5 à 65 ans qu’à celui de patchs étudiés en cœur de forêt naturelle. Il en allait de même pour les zones proches des lisières dans les « très grands fragments »[179]. Les facteurs les plus affectés par la fragmentation étaient la richesse spécifique, la proportion d’essences pionnières, la part des arbres à grosses graines et le taux d'espèces émergent au-dessus de la canopée (d’autant plus réduits que le fragment était petit ou que la placette étudiée était proche d’une lisière de grande parcelle)[179]. L’âge des peuplements de seconde venue expliquait de 31,4 % à 88,2 % des variations des attributs fonctionnels des populations d'arbre. La plupart des traits écologiques caractérisant le fonctionnement de la bande forestière soumise à l'effet-lisière correspondaient au fonctionnement d'un jeune boisement secondaire (de moins de 25 ans)[179] et non à l'âge véritable de la forêt. Les auteurs ont aussi constaté que les traits associés aux petits fragments de forêt évoquaient bien ceux d’un boisement d'âge intermédiaire (25-45 ans) d’un peuplement forestier secondaire[179].
De plus, alors que le type d'habitat avait une influence sur les attributs étudiés, le type de sol, le type de végétation, ou encore la localisation spatiale des parcelles étudiées étaient ici sans influence (sur ces 9 attributs) [179].
Une analyse multivariée a mis en évidence « une dérive floristique saisissante (« striking floristic drift ») au sein des habitats de lisières touchés par l’effet-bordure »[179].
Les auteurs en ont conclu que les actuelles « lignes directrices des politiques de conservation échoueront à protéger les paysages de forêt ancienne hyper-fragmentés d’un appauvrissement drastique tant que les taches de forêt relictuelle seront fortement dominées par les habitats de lisière »[179]. - Un double constat est fait dans le monde entier :
les forêts exploitées les plus fragmentées subissent un recul préoccupant de leur contenu en bois mort[180].
De plus, la dégradation du bois mort y est anormalement rapide[180].
Les grands et vieux arbres y disparaissent d'autant plus vite que la fragmentation est importante[180]. Or, en zone tropicale, ces grands arbres ont une influence positive (« disproportionnée » sur la structure et le fonctionnement de la forêt, car plus denses, plus longévifs, produisant le plus de graines, offrant une diversité plus élevée de perchoirs et micro-habitats aux oiseaux, insectes, épiphytes, champignons et produisant les gros bois-morts nécessaires à la vie de milliers d'insectes saproxylophages.
Les effets à moyens et long terme
La vulnérabilité des forêts se mesure selon les effets de court terme, mais aussi de moyen et long termes.
L'étude de fragments anciens de forêts de quelques décennies ou siècles démontrent des effets à long terme en forêt tempérée comme tropicale[181],[182],[183] comme en forêt tempérée[184]. Une mosaïque de fragments forestiers ne peut conserver les mêmes fonctions et la même biodiversité d'une forêt non fragmentée et les effets de lisière sont alors négatifs[185].
L'actuel réchauffement climatique peut exacerber ces effets, en augmentant par exemple le risque d'incendie de forêt ; la fragmentation anthropique des paysages interrompt des corridors climatiques permettant normalement une migration des espèces vers les pôles ou en altitude, plus frais alors que le climat se réchauffe.
Ceci peut conduire à une moindre résilience écologique et à la disparition de nombreuses espèces[186].
La forêt galerie et les ripisylves sont des écotones et corridors forestiers importants ; linéaires, dépourvus de zone-centrale et relativement étroits, et bien que théoriquement plus résistants à une diminution de la pluviométrie en saison chaude lorsqu'ils ne sont pas accolés à une forêt, ils se montrent appauvris en espèces typiquement forestières et très vulnérables à la fragmentation[187].
La dispersion et la germination des graines participant à la régénération des fragments forestiers sont profondément modifiées à moyen et long terme (Benitez-Malvido, 1998 ; Laurance et al., 1998 a ; Viana et al., 1997) , affectant la composition locale (et régionale[188],[189]) de la distribution et affectant la qualité de la régénération ; deux facteurs qui déterminent l'évolution et l’adaptation future des communautés forestières.
Si à échelle locale on voit apparaître de nouvelles espèces (de milieux ouverts), dans la forêt fragmentée, globalement, les communautés se réorganisent en s'appauvrissant[190]. Les oiseaux chanteurs ont une vulnérabilité particulière à la fragmentation forestière selon une relation non linéaire, c'est-à-dire caractérisée par un seuil de superficie du fragment sous lequel l'espèce régresse brutalement[191], avec une sensibilité variant aussi selon les espèces)[191]. En Suède, une étude a montré que pour certaines espèces capables de continuer à se reproduire un certain temps de manière végétative ou par autopollinisation, leur disparition ou régression peut ne se manifester qu'après 50 ou 100 ans (décalage temporel de la modification du faciès paysager [192] plus ou moins lent selon les traits de vie de l'espèce[193]
La gravité de la fragmentation



Plus de la moitié des forêts du monde sont sous influence directe de lisières artificielles (qui se comptent en millions de kilomètres). Une base de données spéciale a été créée pour mieux suivre leur évolution [194] (Modéliser et prédire les effets d’une lisière dans des paysages de plus en plus fragmentés et hétérogènes et anthropisés et soumis au dérèglement climatique est très difficile mais une (autre) base de données vise à aider les chercheurs à prédire la réponse de la biodiversité au impacts de l'Homme[195]). Quelques espèces plutôt ubiquistes profitent des lisières et beaucoup d’autres en pâtissent. Une étude (2017) a statistiquement compilé un vaste ensemble de données mondiales sur les réponses (en termes d’abondance) de 1 673 espèces de vertébrés à la fragmentation[196]. Selon les données disponibles, au minimum 85 % des espèces forestières ou paraforestières sont affectées par les lisières (positivement ou négativement)[196]. Comme on le sait depuis longtemps, et logiquement ce sont les espèces de la forêt profonde qui en pâtissent le plus, tout comme (cette information est plus nouvelle) celles classées comme « menacées » sur la liste rouge de l’UICN[196]. Ces espèces se montrent moins rares à 200-400 m d'altitude où la fragmentation diminue[196]. La taille des espèces semble influer sur leur vulnérabilité à la fragmentation, mais différemment selon les groupes d’espèces considéré : ainsi les populations d’espèces de petits amphibiens, de grands reptiles et les mammifères non volants de taille moyenne souffrent le plus d’une perte d’habitat non fragmenté (par rapport aux autres espèces « de cœur de forêt »). Selon les auteurs, les lisières forestières tendent à restructurer les communautés écologiques jusqu’à l'échelle mondiale[196]. Et selon Haddad & al. (2015) les impacts de la fragmentation sont planétaires, mais aussi durables dans le temps[197].
L'importance de la fragmentation varie en fonction de nombreux paramètres. Que la fragmentation forestière ait des effets sur l'écosystème forestier est largement admis, en revanche, l'ampleur et la gravité de certains effets sont encore discutées ou mal cernées comme :
- les impacts sur certains groupes (bactéries, microbes, invertébrés de la pédofaune, champignons…) ;
- les effets à long terme ou très long terme. Des biais statistiques et d'observation existent y compris pour des espèces faciles à observer comme les arbres les plus longévifs de durée de vie pluriséculaire ou millénaire en tant qu'individus dans une parcelle isolée mais en étant plus capables de se reproduire ; par exemple pour une espèce dont la graine ne germe qu'après avoir été enterrée ou digérée par un animal éteint ou qui aurait définitivement disparu de la région ou dans le cas des baobabs de Madagascar en contexte d'ouverture croissante du paysage mais en situation de dépérissement car plus dans son écosystème naturel, la forêt sèche. Pour certaines plantes, la reproduction végétative permet également une survie prolongée, mais sans échanges génétiques.
Quelques facteurs aggravants connus :
- l'intensité (efficacité) de la fragmentation pour la faune terrestre forestière non volante :
- les grands canaux bordés de berges verticales abruptes et artificielles sont très fragmentants mais les autoroutes et voies ferrées clôturées s'avèrent aussi très fragmentantes[198];
- la largeur de l'infrastructure et des abords (d'après Baggley, 1998) ;
- le trafic[199],
- les matériaux utilisés ; le type de bas-côté, la présence ou non d'une haie, de clôtures ou d'un fossé, la largeur de la coupure et portée de ses effets (sous forme de dérangement, bruit, odeur, etc.), type de clôture et/ou d'effet bordure ; orientation de la coupure (par rapport au vent, au soleil, par rapport aux flux de migration antérieurs à la fragmentation) ; pollution lumineuse et autres effets de fragmentation physique et micro-climatique.
- l'étendue de l'aire fragmentée ; la fragmentation globale d'un massif forestier est a priori plus grave lorsque tout un massif forestier ou tout un bassin versant est concerné, ou dans le cas d'une petite île, ou encore lorsque ce massif forestier est déjà petit et/ou éloigné des autres massifs les plus proches ;
- une fragmentation touchant un point critique du fonctionnement écopaysager (une zone de connexion biologique ou importante pour le cycle de l'eau par exemple) ;
L'impact d'une coupure augmente (dans l'espace et dans le temps) si elle intersecte un corridor de migration (cas fréquent dans les vallées, cols, ou certains détroits qui sont des lieux de passages obligés pour de nombreux migrateurs volants ; oiseaux, papillons, chauve-souris…)
Ainsi, le canal de Suez (en remplaçant l'antique Canal des pharaons qui n'était utilisable qu'en hautes-eaux) a fait de l'Afrique une île en l'isolant du Moyen-Orient ; creusé dans une zone aride, il n'a pas a priori fragmenté de forêt. Au contraire, plus récemment, le creusement du Canal de Panama a physiquement sectionné l'isthme de Panama et détruit sa fonction de connexion dans l'immense « corridor biologique panaméricain » qui reliait depuis plus de trente millions d'années (pliocène) l'Amérique du Nord à l'Amérique du Sud. Hormis pour certains oiseaux, poissons et insectes volants, ce canal a fonctionnellement coupé un continent (l'Amérique) en deux, interrompant brutalement ce que les biologistes ont nommé « le grand échange interaméricain ». Ailleurs et plus discrètement, des centaines de milliers de routes, canaux et voies ferrées clôturées (type TGV en France) ont - à leur échelle - de tels impacts ;- la proximité et l'importance d'un effet-lisière[179] ;
- la mauvaise santé (ou distance par rapport à l' « écopotentialité » de la forêt concernée ; apprécié au regard du contexte biogéographique (La forêt morcelée est-elle par exemple connectée par des corridors forestiers naturels à d'autres massifs plus grands ou plus riches ? ce qui pourrait peut-être améliorer sa résilience) ;
- la défaunation, qui peut aggraver l'effet d'une fragmentation physique ; car de nombreux arbres, plantes et autres organismes ont des graines ou propagules transportées par des animaux (zoochorie) et parfois même exclusivement « plantées par certains animaux » (dans leurs excréments pour des graines résistantes à la digestion, ou dans des cachettes oubliées (cas du geai des chênes ou de l'écureuil en France).
De plus, les grands carnivores jouent un rôle essentiel dans les équilibres écologiques.
- L'histoire de la forêt fragmentée joue un rôle (Quelle était sa richesse initiale ? La banque de graines du sol est elle importante ? De quand date le morcellement ? avec quelle nature, réversibilité ou gravité de fragmentation ?).
Les forêts anciennes y perdent proportionnellement le plus, car leurs arbres et plantes sont moins capables de coloniser de nouveaux sites forestiers[159], ont une croissance lente, de faibles capacités de dispersion (beaucoup ont une stratégie de dispersion à courte distance[159], ou nécessitent la présence de certains animaux), elles une production de diaspore faible ou irrégulière qui pose des problèmes de recrutement, et une faible capacité compétitive[159] ; - Les milieux associés sont aussi à considérer. Par exemple, un bocage très ancien et densément maillé a pu conserver une partie de la biodiversité d'anciennes clairières forestières, mais en France, les remembrements ont fait reculer ce bocage dans les années 1970-80.
- Il existe une relation entre fragmentation et perte de biodiversité, de type non linéaire[200] (c'est-à-dire que la biodiversité ne régresse pas régulièrement mais brutalement quand certains seuils sont atteints).
La recherche de tels seuils écologiques a été faite (2010) dans les boisements fragmentés d'une zone périurbaine de Lombardie (Italie du Nord), à partir d'indicateurs choisis dans la strate herbacée : plantes vivaces typiques de trois syntaxons phytosociologiques (Fagetalia sylvaticae, Carpinion betuli et Erythronio-Carpinion), et de trois caractéristiques de fragmentation des boisements (localisation, degré d'insularisation, forme). Les facteurs écologiques ont été évalués au moyen des indices de Landolt L et H ; obtenus à partir de relevés antérieurs faits dans des forêts de référence jugées de haute qualité écologique. Certaines herbacées vivaces indiquent des forêts anciennes, et d'autres des forêts très fragmentées[200]. Les modèles linéaires généralisés construits sur la base des relevés faits en forêt montrent une corrélation entre le nombre d'indicateurs de forêt ancienne et la taille des fragments, ainsi qu'avec le degré de connectivité de ces fragments avec d'autres milieux forestiers[200]. Les modèles additifs généralisés confirment une relation non linéaire entre l’indicateur de richesse et la taille des fragments ; avec présence de 2 seuils[200] : 1) Le nombre d'indicateurs forestiers a augmenté jusqu'à un seuil de 35-40 ha ; au-dessus de cette taille, la richesse est restée constante (Dans cette région italienne, il faudrait donc protéger les fragments de forêt atteignant au moins 35 à 40 ha d’un seul tenant). 2) L'autre seuil concerne la relation entre l'aire et le nombre d'indicateurs ; cette relation est moins prononcée dans les fragments de moins de 1 à 1,5 ha. Donc, si les fragments de taille comprise entre ces 2 seuils sont agrandis, leur valeur pour la biodiversité végétale grandira fortement.
L'existence de ces deux seuils suggère une réponse brutale de la flore à certains changements écopaysagers[200]. Elle fournit aussi des objectifs quantitatifs en matière de conservation[200]. Si la Lombardie veut conserver (ou restaurer) sa biodiversité forestière, elle devrait - avant d'atteindre des seuils critiques - défragmenter les forêts périurbaines[200]. Les herbacées vivaces de forêts anciennes ou typiquement associées aux forêts hyperfragmentées semblent être des indicateurs fiables pour l’évaluation environnementale, mais ils varient selon chaque zone biogéographique, concluent les auteurs[200].
Voir aussi
Articles connexes
- Forêt
- Paysage forestier intact
- Écologie du paysage
- Fragmentation écopaysagère
- Intégrité écologique
- Trame verte
- Route HQE, Route forestière
- Écoduc
- Changement d'affectation des sols
- Connectivité écologique
- Corridor biologique
- Mortalité animale due aux véhicules
- Migration animale
- Cloisonnement forestier
- Biologie de la conservation
Bibliographie
- (en) Brocke RH, O’Pezio JP & Gustafson KA (1988) « A forest management scheme mitigating impact of road networks on sensitive wildlife species » dans Richard M. DeGraaf et William M Healy, dir., Is Forest Fragmentation a Management Issue in the Northeast ?, USDA Forest Service, Northeastern Forest Experiment Station, Rochester, 1988, à la p. 13, (en ligne : www.treesearch.fs.fed.us <http://www.treesearch.fs.fed.us/pubs/4191>
- (en) Debinski D.M (2006) Forest fragmentation and matrix effects: the matrix does matter. J. Biogeogr. 33, 1791–179
- (fr) Desmarais M (2012) Le droit applicable aux chemins forestiers du Québec dans une perspective de protection de la biodiversité (Doctoral dissertation, Université Laval).
- (en) Fletcher R.J Jr. (2005) Multiple edge effects and their implications in fragmented landscapes. J. Anim. Ecol. 74, 342–352
- (en) Kevin A. Hovel, Helen M. Regan, 2008 - Using an individual-based model to examine the roles of habitat fragmentation and behavior on predator–prey relationships in seagrass landscapes. Landscape Ecology 23:S1, 75-89 ; 1-Jan-2008 (résumé)
- Lauric C (2008) Quels indicateurs pour évaluer la qualité de sols forestiers soumis à des contraintes environnementales fortes ? Thèse de doctorat de l'université de Grenoble, CEMAGREF, sciences du sol, 245 p.
- (fr) NatureParif, Fragmentation forestière par les infrastructures de transport. IAU Île-de-France, Colloque 2011
- Pfeifer M & al. (2014) BIOFRAG — a new database for analyzing BIOdiversity responses to forest FRAGmentation. Ecol. Evol. 4, 1524–1537
- (en)ScienceDirect ; voir base de données liste d'articles scientifiques relatifs à la fragmentation forestière et aux puits de carbone ou pertes de carbone (745 articles relatif à la fragmentation forestière (forest fragmentation) le 14/12/2012)
- (fr) Vallauri Daniel, André Jean, Blondel Jacques, Le bois mort, une lacune des forêts gérées = Dead wood - a typical shortcoming of managed forests Revue forestière française ; (ISSN 0035-2829), 2003, vol. 55, no2, p. 99-112 ; Ed:ENGREFF, 14 p. (résumé Inist/CNRS)
- Vancutsem Ch. & Achard F (2016) Mapping Intact and Degraded Humid Forests over the Tropical Belt From 32 Years Of Landsat Time Series |Paper 2034 - Session title: Tropical Forest and REDD+ 1. ESA living planet symposium 2016
- (en) Vetter D, Hansbauer M.M, Végvári Z & Storch I (2011) Predictors of forest fragmentation sensitivity in Neotropical vertebrates: a quantitative review. Ecography 34, 1–8 |résumé
- (en) Vivek Shandas, Marina Alberti, 2009 - Exploring the role of vegetation fragmentation on aquatic conditions: Linking upland with riparian areas in Puget Sound lowland streams. Landscape and Urban Planning 90:1-2, 66-75, en ligne te: 15-Mar-2009. (résumé)
Bases de données
- Pfeifer M. & al. (2014) BIOFRAG : a new database for analyzing BIOdiversity responses to forest FRAGmentation. Ecol. Evol. 4, 1524–1537
Références
- Pahari K., Murai S., 1999. Modelling for prediction of global deforestation based on the growth of human population. Journal of Photogrammetry and Remote Sensing 54: 317–324
- Riitters K.H., Wickham J.D., 2003. How far to the nearest road ? Frontiers in Ecology and the Environment 1: 125–129
- Fahrig L., 2003. Effects of habitat fragmentation on biodiversity. Annual review of Ecology, Evolution, and Systematics 34 : 487-515
- Bierregaard R., Gascon C., Lovejoy T.E., Mesquita R., 2001 - Lessons from Amazonia: The Ecology and Conservation of a Fragmented Forest ; (ISBN 0-300-08483-8) (Contenu, prologue, préface)
- Gascon C., Lovejoy T.E., 1998. Ecological impacts of forest fragmentation in central Amazonia. Zoology 101: 273–280
- Carvalho K.S., Vasconcelos H.L., 1999. Forest fragmentation in central Amazonia and its effects on litter-dwelling ants. Biological Conservation 91: 151–157
- Laurance W.F., H.L. Vasconcelos, T.E. Lovejoy, 2000. Forest loss and fragmentation in the Amazon: implications for wildlife conservation. Oryx 34: 39–45
- Kurki S., A. Nikula, P. Helle, H. Linden, 2000. Landscape fragmentation and forest composition effects on grouse breeding success in boreal forests. Ecology 81:1985–1997
- Jules E.S., E.J. Frost, L.S. Mills, D.A. Tallmon, 1999. Ecological consequences of forest fragmentation in the Klamath region. Natural Areas Journal 19: 368–378
- Riitters K.H., J.D. Wickham, R.V. O’Neill, K.B. Jones, E.R. Smith, J.W. Coulston, T.G. Wade, J.H. Smith, 2002. Fragmentation of continental United States forests. Ecosystems 5: 815–822
- Semazzi F.H.M., S. Yi, 2001 - A GCM study of climate change induced by deforestation in Africa. Climate Research 17: 169–182
- Zhang H., Henderson-Sellers A., McGuffie K., 2001 - The compounding effects of tropical deforestation and greenhouse warming on climate. Climatic Change 49: 309–338
- Wade T.G., Riitters K.H , Wickham J.D. , Jones K.B., 2003 - Distribution and causes of global forest fragmentation. Conserv. Ecol. 7, 2: 7
- Saunders D.A., Hobbs R.J., Margules C.R., 1991 - Biological consequences of ecosystem fragmentation: a review. Conserv. Biol. 5: 18-32
- Martin Pascal, Influence de la fragmentation forestière sur la régénération des espèces arborées dans le Sud-Ouest de la Côte d'Ivoire (PDF, 10 MB), Thèse de doctorat : Université de Genève, 2008. - Sc. 3989. 2008/06/03 (et Résumé)
- Sih A., Jonsson B.G., Luikart G., 2000 - Habitat loss: ecological, evolutionary and genetic consequences. Trends in Ecology & Evolution ; 15: 132–134
- Franklin J.F., Lindenmayer D.B., 2009 - Importance of matrix habitats in maintaining biological diversity. Proc. Natl. Acad. Sci. U.S.A. 106(2): 349-350 (résumé)
- Ontario M.N.R., 2000 - Conserving the forest interior : a threatened wildlife habitat, LandOwner Resource Centre, PDF, 12 p, (ISSN 1198-3744)
- Lemieux E., 2009 - Rapport d'analyse de fragmentation du couvert forestier et analyse des forêts d’intérieur (Le Bourdon) ; UAF 06451 ; Association des pourvoiries des Laurentides, 13 juillet 2010, consulté 2012-06-10
- Dufault D., 2007 - Enclaves forestières décidues en matrice agricole et activité reproductrice d'une espèce aviaire sensible aux conditions d'intérieur de forêt. Mémoire de maîtrise, Université du Québec à Montréal
- Porneluzi P.A., Faaborg J., 1999 - Season-long fecundity, survival, and viability of Ovenbirds in fragmented and unfragmented landscapes. Conservation Biology 13 : 1151-1161
- Faaborg, J. M., Brittingham, T. Donovan, and J. Blake. 1993. Habitat fragmentation in thetemperate zone: A perspective for managers. In Status and Management of FeotropicalMigratory Birds, ed. D.M. Finch and P.W. Stangel, 331-38. US Forest Service GeneralTechnical Report RM-229
- Potapov P., Yaroshenko A., Turubanova S., Dubinin M., Laestadius L., Thies C., Aksenov D., Egorov A., Yesipova Y., Glushkov I., Karpachevskiy M., Kostikova A., Manisha A., Tsybikova E., Zhuravleva I. (2008) [Mapping the World’s Intact Forest Landscapes by Remote Sensing]. Ecology and Society 13 (2):51
- Watson M.L., 2005 - Habitat fragmentation and the effects of roads on wildlife and habitats. Background and Literature Review. New Mexico Department of Game and Fish, Santa Fe
- Pfeifer M & al. (2014) BIOFRAG — a new database for analyzing BIOdiversity responses to forest FRAGmentation. Ecol. Evol. 4, 1524–1537
- Le biote est l'ensemble des organismes vivants d'un lieu ou secteur donné, qui avec le biotope constituent le milieu biotiqueGlossaire Aquaportail
- (en) S. Sahney, M.J. Benton et H.J. Falcon-Lang, « Rainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica », Geology, vol. 38, no 12, , p. 1079–1082 (DOI 10.1130/G31182.1, lire en ligne [PDF])
- Les organismes saproxylophages sont ceux qui se nourrissent de bois mort en contribuant à sa décomposition (champignons, bactéries, invertébrés dont de nombreux insectes)
- Harris L.D. 1984 - "The fragmented forest. Island biogeography theory and the preservation of biotic diversity". University of Chicago Press, Chicago, Illinois, États-Unis
- Noss R.F., Csuti B., 1997 - Habitat Fragmentation. In : Principles of Conservation Biology, 2d ed. Meffe G.K. & Carroll C.R. Sinauer Associates, Inc.
- Fahrig L., 2003 - Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 34:487-515
- Alan R. Templeton, Kerry Shaw, Eric Routman and Scott K. Davis (1990), The Genetic Consequences of Habitat Fragmentation ; Annals of the Missouri Botanical Garden Vol. 77, No. 1 (1990), p. 13-27 Published by: Missouri Botanical Garden Press Article Stable URL: https://www.jstor.org/stable/2399621
- Dale V.H., Pearson S.M., 1997 - Quantifying habitat fragmentation due to land-use change in Amazonia. In: W.F. Laurance & R.O. Bierregaard, eds. Tropical forest remnants: ecology, management, and conservation of fragmented communities. University of Chicago Press, Chicago, Illinois, USA
- Riitters K., Wickham J., O'Neill R., Jones B., Smith E., 2000 - Global-scale patterns of forest fragmentation. Conservation Ecology ; 4, 2, 3 [online] URL:
- Wuldera M.A., White J.C. , Andrew M.E. , Seitz N.E., Coops N.C., 2009 - Forest fragmentation, structure, and age characteristics as a legacy of forest management ; Forest Ecology and Management 258, 9, 10: 1938-1949 ; DOI:10.1016/j.foreco.2009.07.041
- Nepstad D.C., Lefebvre P., Davidson E.A., 1999 - Positive feedbacks in the fire dynamic of closed canopy tropical forests. Science 284:1832-1835
- Nepstad D.C., Verissimo A., Alencar A. , Nobre C., Lima E., Lefebvre P., Schlesinger P., Potter C., Moutinho P., Mendoza E., Cochrane M., Brooks V., 1999 -
- Alroy J (2017) Effects of habitat disturbance on tropical forest biodiversity. Proceedings of the National Academy of Sciences, 201611855.
- Leemans R. 1990 - Global data sets collected and compiled by the biosphere project. Working Paper, International Institute of Applied Systems Analysis, Laxenburg, Austria
- Gérard Blake, Les radeaux de végétation de la retenue de Petit-Saut (Guyane) ; Hydroécol. Appl.1997, 9, p. 195-211 DOI:10.1051/hydro:1997008 ; https://dx.doi.org/10.1051/hydro:1997008 (résumé)
- Lindenmayer D.B., Cunningham R.B., Tanton M.T., A.P. Smith, 1990 - The conservation of arboreal marsupials in the montane ash forests of the Central Highlands of Victoria, South-East Australia. II. The loss of trees with hollows and its implications for the conservation of Leadbeater's Possum Gymnobelideus leadbeateris McCoy (Marsupialia: Petauridae). Biological Conservation, 54, 133-145 (résumé) ; voir aussi base documentaire : Victorian Central Highlands Study Publications
- de Maynadier P.G. & Hunter M.L., 2000 - Road effects on amphibian movements in a forested landscape. Nat. Areas J. 20, 56–65
- Freid M.A., 1994 -The effects of ownership patterns on forest road netorks in western oregon ; Oregon State University Department of Geosciences ; Master of Science
- Lyon L.J., 1984 - Road effects and impacts on wildlife and fisheries. Forest Transportation Symposium, December 11-13, 1984, Casper, Wyoming. Proceedings publ. US Department of Agriculture, Forest Service, Rocky Mountain Region, Denver, CO
- Reed R.A., J.Johnson-Barnard, Baker W.L., 1996 - Contribution of roads to forest fragmentation in the Rocky Mountains. Conservation Biology 10:1098-1106 (résumé)
- Megahan W., 1987 - Effects of forest roads on watershed function inmountainous areas, p. 335-347. In Balasubramanivam et al, (eds.), Environmental Geotechniques and Problematic Soils. Balkema, Rotterdam, Pays-Bas
- Harr R., Harper J., 1975 - Changes in storm hydrographs after roadbuilding and clear cutting in the Oregon Coast Range. Water Resources Research, 11(3), p. 436-444
- Grant G., Crozier M., Swanson F., 1984 - An Approach ToEvaluating Off-site Effects of Timber Harvest Activities on Channel Morphology. Symposium on Effects of Forest Land Use on Erosion and Slope Stability. O'Laughlin C. & Pearce A. eds. May 7-11, 1984. Honolulu, HI]
- Jones J.A. (Oregon State University, Corvallis); Grant G.E. (Apr 1996), "Peak flow responses to clear-cutting and roads in small and large basins, western Cascades, Oregon", Water resources research ; (ISSN 0043-1397) ; Vol. n°.v. 32(4) p. 959-974 (résumé)
- Wemple, B.C. (Oregon State University, Corvallis, OR), Jones J.A., Grant G.E., "Channel network extension by logging roads in two basins, western Cascades, Oregon", Water resources bulletin ; (ISSN 0043-1370) ; v. 32(6) p. 1195-1207 (résumé)
- Charrier S., Petit S., Burel F., 1997 - Movements of Abax parallelepipedus (Coleoptera, Carabidae) in woody habitats of a hedgerow network landscape: a radio-tracing study. Original Research Article Agriculture, Ecosystems & Environment, 61, 2–3, Feb. 1997, 133-144 (résumé)
- Burel Françoise, Dynamique d'un paysage, réseaux et flux biologiques = Landscape dynamics: structural networks and biological fluxes (Thèse de nouveau doctorat, en Écologie, sous la direction de J.-C. Lefeuvre) ; Université de Rennes 1 ; 1991 [235 p. (bibl.: 410 ref.)résumé
- Normandin D. (INRA, Laboratoire d'Économie Forestière (ENGREF, AgroParisTech), Sciences Sociales, Agriculture et Alimentation, Espace et Environnement, Centre de recherche de Nancy ; Guide d'aide à la décision économique en matière d'équipement routier des forêts ; 1984, 43 p. ; fait suite à l'étude : Normandin D., 1978 - Économie comparée des transports forestiers. Rentabilité comparée des voies forestières. INRA, 173 p.
- Laurance SG, Stouffer P, Laurance WF (2004) Effects of road clearings on movement patterns of understory rainforest birds in Central Amazonia. Conserv Biol 18(4):1099–1109
- Laurance SG & Gomez MS (2005) Clearing width and movements of understory rainforest birds. Biotropica 37(1):149–152
- Riitters K., Wickham J., O'Neill R., Jones B., Smith E., 2000 - Global-scale patterns of forest fragmentation. Conservation Ecology 4(2): 3 )
- Skole D & Tucker C (1993) Tropical deforestation and habitat fragmentation in the Amazon : satellite data from 1978 to 1988. Science, 260 : 1905-1910.
- Claessens O, Granjon L, de Massary J.C & Ringuet S (2002) La station de recherches de Saint-Eugène: situation, environnement et présentation générale. Revue d'écologie.
- Forget P-M & Pons J-M (2002) Perspectives de recherches et futur de la Station de Saint-Eugène. Rev. Eco/. (Terre Vie), suppl.8 : 195-199
- Birnbaum P (1996) Nouveaux milieux, évolution floristique et structurale du massif forestier de Petit Saut 18 mois après la mise en eau du barrage, in : Grégoire A. (ed.), Actes des Sixièmes Journées « Environnement » relatives à la protection du site de Petit Saut (Guyane) - Sinnamary, 21 et 22 juin 1995. EDF.
- Claessens O (2000) Effets de la fragmentation de l'habitat sur les peuplements d'oiseaux forestiers tropicaux. Le cas de la mise en eau du barrage de Petit Saut, Guyane française ; Thèse de doctorat, Muséum national d'Histoire naturelle, Paris. 348 p.
- De Massary J-C (2001) Effets de la fragmentation de l'habitat sur les peuplements et les populations de lézards terrestres en forêt tropicale : l 'exemple du barrage de Petit Saut en Guyane française. Thèse de doctorat, Muséum national d'Histoire naturelle, Paris, 171 p
- Horeau V, Cerdan P & Champeau A (1997) La mise en eau du barrage hydroélectrique de Petit Saut (Guyane) : ses conséquences sur les peuplements d'invertébrés aquatiques et sur la nourriture des poissons. Hydroécol. Appl., 9 : 213-240.
- Kapos V (1989) Effects of isolation on the water status of forest patches in the Brasilian Amazon. J. Trop. Eco/., 5 : 173-185.
- Riguet S. (1998) Evolution des peuplements et des populations de micromammijères terrestres de forêt tropicale à la suite de la fragmentation de leur habitat : l'exemple du barrage de Petit Saut (Guyane française). Thèse de doctorat, Muséum national d'Histoire naturelle, Paris, 194 p.
- Turner I.M & Corlett R.T (1996) The conservation value of small, isolated fragments of lowland tropical rain forest Trends Eco/. Evol., 1 1 : 330-333
- Vié J.C. (1998) Les effets d'une perturbation majeure de l'habitat sur deux espèces de primates en Guyane française : translocation de singes hurleurs roux (Alouatta seniculus) et translocation et insularisation de sakis à face pâle (Pithecia pithecia). Thèse de doctorat, Université de Montpellier II, 204 p.
- Villiers J.F (1998) Structure et flore de la forêt dense fragmentée à Saint-Eugène. Echos du Courcibo, 6 : 7-8
- Dale V.H., Pearson S.M., 1997 - Quantifying habitat fragmentation due to land-use change in Amazonia. in W.F. Laurance, R.O. Bierregaard, eds. Tropical forest remnants: ecology, management, and conservation of fragmented communities. University of Chicago Press, Chicago, Illinois, États-Unis
- Riitters K., Wickham J.D., Jones K.B., 2003 - Distribution and Causes of Global Forest Fragmentation Résumé
- K.H. Riitters, J.W. Coulston ; Environmental assessment; Hot Spots of Perforated Forest in the Eastern United States ; Environmental Management ; 35, 4 : 483-492, DOI: 10.1007/s00267-003-0220-1 (Résumé)
- Estreguil, C., Caudullo, G., de Rigo, D., San-Miguel-Ayanz, J., (2013), Forest landscape in Europe: Pattern, fragmentation and connectivity. Reference Report by the Joint Research Centre of the European Commission. EUR – Scientific and Technical Research 25717 (JRC 77295). (ISSN 1831-9424). (ISBN 978-92-79-28118-1). doi:10.2788/77842. Luxembourg: Publications Office of the European Union (Reproduction is authorised provided the source is acknowledged)
- JRC European-wide forest pattern web map viewer, voir http://forest.jrc.ec.europa.eu/efdac/ European Forest Data Centre] (EFDAC) et son Catalogue de métadonnées
- Lindenmayer, D., Hobbs R.J., Montague-Drake , R., Alexandra, J., Bennett, A., Burgman, M., Cale, P., Calhoun, A., Cramer, V., Cullen, P., Driscoll, D., Fahrig, L., Fischer, J., Franklin, J., Haila, Y., Hunter, M., Gibbons, P., Lake, S., Luck, G., MacGregor, C., McIntyre, S., MacNally, R., Manning, A., Miller, J., Mooney, H., Noss, R., Possingham, H., Saunders, D., Schmiegelow, F., Scott, M., Simberloff, D., Sisk, T., Tabor, G., Walker, B., Wiens, J., Woinarski, J., Zavaleta, E., (2008), A checklist for ecological management of landscapes for conservation. Ecology Letters 11(1), 78-91. doi: 10.1111/j.1461- 0248.2007.01114.x
- Kettunen, M, Terry, A., Tucker, G., Jones A. (2007), Guidance on the maintenance of landscape features of major importance fo r wild flora and fauna - Guidance on the implementation of Article 3 of the Birds Directive (79/409/EEC) and Article 10 of the Habitats Directive (92/43/EEC). Institute for European Environmental Policy (IEEP), Brussels, 114 pp. and Annexes. Report on the review of measures to promote connectivity. EC Project ‘Guideli nes: Adaptation, Fragmentation’. ENV.B.2/ETU/2006/0042r
- J. Seppala et al., Greenhouse gas emissions from the use of primary energy in forest operations and long-distance transportation of timber in Finland ; Resources, Conservation and Recycling 23 (1998) 87–105, Finnish Environment Institute, Helsinki, Finland voir page 101
- Satu Löfman & Jari Kouki, Fifty Years of Landscape Transformation in Managed Forests of Southern Finland. Scandinavian Journal of Forest Research 16, 1, 2001 ; 44-53 ; DOI:10.1080/028275801300004406 (Résumé)
- Saura, S., Estreguil, C., Mout on, C., Rodríguez-Freire, M. (2011), Ecological Indicators - Network analysis to assess landscape connectivity trends: Application to European forests (1990–2000). Ecological Indicators 11(2) 407–416. doi: 10.1016/j.ecolind.2010.06.011 (résumé)
- Source : données IGN BDTopo 2011, IFN, INSEE, Corine Land Cover 2006 (information géographique pré-traitée par J. Bonneau 2011 pour NatureParif ; Fragmentation forestière par les infrastructures de transport
- IRSTEA/EFNO, Interactions gestion forestière et biodiversité spécifique, consulté 2013-01-10
- Point sur le bilan patrimonial des forêts (Dossier no 3, ONF, mai 2006, page 2/4)
- Rapport du 27 avril 1999 "L'avenir de la Forêt de Fontainebleau" de Jean Dorst, Geneviève Humbert et Dominique Bourg ; version pdf
- Lamiche F., 2011 - http://scd.uhp-nancy.fr/docnum/SCDSCI_M_2011_LAMICHE_FLAVIEN.pdf Importance de la discontinuité spatiale dans le cas de la restauration de la connectivité de la trame forestière en Lorraine], Inra/Université de Nancy, UMR Écologie et Ecophysiologie forestières ; Mémoire de stage, Nancy le 06/09/2011, PDF, 38 p. (source : Résumé (page 38), voir aussi Conclusion p. 27)
- Piveteau V., Arnould P., Barthod C., Bras P., Hotyat M., de Montgolfier J., Pointereau P., 1999 - Les espaces boisés en France : bilan environnemental, IFN (Institut français de l'environnement, Orléans) ; Les Dossiers de l'environnement ; (ISSN 1272-1026), 197 p. ; (ISBN 2-911089-15-4 et 2-87671-289-X) (Fiche INIST/CNRS)
- L'Indice de compacité d'un ensemble forestier est le rapport du périmètre d'un cercle (= compacité max.) ayant la même surface que la moyenne de l'ensemble concerné, au périmètre moyen
- Baur A. & Baur B., 1990 -'Are roads barriers to dispersal in the land snail Arianta arbustorum ? Can J. Zool. 68, 613-617. (Résumé)
- Edenhamn, p. 1996 Spatial dynamics of the European tree frog (Hyla arborea L.) in a heterogenous landscape. PhD thesis, Uppsala University, Uppsala, Sweden
- A Rio en juin 1992, l'ONU. L'ensemble des États-membres a identifié deux enjeux environnementaux majeurs pour l'humanité : l'érosion de la biodiversité, et le réchauffement climatique
- Bird M., 1995 - Fire, prehistoric humanity, and the environment. Research School of Earth Sciences, The Australian National University, Canberra, Australia, Interdisciplinary Science Reviews, 20, 2, June 1995, p. 141-154(14) ; d:Maney Publishing
- John F. Boyle, Marie-José Gaillard, Jed O. Kaplan, John A. Dearing (2011), Modelling prehistoric land use and carbon budgets: A critical review ; The Holocene 21: 715-722, 1re publication: 17, 2011-02-17 (Résumé)
- F.S. Anselmetti, D.A. Hodell, D. Ariztegui, M. Brenner, M.F. Rosenmeier, "Quantification of soil erosion rates related to ancient Maya deforestation" ; The Geological Society of America, Inc (GSA). ; doi: 10.1130/G23834A.1 v. 35 no. 10 p. 915-918 (Résumé)
- J.O. Kaplana, K.M. Krumhardta, N. Zimmermann, "The prehistoric and preindustrial deforestation of Europe" ; Quaternary Science Reviews28, 27–28, December 2009, 3016–3034 (Résumé)
- La zoochorie est le transport de grains de pollen et spores, graines ou propagules par des animaux, qui permet la dispersion et l'entretien de la diversité génétique de nombreuses espèces de plantes (dont arbres) et champignons
- Henschel C., 2003 - The Road Less Travelled? A Report on the Effectiveness of Controlling Motorized Access in Remote Areas of Ontario, A Case Study in Temagami, Global Forest Watch Canada Report, 42 p.
- John A. Kupfer ; National assessments of forest fragmentation in the USA, Global Environmental Change ; Volume 16, Issue 1, February 2006, Pages 73-82 ; doi:10.1016/j.gloenvcha.2005.10.003 (Résumé)
- The H. John Heinz III Center for Science, Economics, and the Environment. 2005, Forest pattern and fragmentation. In: The state of the nation’s ecosystems: Measuring the lands, waters, and living resources of the United States. New York, NY: Cambridge University Press. Web update 2005.
- Riitters, Kurt H.; Wickham, James; Coulston, John 2004. Use of Road Maps in National Assessments of Forest Fragmentation in the United States. Ecology and Society 9(2): 13. (Résumé)
- EPA, Forest Fragmentation (explications plus détaillées, en anglais)
- Riitters, K.H. 2003. Report of the United States on the criteria and indicators for the sustainable management of temperate and boreal forests, criterion 1: Conservation of biological diversity, indicator 5: Fragmentation of forest types. Final report. FS-766A. In: Darr, D., ed. Data report: a supplement to the National Report on Sustainable Forests. Washington, DC: USDA Forest Service
- ECOFOR (2012): présentation du programme « AM-Tools » sur les outils écologiques et légaux pour la « migration assistée des forêts » ; AGROBIOSPHERE (Appel à projets 2011), coordonné par CNRS/AgroParisTech - LESE Laboratoire Écologie, consulté 2013-08-24
- Van Dongen S., Backeljau T., Matthysen E., Dhondt A.A., 1994 - Effects of forest fragmentation on the population structure of the winter moth Operophtera brumata L. (Lepidoptera, Geometridae). Acta oecologica, 15, 2, p. 193-206 (1 p. 1/2), (ISSN 1146-609X) (Résumé Inist/CNRS)
- Hingrat Y.et al., 2002 - Effets de la fragmentation forestière sur l'activité des coléoptères coprophages : Dispersion secondaire des graines en Guyane Française. Revue d'écologie, suppl., 2002 - (Fiche Inist CNRS)
- Forman R.T.T., 2000 - Estimate of the Area Affected Ecologically by the Road System in the United States, Conservation Biology, 14, 1, p. 31-35
- Mathieu D., 2004 - Études sur la valeur économique de l’écotourisme et du tourisme d’aventure, Rapport DBSF, Aventure Écotourisme Québec
- Tourisme Québec, 2003 - Écotourisme et tourisme de nature, orientations et plan d’action 2003-2008, Direction du développement des produits touristiques, Gouvernement du Québec, 73 p.,
- Kneeshaw D., Gauthier S., 2006 - Dossier Accessibilité forestière accrue : panacée ou boîte de Pandore ? ; Téoros, revue de recherche en tourisme ; 25-3 : Forêt, p. 36-40
- Lequin M., Carrière J.-B., 2003 - Planification de l’accessibilité à l’expérience globale en écotourisme. Téoros, 22, 3, p. 29-36
- Bourgeois L., Kneeshaw D.D., Boisseau G., 2006 - « routes forestières au Québec : Les impacts environnementaux, sociaux et économiques », Vertigo, 6, 2, septembre 2006
- du Bus de Warnaffe G., Lebrun P., 2002 Effects of forest management on carabid beetles in Belgium: implications for biodiversity conservation Biological Conservation, 118, 219-234
- IPCC, L’utilisation des terres, le changement d’affectation des terres et la foresterie ; Résumé à l’intention des décideurs ; Rapport spécial du Groupe d’experts intergouvernemental sur l’évolution du climat ; Publié pour le Groupe d’experts intergouvernemental sur l’évolution du climat ; (ISBN 92-9169-214-X)
- Foltz R., H. Rhee, K. Yanosek, 2007 - Infiltration, Erosion, and Vegetation Recovery Following Road Obliteration. Transactions of the American Society of Agricultural and Biological Engineers 50: 1937-1945
- Kervliet J., Hicks J., Wilmer B., Carbon sequestrered when uneeded national forest roads are revegetated (Brief-carbon-and-roads.pdf), the Wilderness Society, février 2010
- Weaver, W. and D. Hagans. (2004) Road Upgrading, Decommissioning, and Maintenance—Estimated Costs on Small and Large Scales. Salmon Habitat Restoration Cost Workshop. Gladstone, OR
- Inhara D., S. Hackett, J. Manning. (2003) Reinvestment in Jobs, Communities, and Forests: The Benefits and Costs of a National Program for Road Removal on U.S. Forest Service Lands, a Preliminary Analysis. The Center for Environmental Economic Development, Arcata, CA. 75 p.
- Kerkvliet J., Powell J., 2007 -The Ecologic and Economic Impacts of Decommissioning Unwanted Region 1 Forest Service Roads. Working Paper. The Wilderness Society. Bozeman, Montana
- Swatalski T., J. Bissonette, T. DeLuca, C. Luce, Madie M., 2004 - Benefits and Impacts of Road Removal. Frontiers in Ecology and the Environment 2: 21-28.
- Taxpayers for Common Sense. (2004). Road Wrecked: Why the $10 Billion Forest Service Road Maintenance Backlog Is Bad for Taxpayers
- voir figure 7 adapté de Cochrane & Laurance, 2002, in W.F. Laurance ; Review ; Theory meets reality: How habitat fragmentation research has transcended island biogeographic theory. Biological Conservation 141, 7, July 2008,1731–1744 ([Résumé])
- Laurance, W. F., S. G. Laurance, P. Delamonica, 1998 - Tropical forest fragmentation and greenhouse gas emissions. Forest Ecology and Management 110:173-180 (Résumé
- FAO, B.A. Wilcox & B. Ellis, Forests and emerging infectious diseases of humans With the interweaving of forests, pathogens and the development of human civilization, deforestation and other land use changes have an important part in the emergence of disease. ; (« Forest fragmentation affects disease dynamics by influencing host and vector abundance and distribution and thus the abundance or dispersal of pathogens »
- J.M. Morvan, E. Nakouné, V. Deubel & M. Colyn, Écosystèmes forestiers et virus Ebola, Man.no 2155/RIP9. 3e colloque du réseau international des Instituts Pasteur et instituts associés. 14-15 octobre 1999, Institut Pasteur de Paris
- Donahue JG, Piesman J, Spielman A. (January 1987). "Reservoir competence of white-footed mice for Lyme disease spirochetes. Am. J. Trop. Med. Hyg. 36 (1): 92–6.
- Secrétariat de la Convention sur la diversité biologique, L’Initiative LifeWeb (plaquette couleur recto-verso)
- Hansen M.C et al. (2013) High-resolution global maps of 21st-century forest cover change. Science 342, 850–853
- Tracewski, Ł. et al. (2016) Toward quantification of the impact of 21st-century deforestation on the extinction risk of terrestrial vertebrates. Conserv. Biol. 30, 1070–1079
- Barlow J et al (2016). [Barlow, J., Lennox, G. D., Ferreira, J., Berenguer, E., Lees, A. C., Mac Nally, R., ... & Parry, L. (2016). Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature, 535(7610), 144-147 Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation]. Nature 535, 144–147
- Matthew G. Betts, Wolf C, William J. Ripple, Ben Phalan, Kimberley A. Millers, Adam Duarte, Stuart H. M. Butchart& Taal Levi (2017), Global forest loss disproportionately erodes biodiversity in intact landscapes ; Nature |Doi:10.1038/nature23285 |résumé
- Kareiva, P., Watts, S., McDonald, R. & Boucher, T. (2007) Domesticated nature: shaping landscapes and ecosystems for human welfare. Science 316, 1866–1869
- Peres C.A Why we need megareserves in Amazonia. Conserv. Biol. 19, 728–733
- Buckley R., 1982 - The habitat model of island biogeography. Journal of Biogeography 9, 334–339
- Simberloff D.S., 1976 - Experimental zoogeography of islands: effects of island size. Ecology 57, 629–648
- Fahrig L., Merriam G., 1985 - Habitat patch connectivity and population survival. Ecology 66:1762–1768
- Grasman J., Hille Ris Lambers R., 1997- On local extinction in a metapopulation. Ecological Modelling 103: 71–80
- Hanski I., 1997 - Metapopulation dynamics: from concepts and observations to predictive models. In: Hanski I., Gilpin M.E., eds. Metapopulation biology: ecology, genetics and evolution. London, Royaume-Uni: Academic Press, 69–92
- Kirchner F., Ferdy J.B., Andalo C., Colas B., Moret J., 2003 - Role of corridors in plant dispersal: an example with the endangered Ranunculus nodiflorus. Conservation Biology 14: 401–410
- Raoul F., Quere J., Pleydell D., Vaniscotte A., Giraudoux P., 2008 - Small mammals assemblage response to deforestation and aforestation in Central China : a multinomial based modelling approach. Mammalia 72
- Ferraz G., Russell G.J., Stouffer P.C., Bierregaard Jr. R.O., Pimm S.L., Lovejoy T.E., 2003 - Rates of species loss from Amazonian forest fragments. Proceedings of the National Academy of Sciences 100: 14069-14073
- Anak Pattanavibool, Dearden P. (University of Victoria), 2002 - Fragmentation and wildlife in montane evergreen forests, Northern Thailand. Biological Conservation 107, 155–164
- Laurance W.F., Ferreira L.V., Merona J.M. R., Laurance S.G., Hutchings R.W. & Lovejoy T.E., 1998 - Effects of forest fragmentation on recruitment patterns in Amazonian tree communities. Conservation Biology 12: 460-464
- Jacquemyn H., Butay J. & Hermy M., 2001 - Forest plant species richness in small, fragmented mixed deciduous forest patches: the role of area, time and dispersal limitation. Journal of Biogeography 28: 801-812
- Laurance W.F., Delamônica P., Laurance S.G., Vasconcelos H.L. & Lovejoy T.E., 2000 - Rainforest fragmentation kills big trees. Nature 404: 836
- Bender, D. J., Contreras, T. A. & Fahrig, L., 1998 - Habitat loss and population decline: a meta-analysis of the patch size effect. Ecology 79: 517-533
- Bierregaard Jr. R.O., Lovejoy T.E., Kapos V., dos Santos A.A. & Hutchings R. W., 1992 - The biological Dynamics of tropical rainforest fragments: A prospective comparison of fragments and continuous forest. BioScience 42: 859-866
- Price O.F., Woinarski J.C.Z., Robinson D., 1999 - Very large requirements for frugivorous birds in monsoon rainforests of the Northern Territory, Australia. Biological Conservation 91, 169–180
- Lynam A.J., Billick I., 1999 - Differential responses of small mammals to fragmentation in a Thailand tropical forest. Biological Conservation 91, 191–200
- Kupfer J.A., Scott B. Franklin, 2009 - Linking Spatial Pattern and Ecological Responses in Human-Modified Landscapes: The Effects of Deforestation and Forest Fragmentation on Biodiversity. Geography Compass 3:4, 1331-1355
- Gascon C., Lovejoy T.E., Bierregaard Jr. R.O., Malcolm J.R., Stouffer P.C., Vasconcelos H.L., Laurance W.F., Zimmerman B., Tocher M., Borges S., 1999 - Matrix habitat and species richness in tropical forest remnants. Biological Conservation 91, 223–229
- Umapathy G., Kumar A., 2000 - The occurrence of arboreal mammals in the rain forest fragments in the Anamalai Hills, south India. Biological Conservation 92, 311–319
- Tewksbury, J.J., Hejl, S.J. & Martin, T.E., 1998. Breeding productivity does not decline with increasing fragmentation in a western landscape. Ecology 79(8): 2890-2903
- W. Andrew Cox, Frank R. Thompson, John Faaborg. (2012) Landscape forest cover and edge effects on songbird nest predation vary by nest predator. Landscape Ecology Online publication date: 4-Feb-2012 (Résumé)
- Tewksbury, Joshua J., Sallie J. Hejl, and Thomas E. Martin. (1998), Breeding productivity does not decline with increasing fragmentation in a western landscape ; Ecology 79:2890–2903. (résumé)
- Beier, P., Van Drielen, M. & Kankam, B. O., 2002. Avifaunal collapse in west african forest fragments. Conservation Biology 16: 1097-1111
- S. Pütz, J. Groeneveld, L.F. Alves, J.P. Metzger, A. Huth, Fragmentation drives tropical forest fragments to early successional states: A modelling study for Brazilian Atlantic forests ; Ecological Modelling, Volume 222, Issue 12, 24 June 2011, Pages 1986-1997 (Résumé)
- Guadalupe Williams-Linera, Tree species richness complementarity, disturbance and fragmentation in a Mexican tropical montane cloud forest ; Biodiversity and Conservation Volume 11, Number 10 (2002), 1825-1843, DOI: 10.1023/A:1020346519085 (Résumé)
- Cullen, L., Jr. (1997), Hunting and biodiversity in Atlantic forest fragments, Sao paulo, Brazil. MS thesis, Université de Floride, Gainesville
- Cullen L., Jr. E. Bodmer, C.V. Padua, 2000 -Effects of hunting in habitats fragments ot the Atlantic forests, Brazil, Biological Conservation 95; 49-56, 16 p.)
- Ayres J.M., D.M. Lima, E.D.S. Martins, J.L.K. Barreiros, 1991 - On the track of the road : changes in subsistence hunting in a brazilian amazonian village. p. 82-92, in J.G. Robinson, K.H. Redfort, editors, Neotropical wildlife use and conservation. Université de Chicago
- Eric Motard, Audrey Muratet, Danielle Clair-Maczulajtys, Nathalie Machon, Les populations invasives d’Ailanthus altissima menacent-elles la diversité floristique des forêts tempérées périurbaines ? Comptes Rendus Biologies, 334, 12, December 2011 : 872-879 (résumé)
- (en) Martin Hermy, Olivier Honnay, Les Firbank, Carla Grashof-Bokdam et Jonas E. Lawesson, « An ecological comparison between ancient and other forest plant species of Europe, and the implications for forest conservation. », Biological Conservation, vol. 91, no 1, , p. 9–22 (résumé)
- Buée M., Maurice J.-P., Zeller B., Andrianarisoa S., Ranger J., Courtecuisse R., Marçais B., Le Tacon F., 2011 - Influence of tree species on richness and diversity of epigeous fungal communities in a French temperate forest stand. Fungal Ecology 4: 22-31
- Diédhiou A.G., Dupouey J.-L., Buée M., Dambrine E., Laüt L., Garbaye J., 2009 - Response of ectomycorrhizal communities to past Roman occupation in an oak forest. Soil Biol. Biochem. 41: 2206-2213 (résumé)
- A.G. Diedhiou, J.-L. Dupouey, M. Buée, É. Dambrine, L. Laüt, J. Garbaye, The functional structure of ectomycorrhizal communities in an oak forest in central France witnesses ancient Gallo-Roman farming practices. Soil Biology and Biochemistry, 42, 5, May 2010 : 860-862 (résumé)
- Esseen PA & Renhorn KE (1998) Edge Effects on an Epiphytic Lichen in Fragmented Forests Conservation Biology Vol 12, no 6, décembre 1998, p. 1307–1317, publié en ligne 2008-07-07, DOI:10.1111/j.1523-1739.1998.97346.x
- Felipe Pimentel Lopes de Melo, Rodolfo Dirzo, Marcelo Tabarelli, “Biased seed rain in forest edges: Evidence from the Brazilian Atlantic forest” ; Biological Conservation ; Volume 132, Issue 1, September 2006, Pages 50–60 (Résumé)
- Janaina B.P. Costa, Felipe P.L. Melo, Bráulio A. Santos, Marcelo Tabarelli, 2012 -Reduced availability of large seeds constrains Atlantic forest regeneration. Acta Oecologica, 39, 61-66 (Résumé)
- Janaina B.P. Costa, Felipe P.L. Melo, Bráulio A. Santos Marcelo Tabarelli, 2012 - Reduced availability of large seeds constrains Atlantic forest regeneration. Acta Oecologica 39, 61–66 (Résumé)
- William F. Laurance, Susan G. Laurance, Leandro V. Ferreira, Judy M. Rankin-de Merona, Claude Gascon et Thomas E. Lovejoy, Rapport : "Biomass Collapse in Amazonian Forest Fragments" ; Science 7 November 1997: Vol. 278 no. 5340 p. 1117-1118 ; DOI:10.1126/science.278.5340.1117 (Résumé)
- Laurance, W. F., Ferreira, L. V., Gascon, C. & Thomas, E., 1997a. Biomass decline in Amazonian forest fragments. Science 278: 1117-1118
- David Cowles, "Biomass Decline in Amazonian Forest Fragments" ; Science 27 November 1998:Vol. 282 no. 5394 p. 1611 ; DOI: 10.1126/science.282.5394.1611a
- Izaya Numata, Mark A. Cochrane, Dar A. Roberts, João V. Soares, Carlos M. Souza, Marcio H. Sales. (2010) Biomass collapse and carbon emissions from forest fragmentation in the Brazilian Amazon. Journal of Geophysical Research 115:G3, En ligne : 1-Jan-2010
- Izaya Numata, Mark A Cochrane, Carlos M Souza Jr, Marcio H Sales. (2011) Carbon emissions from deforestation and forest fragmentation in the Brazilian Amazon. Environmental Research Letters 6:4, 044003
- M.A. Oliveira, A.M.M. Santos, M. Tabarelli, “Profound impoverishment of the large-treestand in a hyper-fragmented landscape of the Atlantic forest” ; Forest Ecology and Management, 256, 11, 20 November 2008 : 1910–1917 (Résumé)
- José Julio de Toledo, William E. Magnusson, Carolina V. Castilho, Henrique E.M. Nascimento. (2012) Tree mode of death in Central Amazonia : Effects of soil and topography on tree mortality associated with storm disturbances. Forest Ecology and Management 263, 253-261
- Zachos F.E., Althoff C., Steynitz Y.V., Eckert I., Hartl G.B. [2007]. Genetic analysis of an isolated red deer (Cervus elaphus) population showing signs of inbreeding depression. European Journal of Wildlife Research, 53 : 61-67 (7 p., 2 fig., 2 tab., 61 réf.). Pour lire l’article : European Journal of Wildlife Research (www.springerlink.com/content/1439-0574/
- William F. Laurance, Diana C. Useche, 2009 - Environmental Synergisms and Extinctions of Tropical Species. Conservation Biology 23:6, 1427-1437
- Izaya Numata, Mark A. Cochrane, Dar A. Roberts, João V. Soares, 2009 - Determining dynamics of spatial and temporal structures of forest edges in South Western Amazonia. Forest Ecology and Management 258:11, 2547-2555 ; En ligne: 10-Nov-2009
- Laurance W. F., Lovejoy T. E., Vasconcelos H. L., Bruna E. M., Didham R. K., Stouffer P. C., Gascon C., Bierregaard Jr. R. O., Laurance S. G. & Sampaio E., 2002 - Ecosystem decay of Amazonian forest fragments : a 22-year investigation. Conservation Biology 16: 605-618
- P.F. Falcão, S.R.R. Pinto, R. Wirth, I.R. Leal., 2010 - Edge-induced narrowing of dietary diversity in leaf-cutting ants. Bulletin of Entomological Research1-7
- Bráulio A. Santosa, Carlos A. Peresb, Marcondes A. Oliveirac, Alexandre Grilloc, Cecília P. Alves-Costac, Marcelo Tabarellic, Drastic erosion in functional attributes of tree assemblages in Atlantic forest fragments of northeastern Brazil. Biological Conservation 141, 1, January 2008 : 249–260 (Résumé)
- M. Oliveira, A. Santos, M. Tabarelli, 2008 - Profound impoverishment of the large-tree stand in a hyper-fragmented landscape of the Atlantic forest. Forest Ecology and Management 256:11, 1910-1917 (résumé)
- Laurance W.F., Bierregaard R.O. (Eds.), 1997 -Tropical Forest Remnants : Ecology, Management, and Conservation of Fragmented Communities. University of Chicago Press, Chicago
- Guindon C.F., 1996 -The importance of forest fragments to the maintenance of regional biodiversity in Costa Rica. In: Schelhas J., Greenberg R. (Eds.), Forest Patches in Tropical Landscapes. Island Press, Washington, DC, p. 168–186
- Corlett R.T., Turner I.M., 1997 - Long-term survival in tropical forest remnants in Singapore and Hong Kong. In: Laurance W.F., Bierregaard Jr. R.O. (Eds.), Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities. University of Chicago Press, Chicago, p. 333–345
- Cornelius C., Cofre H., Marquet P.A., 2000 - Effects of habitat fragmentation in a relict temperate forest in semiarid Chile. Conservation Biology 14, 534–543
- Lovejoy T.E., Bierregaard Jr. R.O., Rylands A.B., Malcolm J.R., Quintela C.E., Harper L.H., Brown Jr. K.S., Powell A.H., Powell G.V.N., Schubart H.O.R., Hays M.B., 1986 - Edges and other effects of isolation on Amazon forest fragments. In: Soule M.E. (Ed.), Conservation Biology: The Science of Scarcity and Diversity. Sinauer Associates, Sunderland, Massachusetts, p. 257–285
- Bierregaard Jr. R.O., Lovejoy T.E., Kapos V., dos Santos A.A., Hutchings R.W., 1992 - The Biological Dynamics of Tropical Rainforest Fragments.BioScience, 42, 11, Stability and Change in the Tropics (Dec., 1992), p. 859-866 (Résumé)
- Pither R., Kellman M., 2002 -Tree species diversity in small, tropical riparian forest fragments in Belize, Central America. Biodiversity and Conservation 11, 9: 1623-1636, DOI: 10.1023/A:1016831916994 (Résumé)
- Jacquemyn H., Butaye J. & Hermy M. (2003), Influence of environmental and spatial variables on regional distribution of forest plant species in a fragmented and changing landscape ; Ecography 26: 768-776
- Moisès Guardiola, Joan Pino, Ferran Rodà. (2013) Patch history and spatial scale modulate local plant extinction and extinction debt in habitat patches. Diversity and Distributionsn/a-n/a
- Kellman M., 1996. Redefining roles: plant community reorganization and species preservation in fragmented systems. Global Ecology and Biogeography Letters 5: 111-116
- M.G. Betts, G.J. Forbes, A.W. Diamond, P.D. Taylor, 2006 - Independent Effects of Fragmentation on Forest Songbirds: An Organism-Based Approach. Ecological Applications 16:3, 1076-1089 Online publication date: 1-Jun-2006 résumé
- Lindborg, Regina, et Ove Eriksson (2004), Historical landscape connectivity affects present plant sepcies diversity ; Ecology 85:1840–1845. https://dx.doi.org/10.1890/04-0367 (résumé)
- D. Lauterbach, C. Römermann, F. Jeltsch, M. Ristow. (2013) Factors driving plant rarity in dry grasslands on different spatial scales: a functional trait approach. Biodiversity and Conservation
- Pfeifer, M. et al. BIOFRAG — a new database for analyzing BIOdiversity responses to forest FRAGmentation. Ecol. Evol. 4, 1524–1537 (2014)
- Hudson, L. N., Newbold, T., Contu, S., Hill, S. L., Lysenko, I., De Palma, A., ... & Choimes, A. (2014). The PREDICTS database: a global database of how local terrestrial biodiversity responds to human impacts. Ecology and evolution, 4(24), 4701-4735.
- Pfeifer, M., Lefebvre, V., Peres, C. A., Banks-Leite, C., Wearn, O. R., Marsh, C. J., ... & Cisneros, L. (2017). Creation of forest edges has a global impact on forest vertebrates. Nature|résumé.
- Haddad, N. M. et al. (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 1, e1500052
- Trombulak S.C., Frissell C.A., 2000 - Review of ecological effects of roads on terrestrial and aquatic communities. Conservation Biology 14:18-30
- Spellerberg I.F., 1998 - Ecological effects of roads and traffic : a literature review. Global Ecology and Biogeograhpy Letters 7:317-333
- Patrizia Digiovinazzo, Gentile Francesco Ficetola, Luciana Bottoni, Carlo Andreis, Emilio Padoa-Schioppa, 2010 - Ecological thresholds in herb communities for the management of suburban fragmented forests."Forest Ecology and Management 259, 3, 343–349 Résumé
 Portail de l’écologie
Portail de l’écologie  Portail de la conservation de la nature
Portail de la conservation de la nature  Portail du bois et de la forêt
Portail du bois et de la forêt