Fruit (botanique)
Le fruit, en botanique, est l'organe végétal contenant une ou plusieurs graines. Caractéristique des Angiospermes, il succède à la fleur par transformation du pistil. La paroi de l'ovaire forme le péricarpe du fruit et l'ovule donne la graine[1].

Fonctions du fruit

Le fruit favorise la reproduction de l'espèce, en protégeant la ou les graines et en favorisant leur dissémination.
La protection des graines en formation se fait contre les animaux (le jeune fruit n'attire pas les animaux) et les contraintes climatiques.
Le fruit mûr favorise ensuite la dissémination des graines. Cette dissémination se fera d'une manière spécifique au type de fruit :
- par le biais des animaux pour les fruits en général colorés, sucrés (mûres) ou riche en nutriments (châtaigne) = endozoochorie
- par le vent pour les fruits disposant de parachute (pappus ou akène de pissenlit) ou d'une aile (samare d'érable) = anémochorie
- par l'eau pour les fruits flotteurs (noix de coco).
Formation du fruit
Le fruit se forme à partir de la fleur qui a été pollinisée. Il peut se présenter sous différentes formes : drupe, baie, gousse, capsule, akène, etc.
La formation du fruit résulte de la transformation du pistil après la fécondation, ou parfois sans fécondation (on parle dans ce cas de parthénocarpie). C'est plus précisément la paroi de l'ovaire (partie du pistil qui renferme l'ovule) qui devient la paroi du fruit, appelée péricarpe, entourant les graines. L'épiderme externe de cette paroi devient l'épicarpe, le parenchyme devient le mésocarpe, et l'épiderme interne, l'endocarpe. Selon les transformations de cette paroi, on obtient les différents types de fruits énumérés ci-après.







Faux-fruits
Dans certains cas, le « fruit » ne résulte pas de la transformation du pistil et peut avoir une origine plus complexe, c'est un faux-fruit. Sa formation peut résulter soit :
- de la transformation d'une autre partie de la fleur, le réceptacle floral. L'exemple le plus connu de faux-fruit est la pomme ou la fraise.
- de la transformation d'une autre partie de la fleur, les carpelles, devenant des drupéoles. Ces fruits multiples sont donc des petites drupes agglomérées, on parle alors de polydrupes. C'est le cas de la framboise et de la mûre (fruit de la ronce).
- de la transformation de plusieurs fleurs d'une inflorescence que l'on appelle une infrutescence. C'est par exemple le cas de l'ananas, de la figue ou de la mûre (fruit du mûrier). Ils font aussi partie des faux-fruits.
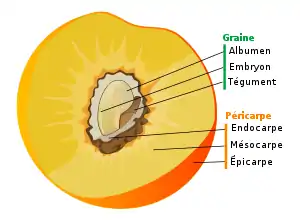
Structure du fruit
.png.webp)
Péricarpe
Le péricarpe[1] est la paroi du fruit issue de la transformation après la fécondation de la paroi de l’ovaire. Pour les faux-fruits, il résulte de la transformation du réceptacle floral.
Le péricarpe est formé de trois couches :
Épicarpe
L'épicarpe ou exocarpe est en botanique la paroi extérieure d'un fruit.
Il recouvre la couche appelée mésocarpe.
Il est généralement coloré.
Il est usuellement appelé peau ou écorce.
Mésocarpe
Le mésocarpe constitue la partie intermédiaire du fruit communément appelée pulpe quand il s'agit de fruits charnus.
Il dérive de la transformation du parenchyme de la paroi de l'ovaire.
Dans le cas particulier des agrumes, la partie externe du mésocarpe, blanche et spongieuse, se nomme albédo.
Endocarpe
L'endocarpe est le feuillet le plus interne du péricarpe, tissu du fruit entourant la graine. Il permet notamment de différencier une baie d'une drupe parmi les fruits charnus. S'il est sclérifié, il forme un noyau autour de la graine (le fruit sera une drupe), s'il ne l'est pas, la graine sera nommée pépin (le fruit sera une baie).
Ainsi, en botanique, on considérera par exemple que l'avocat contient un pépin et la pêche un noyau.
Typologie
On distingue en botanique :
- Les fruits charnus :
- Les fruits secs :
- fruits secs déhiscents (qui finissent par s'ouvrir) :
- follicule : hellébore, pivoine, etc ;
- gousse : fruit caractéristique des Fabacées, appelé aussi légumineuses : petit pois, soja, robinier, luzerne, etc ;
- capsule :
- capsule à déhiscence par fentes (septicide) : colchique, tabac, gentiane, etc.
- capsule loculicide : tulipe, lys, violette, etc.
- silique, à déhiscence paraplacentaire : fruit caractéristique des brassicacées : chou, colza, etc.
- pyxide, à déhiscence circulaire : mouron rouge, etc.
- capsule à déhiscence apicale : œillet, pavot, etc.
- fruits secs indéhiscents (qui ne s'ouvrent pas) :
- fruits secs déhiscents (qui finissent par s'ouvrir) :
Tendances évolutives
La conquête des terres émergées par les plantes chlorophylliennes puis par les premiers végétaux vasculaires à la limite Ordovicien-Silurien (445 Ma) est suivie au moins dès le Permien (300 Ma), par l'adoption d'une stratégie de dispersion différente, la zoochorie, qui favorise le transport des fruits charnus et des graines à arille (aliments riches en sucres et autres éléments nutritifs) consommées par des vertébrés herbivores[2]. Les groupes adaptés à la zoochorie se développent au Mésozoïque où les graines des fruits sont disséminées par les dinosaures herbivores et simultanément, par différents groupes de petits vertébrés (dont les reptiles)[2]. Au Crétacé qui marque la fin du Mésozoïque, les grands reptiles laissent la place aux oiseaux et aux mammifères frugivores qui ont pu établir des associations spécifiques vertébrés-plantes[2]. La majorité des Gymnospermes ont disparu au profit des plantes à fleurs et à fruits dont les graines ne sont pas nues mais protégées par des fruits qui sont encore de petite taille[2]. L'augmentation de taille des fruits résulte initialement d'un processus de coévolution avec les rongeurs et oiseaux frugivores qui participent à la dissémination de leurs graines[3]. Il est possible que l'évolution des fruits charnus corresponde au développement de traits de défense contre les prédateurs invertébrés plutôt que de traits attracteurs de vertébrés zoochores[4].
Actuellement, 64 % des familles de gymnospermes et 27 % des familles d'angiospermes produisent des fruits qui attirent les animaux favorisant la dispersion des graines[5],[6].
Voir aussi
Articles connexes
Lien externe
Notes et références
- Bernard Boullard, Plantes & Champignons, Éditions Estem, , 878 p.
- (en) Bruce H. Tiffney, « Vertebrate Dispersal of Seed Plants Through Time », Annual Review of Ecology, Evolution, and Systematics, vol. 35, , p. 1-29
- (en) David Dilcher, « Toward a new synthesis: Major evolutionary trends in the angiosperm fossil record », PNAS, vol. 97, no 13, , p. 7035.
- (en) Andrew L. Mack, « Did fleshy fruit pulp evolve as a defence against seed loss rather than as a dispersal mechanism? », Journal of Biosciences, vol. 25, , p. 93-97 (DOI 10.1007/BF02985186)
- (en) Carlos M. Herrera, Olle Pellmyr, Plant Animal Interactions: An Evolutionary Approach, John Wiley & Sons, (lire en ligne), p. 185.
- Thor Hanson, Le triomphe des graines, Buchet/Chastel, , p. 223
 Portail de l’agriculture et l’agronomie
Portail de l’agriculture et l’agronomie  Portail de la botanique
Portail de la botanique