Plan d'organisation
On appelle plan d'organisation d'un organisme la disposition relative de ses différents éléments constitutifs (axes de polarité, organes et organites). On doit cette notion à Georges Cuvier qui l'a appliquée à la zoologie, mais c'est Étienne Geoffroy Saint-Hilaire qui établit le lien avec l'évolution des espèces en introduisant la notion d'homologie.
Les plans d'organisation représentent un plan anatomique de base, partagé par tous les organismes d’un taxon, au niveau du phylum, de l’ordre, ou de la classe.
Un plan d'organisation est déterminé génétiquement puisque c'est un caractère héréditaire. Il est déterminé par des gènes du développement comme les gènes homéotiques chez la plupart des bilatériens.
Classiquement les plans d'organisation des animaux définissent les différents embranchements, mais la modernisation de cette notion a permis son extension à l'ensemble du monde vivant, à toutes les échelles (y compris unicellulaire) et à tous les niveaux hiérarchiques de la classification évolutionniste[1].
Les arbres se construisent aussi, en dendrologie, selon des plans d'organisation propres à leurs espèce et famille, et si une contrainte les éloigne de ce plan, ils forment des bois de tension ou des bois de compression pour s'en rapprocher[2].
Le concept des plans d’organisation fait partie intégrale d’autres concepts en biologie évolutive du développement, tels que l’homologie, la modularité et intégration, la canalisation, l’hétérochronie, la variabilité, ou l’évolutivité, d’où sa grande importance.
Historique
Le concept des «plans d’organisation» vient de l’anglais body plan, qui lui, provient de l’allemand bauplan qui se traduit par plan de construction.

On retrace les premiers concepts de plans d’organisation jusqu’à Aristote, qui décrit un plan structurel pour classifier les animaux, basé sur l’échelle de la nature. L’évolution du concept de plans d’organisation passera ensuite par Buffon, Cuvier, Lamarck, Geoffroy, von Baer, Owen, Darwin et finalement Haeckel, qui proposera la définition qui se rapporte le plus à celle qui est utilisée actuellement. Les travaux de von Baer furent ceux qui ont influencé le plus le concept actuel. Il mit au point 4 lois de développement (loi de Von Baer) :
- Durant le développement embryonnaire, les caractères généraux (ce que nous appelons maintenant les plans d’organisation) apparaissent avant les caractères particuliers.
- À partir des caractères les plus généraux (ou des plans d’organisation) se développent ceux qui sont moins généraux et finalement les caractères spécifiques.
- Ainsi, durant son développement, le plan d’organisation d’un animal s’écarte de plus en plus de celui des autres animaux.
- Les stades embryonnaires d’un animal ressemblent aux stades embryonnaires des animaux situés plus bas dans l’échelle du phylum, mais pas aux stades adultes.
Ces quatre lois étaient très proches de ce qui définit actuellement les plans d’organisation. C'est Darwin qui, un quart de siècle après von Baer, proposa que les caractéristiques communes des groupes d’animaux viennent, non pas des stades embryonnaires, mais bien d’un ancêtre commun.
Toutes les informations étaient maintenant en place pour obtenir la définition moderne des plans d’organisation, donnée par Woodger (en), en 1945 : les plans de corps représentent un plan anatomique de base, partagé par tous les organismes d’un taxon, au niveau du phylum, de l’ordre, ou de la classe. En biologie évolutive du développement, les plans de corps se concentrent sur l’origine et le maintien de caractéristiques développementales partagées par tous les membres d’un même phylum[3].
Des plans d'organisation à différents niveaux
Le plan d'organisation d'un taxon permet de définir un type, c'est-à-dire un organisme idéal possédant toutes les caractéristiques anatomiques essentielles de ce taxon sans tenir compte des particularités de chaque espèce en faisant partie. On dit alors que les cas particuliers qui représentent ces plans d'organisation sont des variations sur un même thème.
Les niveaux de l'empire et du règne
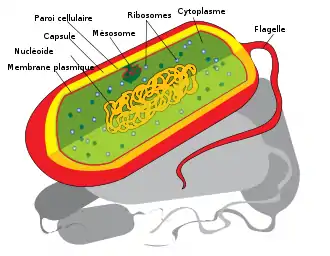

Aux niveaux hiérarchiques les plus élevés le plan d'organisation est défini par l'arrangement relatif des principales structures cellulaires. On compte ainsi deux empires dont l'organisation cellulaire est radicalement opposée : les procaryotes et les eucaryotes[4]. Chez les premiers la cellule est simplement constituée d'un cytoplasme entouré par une membrane plasmique puis généralement par une paroi dont la composition est variable (muréine, membrane externe, protéines, etc.). Chez les derniers la cellule possède un noyau qui isole le matériel génétique dans un compartiment exempt de synthèse protéine ribosomique et un système d'endomembranes qui permet un flux membranaire constant. Les eucaryotes possèdent de plus d'autres organites membranés comme la mitochondrie ou le péroxysome.
Lorsque l'on descend d'un niveau pour arriver à celui du règne, on précise davantage le plan d'organisation. Le règne bactérien est l'unique règne du domaine Prokaryota, les variations de plan observées ne sont pas suffisamment grandes pour justifier la création de règnes séparés, ce qui est en revanche le cas pour les 5 règnes eucaryotes :
- Le règne basal est celui des protozoaires, il représente le grade évolutif inférieur[5] ;
- Le règne animal s'en distingue par le passage à la pluricellularité et la différenciation cellulaire ;
- Le règne fongique par l'acquisition d'une paroi chitineuse, la perte de la phagotrophie et son remplacement par un système d'exodigestion et d'absorbotrophie ;
- Le règne plantal par l'acquisition ancestrale du chloroplaste par symbiogenèse avec une cyanobactérie ;
- Le règne chromiste par l'endosymbiose secondaire d'une algue rouge qui augmente la complexité de la topologie membranaire.
Le niveau de l'embranchement

C'est le niveau le plus classique pour l'étude de la diversité du vivant. En ancienne classification, chaque phylum, appelé embranchement en zoologie ou division en botanique, était caractérisé par une description schématique appelée « plan d'organisation »[6]. On distingue une soixantaine d'embranchements et donc de plans d'organisation.
Si on compare par exemple arthropodes et chordés on retrouve des points communs comme l'existence d'une symétrie bilatérale avec deux axes de polarité antéropostérieur et dorsoventrale (ce sont deux embranchements du sous-règne Bilateria) et un corps au moins partiellement métamérisé. Mais par la suite on observe des différences importantes. Le corps des chordés est constitué de trois parties, une tête, un tronc et une queue post-anale, il est peu métamérisé (somites seulement), le système nerveux est dorsal (épineurien), le cœur est ventral et le squelette est interne (chorde ou squelette osseux).
Au contraire, le corps des arthropodes est très métamérisé, constitué de segments répétés visibles en morphologie externe (structure annelée des myriapodes par exemple) parfois rassemblés en trois parties plus ou moins distinctes : l'abdomen, le thorax et le céphalon. Le système nerveux est ventral (hyponeurien), le cœur est dorsal, le squelette formé de chitine est externe, et certains segments portent des appendices articulés.
Les niveaux inférieurs
Lorsqu'on continue à descendre dans les niveaux hiérarchiques de la classification, on continue à préciser toujours davantage le plan d'organisation de plus en plus particulier auquel on a affaire.
Par exemple chez les tétrapodes, une superclasse parmi les chordés, on peut rajouter au plan général un crâne, quatre membres chiridiens et une paire de poumons pour le système respiratoire. Plus bas dans la classification, on distingue par exemple la famille des hominidés de celle des pongidés grâce à la bipédie et aux adaptations anatomiques associées.
Bases biologiques : Les réseaux de régulation génétique (RRG)
Que sont les RRG?
Les réseaux de régulation génétique sont très importants dans le domaine des plans de corps parce qu’ils déterminent le cours du développement d’un animal. Il est possible de dire que ces réseaux sont en fait les liens fonctionnels entre les gènes régulateurs qui produisent les facteurs de transcription et leur cibles, ainsi que d’autre gènes qui expriment des composantes signalétiques très importantes. On dit que la structure de ces réseaux de régulation génétique est hiérarchique parce que le développement d’un animal comporte plusieurs stades; à chacun de ces stades agissent des réseaux de régulation génétique. Ces réseaux auront plus d’effet sur les plans d'organisation s’ils arrivent à un stade précoce du développement parce que la modification aura une influence sur toutes les étapes subséquentes du développement. Au contraire, une modification dans un réseau de régulation génétique qui agit sur des évènements terminaux aura moins de conséquences sur le reste de la cascade de développement. La structure des réseaux de régulation génétique est aussi modulaire. Cette caractéristique permet aux erreurs et aux perturbations de se limiter à un seul module, et ainsi de ne pas affecter tout le réseau, situation qui pourrait créer des erreurs de développement létales[7],[8].
Les composantes du RRG
On compte quatre composantes principales dans les réseaux de régulation génétique, chacune ayant son propre niveau d’effet en termes de plans d'organisation: les réseaux de régulation génétique au niveau du noyau, les plugins et les interrupteurs des réseaux de régulation génétique ainsi que les batteries de différenciation des gènes[7],[8].
Noyaux des RRG
Les réseaux de régulation génétique au niveau du noyau sont en lien avec les caractéristiques des plans d’organisation au niveau des phylums et les super-phylums. Les modifications et les variations à ce niveau des plans d'organisation sont très rares. On parlerait de variations au niveau de l’organisation des parties du corps (ou la présence et l’absence des parties du corps) ou encore de l’organisation du système nerveux. Dû à la haute importance des réseaux de régulation génétique au niveau du noyau, les variations et les changements à ce niveau sont très rares[7],[8].
Le développement de tous les animaux est contrôlé par le même ensemble de gènes : les gènes homéotique Hox. Les gènes Hox agissent au niveau des noyaux des réseaux de régulation génétique. C’est cet ensemble de gène qui contrôle le développement des embryons sur l’axe antéro-postérieur. Par la suite, les protéines Hox confèrent le type de structures que les segments développeront. Les gènes régulateurs Hox déterminent où, quand et à quelle intensité et pendant combien de temps les segments se développeront, résultant ainsi dans la grande diversité des plans d’organisation des animaux que nous connaissons aujourd’hui.
La fonction de certains gènes Hox a évoluer, au fil du temps. Cet évènement se produit souvent après la duplication du gène. Avoir une copie du gène Hox confère plus de liberté au niveau des fonctions de cette copie, pouvant ainsi mener à l’apparition de nouvelles fonctions. Un bon exemple de l’évolution des fonctions à partir du gène Hox est celui de la drosophile. Chez Drosophila, la duplication du gène Hox, a entrainé l’apparition d’un nouveau gène, Zen, qui occupe maintenant la fonction de la détermination des types de tissus dans l’embryon. Encore plus récemment dans l’évolution, la duplication du gène Zen a mené à l’apparition du gène Bicoid qui détermine l’axe de la tête et de la queue du corps de Drosophila[3].
Plugins et interrupteurs des RRG
Les plugins et les interrupteurs des réseaux de régulation génétique sont en lien avec les caractéristiques des plans d’organisation au niveau des classes, des ordres et des familles. Il serait question, par exemple, de modifications au niveau des signaux des systèmes de transduction, ou encore au niveau des patrons d’ailes[7].
Batterie de différenciation des gènes
Les batteries de différenciation des gènes sont en lien avec les caractéristiques des plans d’organisation au niveau de la spéciation. Dans ce niveau de réseau de régulation des gènes, nous avons par exemple, les gènes qui codent des protéines qui vont engendrer des changements au niveau de la composition minérale du squelette, de la peau, des cellules musculaires ou encore les synapses au niveau du système neural. Les batteries de différenciation des gènes, contrairement aux 3 structures énoncées plus haut, ne sont pas régulatrices[7],[8].
Les conséquences évolutives des modifications du RRG
Une ou plusieurs modifications dans le circuits, par exemple une mutation sur un gène ou sur une séquence régulatrice, peut avoir des conséquences sur les résultats du développement de l’organisme, et ultimement avoir des conséquences sur l’évolution. Cela dit, dans le but de conservation, les gènes évoluent à des rythmes différents, qui sont proportionnels à la hiérarchie dans les RRG : plus les gènes sont hauts dans la hiérarchie des RRG (par exemple les gènes régulateurs dans les noyaux) moins ils sont soumis aux variations. Au contraire, plus les gènes sont bas dans la hiérarchie des réseaux de régulation des gènes (par exemple au niveau des batteries de différenciation des gènes) plus ils sont enclins aux variations. C’est la structure hiérarchique des réseaux de régulation des gènes qui explique l’évolution des plans d’organisation que nous trouvons aujourd’hui. Les gènes les plus hauts dans la hiérarchie, qui déterminent les aspects généraux d’un phylum, sont très peu soumis aux mutations et aux variations génétique, ce qui explique la conservation des plans d’organisation de base des animaux d’un phylum. À l’inverse, puisque les gènes les plus bas dans la hiérarchie des RRG déterminent les caractéristiques de la spéciation et qu’ils sont le plus enclins aux variations génétiques, nous observons une grande diversité de caractéristiques[3].
Plan d'organisation et évolution
Convergence évolutive et adaptation

Le plan d'organisation, en tant que structure fonctionnelle, représente une forme d'adaptation de l'organisme à sa niche écologique. On observe ainsi certaines corrélations entre le plan d'organisation et le mode de vie ou la zone adaptative. Par exemple, la présence d'un seul axe de polarité (et donc d'une symétrie radiale) est le plus souvent associée avec un mode de vie fixé ou errant (éponges, méduses, plantes terrestres). Tandis que la présence de deux axes de polarité (et donc d'une symétrie bilatérale) est associée avec une mobilité active (animaux bilatériens).
Des organismes appartenant à des groupes très différents, mais soumis à des pressions sélectives similaires peuvent donc adopter des plans d'organisation plus ou moins similaires : c'est la convergence évolutive (voire le cas des ailes ci-contre).
Au contraire, deux plans d'organisation similaires peuvent mener à des modes de vie très différents (par exemple les cétacés et les oiseaux répondant à un plan d'organisation proche) alors que deux plans d'organisation très différents peuvent permettre des modes de vie similaires (éponges et ascidies).
Divergence évolutive et essentialisme
Au sein d'un taxon, le plan d'organisation général peut être plus ou moins modifié dans le cas particulier d'un taxon d'ordre inférieur. L'ampleur de cette modification (quantité et qualité des caractères modifiés, et valeur adaptative de ces derniers) peut entraîner la catégorisation de ce taxon au même niveau linnéen (voire un niveau supérieur) que le taxon ancestral (dans le cadre de la pratique taxonomique évolutionniste). Ce n'est pas le cas lorsque la modification (même importante) ne touche qu'un faible nombre de caractères. Cette divergence évolutive peut inclure une régression vers un grade antérieur.
Par exemple, bien que le plan d'organisation général du règne des chromistes inclut la présence d'un chloroplaste à quatre membranes, certaines chromistes ont secondairement perdu ce plaste (régressant ainsi plus ou moins vers un plan d'organisation « de type protozoaire ») comme les embranchements des ciliés ou des pseudochampignons (on appelle chromophytes les chromistes qui au contraire ont conservé le plan d'organisation ancestral permettant la photosynthèse). Un autre exemple connu concerne par exemple la régression du sous-règne animal Myxozoa vers l'unicellularité. Dans ce cas, bien que les myxozoaires émergent généalogiquement parmi les radiés, ils ne font pas partie de ce sous-règne à cause l'ampleur de la modification phénotypique. Comme dans le cas des myxozoaires, la régression du plan d'organisation est souvent liée à un mode de vie parasitaire.
Le plan d'organisation ne désigne donc pas une essence fixe et éternelle d'un taxon (qui est de toute façon soumis aux transformations qu'implique l'évolution biologique), c'est-à-dire un ensemble de caractères qui le définirait exclusivement (aucun autre groupe ne posséderait un de ces caractères) et nécessairement (tous les organismes de ce taxon posséderaient tous ces caractères). Il représente simplement un type idéalisé représentant les traits morphoanatomiques que l'on rencontre généralement chez les organismes appartenant à ce taxon.
Histoire évolutive

Les premiers fossiles d’animaux datent de 550-530 millions d’années. Cette période, la période du Cambrien, est la plus grande évolution que nous connaissons à ce jour. Tous les représentants des phylums modernes sont apparus dans le Cambrien. Sur le plan évolutif, la plupart des plans d'organisation animaux sont apparus pendant l'explosion cambrienne. Il y a eu depuis relativement peu d'innovations (aucune apparition d'un nouvel embranchement), ce qui laisse supposer que des contraintes importantes existent et qui auraient pour conséquence de rendre toute modification importante du plan d'organisation peu avantageuse sur le plan évolutif. En biologie évolutive du développement, on définit la contrainte comme étant la limite à la variation ou la canalisation de la variation. Pour que la sélection agisse sur un organisme, il doit y avoir variation. Comme la contrainte agit sur la limite possible à la variation, la sélection est donc, elle aussi, limitée par les contraintes. Les contraintes sont mises en place pour s’assurer que des structures essentielles pour la survie ne soient pas soumises aux variations durant leur développement. Ces contraintes sont maintenues en place par la sélection stabilisante. Par opposition aux contraintes, existe le concept d’évolutivité, défini par l’habileté à une population de produire des variations qui sont plus adaptées que celles qui existent déjà[3].
| Animaux |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
- Les types d'organisation présentés ici sont des grades évolutifs ne correspondant généralement pas à des groupes monophylétiques, mais paraphylétiques (ne comportant pas tous les descendants d'un même ancêtre — exemple : les descendants d'ancêtres vermiformes ne sont pas tous aujourd'hui des vers, etc.).
- En jaune : les principales explosions radiatives.
Plans d'organisation polyphylétiques
Les groupes définis par leur plan d'organisation ne sont pas toujours monophylétiques. Le plan d'organisation est en effet défini à partir d'un ensemble de caractères morphoanatomiques pouvant dans certains cas correspondre à un ensemble d'analogies. Par exemple les systématiciens classaient autrefois les plans d'organisation animaux en fonction de la présence ou de l'absence d'une cavité générale car ils pensaient qu'il s'agissait d'un critère généalogiquement pertinent. Aujourd'hui on sait que les groupes ainsi formés (cœlomates, pseudocœlomates et acœlomates) sont polyphylétiques, la perte du cœlome étant probablement un cas de convergence évolutive dans plusieurs groupes. De même, quand on parle des organismes vermiformes on fait référence à un groupe polyphylétique. C'est le cas aussi de certains grades évolutifs comme l'unicellularité qui est également polyphylétique.
Plan d'organisation et taxonomie
En règle générale, on appelle « embranchement » un taxon dont le plan d'organisation est si différent des autres qu'on le considère comme un plan d'organisation original.
Comme expliqué ci-dessus, un plan d'organisation ne désigne cependant pas nécessairement un clade. En effet, les taxons étant tous apparentés les uns aux autres, l'apparition d'un nouveau plan d'organisation se produit nécessairement à partir d'un plan d'organisation précédent. Ainsi le plan d'organisation primitif ne peut que renvoyer à un groupe paraphylétique.
Par exemple, on peut citer le taxon Acanthocephala qui a un plan d'organisation proche des Aschelminthes, alors qu'il est apparu au sein des Rotifera. Citons aussi le cas de Siboglinidae dont le plan d'organisation rappelle une organisation « deutérostomien/epithélioneurien » mais qui sont en fait des Annelida. Aujourd'hui, on tendra alors à considérer les Syndermata comme l'ensemble rassemblant acanthocephales et rotifères où les rotifères sont paraphylétiques. L'embranchement syndermata est alors constitué de deux plans d'organisations dont l'un est primitif (rotifères) et l'autre dérivé du premier. De même, les Siboglinidae sont considérés comme appartenant aux annélides qui consensuellement comprennent au moins 5 plans d'organisation différents : Echuria, Myzostomida, Sipuncula, Siboglinidae et Annelida.
On comprend alors que le plan d'organisation ne doit pas être considéré comme une essence (ce qui serait une régression aux conceptions fixistes de Cuvier).
Position des écoles de taxonomie
Les écoles de pensée de taxonomie s'opposent sur la prise en compte ou non du plan d'organisation dans l'établissement de la classification scientifique des espèces, c'est-à-dire sur le fait qu'un grade puisse être retenu comme élément de taxinomie. Si les cladistes et les évolutionnistes s'accordent sur l'exclusion des groupes polyphylétiques fondés sur des analogies, ils se disputent en revanche sur le statut des groupes paraphylétiques.
- Pour les cladistes, tout taxon doit être un clade (donc monophylie) et tout clade est un taxon. Les grades et le plan d'organisation n'ont donc pas de pertinence dans la classification phylogénétique, organisant celle-ci uniquement à partir de synapomorphies (les plus récents d'entre eux étant souvent obtenus à partir d'arbres par les techniques de phylogénie moléculaire, sur lesquels on définit a posteriori des synapomorphies).
- Au contraire pour les évolutionnistes la classification doit refléter la dynamique évolutive réellement à l’œuvre et les changements de plan d'organisation, et les regroupements entre espèces ont d'autant plus de sens qu'ils reflètent des plans d'organisations comparables ; ce qui revient reconnaître comme valide des taxons paraphylétiques.
Il s'agit bien uniquement d'une différence de philosophie dans l'approche de la classification scientifique, puisque l'école évolutionniste ne rejette pas, bien au contraire, l'utilisation des techniques de phylogénie moléculaire et l'établissement de cladogrammes (qui ne sont alors considérés que comme des données brutes devant être traduites en une classification plus fine, c'est-à-dire ayant un pouvoir explicatif quant à l'histoire de la vie). Inversement, pour discuter des regroupements et différenciations entre espèces, les deux écoles s'appuient sur des comparaisons morphologiques, anatomiques, voire écologiques qui n'ont de sens que par rapport à ces plans d'organisation.
Voir aussi
Articles connexes
- Biologie du développement
- Évolution des plans d'organisation des animaux
- Gène Hox
- Réseaux de co-expression de gènes
- Réseau de régulation génique (en)
Liens externes
Bibliographie
- Atlas de biologie animale - tome 1, Heusser S. & Dupuy H.-G.
- Manuel M. 2009. Évolution animale : les péripéties de la phylogénie. Encyclopédie Universalis, 134-149.
Références
- Thomas Cavalier-Smith, 1998. A revised six-kingdom system of life. Biol. Rev. 73: 203-266.
- Chanson B (1992) Hétérogénéités angulaires des déformations de maturation. interprétation basée sur le concept de plan d'organisation des arbres, In Quatrième séminaire ASMA (Architecture, structure et mécanique de l'arbre), université de Montpellier, 24-25 février 1992, p. 120-131
- (en) Katherine E. Willmore, « The Body Plan Concept and Its Centrality in Evo-Devo », Evo Edu Outreach, no 5, , p. 219-230
- Aubert D. La transition bimembranées/unimembranées : une révolution au royaume des bactéries ? 2013. <hal-01063767>
- (en) Cavalier-Smith T, « Kingdoms Protozoa and Chromista and the eozoan root of the eukaryotic tree », Biol. Lett., vol. 6, no 3, , p. 342–5 (PMID 20031978, DOI 10.1098/rsbl.2009.0948, lire en ligne)
- Daniel Richard (sous la direction de), Patrick Chevalet, Sylvie Fournel, Nathalie Giraud, Frédéric Gros, Patrick Laurenti, Fabienne Pradere, Thierry Soubaya, Biologie : Tout le cours en fiches, 2e éd., Dunod, Paris, 2012, p. 83. (ISBN 978-2-10-058878-7)
- (en) Davidson E., Erwin D., « Gene Regulatory Networks and the Evolution of Animal Body Plans », Science, no 311,
- (en) He J., Deem M., « Hierarchical Evolution of Animal Body Plans », Developmental Biology, no 337, , p. 157-161
 Portail de la zoologie
Portail de la zoologie  Portail de la botanique
Portail de la botanique