Chloroplaste
Les chloroplastes sont des organites présents dans le cytoplasme des cellules eucaryotes photosynthétiques (plantes, algues). Ils sont sensibles aux expositions des différentes ondes du spectre lumineux. Ils jouent un rôle essentiel dans le fonctionnement d'une cellule végétale étant donné qu'ils permettent de capter la lumière à l'origine de la photosynthèse. Par l'intermédiaire de la chlorophylle qu'ils possèdent et de leurs ultrastructures, ces organites sont capables de transférer l'énergie véhiculée par les photons à des molécules d'eau. Les chloroplastes jouent un rôle important dans le cycle du carbone, par la transformation du carbone atmosphérique en carbone organique. Les chloroplastes appartiennent à une famille d'organites appelés les plastes ; ceux-ci sont le fruit de l'endosymbiose d'une cyanobactérie, il y a environ 1,5 milliard d'années.
-fr.svg.png.webp)
Historique
Le chloroplaste a été découvert au cours de recherches scientifiques menée sur des plantes. Les premières recherches ont commencé par Joseph Priestley en 1772. Il s'était intéressé à l’étude des gaz et en identifia plusieurs. Plus tard, il démontra que les plantes sont capables de régénérer les gaz qui viennent des animaux.
Quatre ans plus tard, Jan Ingenhousz reprend les travaux de Priestley et montre que le dégagement de dioxygène se produit uniquement à la lumière. Pendant la nuit, les plantes rejettent un gaz rendant impossible la combustion d’une bougie.
À la fin du XVIIIe siècle, les recherches ont conclu que, comme les animaux, les plantes respirent. En 1837, Henri Dutrochet découvre que le pigment vert dans les feuilles est la chlorophylle. En 1862, Julius von Sachs prouve que l’assimilation chlorophyllienne se déroule dans des chloroplastes. Ce n'est qu'en 1898 que le scientifique Charles Reid Barnes invente le terme photosynthèse.
Localisation

Les chloroplastes se situent dans le cytoplasme des cellules eucaryotes photosynthétiques. Chez les algues du genre Chlamydomonas, il y a un chloroplaste par cellule, généralement en forme de cloche et occupant une part importante du cytoplasme.
Chez les végétaux supérieurs, la plupart des parties aériennes en contiennent, ce sont les feuilles (caduques ou non) qui en contiennent le plus., Leur lieu de prédilection est tout particulièrement le mésophylle de la feuille où chaque cellule peut contenir plusieurs dizaines de chloroplastes. Divers tissus aériens en contiennent peu : les cellules de revêtement de l'épiderme, les cellules stomatiques aquifères (des groupes de cellules qui évacuent de l'eau chez certaines feuilles). L'écorce et les pétioles en contiennent un peu[1], mais en général les cellules de tissus non exposés à la lumière, comme les racines, n'en comportent pas.
Structure
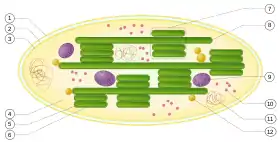
(1) membrane externe ;
(2) espace intermembranaire ;
(3) membrane interne ;
(4) stroma ;
(5) lumen du thylakoïde ;
(6) membrane du thylakoïde ;
(7) granum (empilement de thylakoïdes) ;
(8) thylakoïde ;
(9) amidon ;
(10) ribosome ;
(11) ADN chloroplastique ;
(12) plastoglobule (gouttelette lipidique).
La taille des chloroplastes est de l'ordre du micromètre (µm). Ils prennent souvent la forme de disques aplatis de 2 à 10 µm de diamètre pour une épaisseur d'environ 1 µm. Le chloroplaste est un organite composé de deux membranes séparées par un espace inter-membranaire (2). Il contient un réseau membraneux constitué de sacs aplatis nommés thylakoïdes (8) qui baignent dans le stroma (4) (liquide intra-chloroplastique). Les thylakoïdes sont composés d'un lumen (espace intra-thylakoïde, 5) entouré d'une membrane (6), et contiennent de la chlorophylle (pigments verts) et des caroténoïdes (pigments jaune orange). Un empilement de thylakoïdes se nomme granum (7) (au pluriel : des grana). D'autre part, le stroma contient quelques réserves sous forme d'amidon (9), et des structures lipidiques dont le rôle est encore mal compris, les plastoglobules(12).
Génome chloroplastique
De plus, ces organites contiennent de l'ADN regroupé en nucléoïdes (11) ; chaque chloroplaste peut contenir jusqu'à 100 copies du génome[2]. Les molécules d'ADN du génome chloroplastique sont généralement linéaires ou ramifiées[3]. Le génome chloroplastique est très réduit, 37 à 220 kb et contient généralement une centaine de gènes, alors que par comparaison celui d'une cyanobactérie (origine des chloroplastes) fait plusieurs mégabases et comporte plusieurs milliers de gènes.
Les ribosomes (10) sont constitués d'ARNr, synthétisés dans les chloroplastes, et de protéines codées par les génomes nucléaires et chloroplastiques.
L'ADN du chloroplaste ne lui permet pas de subvenir à tous ses besoins ; il y a une coopération entre la cellule et les chloroplastes, analogue aux relations entre la cellule et ses mitochondries. Par exemple, la Ribulose 1,5 Bisphosphate Carboxylase/Oxygénase (ou Rubisco) est composée de deux parties : une grande et une petite qui sont répétées chacune huit fois. La grosse sous-unité (55 kDa) est formée dans le chloroplaste et la petite sous-unité (15 kDa) est synthétisée dans le cytoplasme de la cellule, sous la forme de précurseurs, puis pénètre dans le chloroplaste. On estime qu'il s'agit de la protéine la plus abondante de notre biosphère. En règle générale, les protéines codées par l'ADN nucléaire mais destinées à une localisation chloroplastique sont synthétisées sous la forme de précurseurs pourvus d'un signal de transit en N-terminal, qui consiste en un peptide d'une cinquantaine d'acides aminés (taille variable d'une protéine à une autre), et qui sera clivé au moment du passage à l'intérieur du chloroplaste pour aboutir à la protéine mature. Les protéines destinées à la membrane des thylakoïdes du chloroplaste peuvent même être pourvues de deux séquences de transit, la première pour entrer dans le chloroplaste, la deuxième pour être intégrée dans la membrane. Ces mécanismes font intervenir deux complexes protéiques connus sous le nom de TIC et TOC.
Rôle

Le chloroplaste, est le siège de la photosynthèse. Il absorbe l’énergie lumineuse pour fixer le carbone inorganique (CO2) sous forme de glucose, au cours de ce processus de l'énergie chimique sous forme d'adénosine triphosphate (ATP) est également produite. Cet ATP intervient dans la phase photochimique de la photosynthèse.
Le chloroplaste absorbe l'ensemble du spectre de la lumière visible. La chlorophylle se trouve dans la membrane des thylakoïdes. Les différentes étapes de la photosynthèse qui convertissent la lumière en énergie chimique se déroulent dans les thylakoïdes tandis que les étapes de conversion de l'énergie en glucide se déroulent dans le stroma du chloroplaste.
Le chloroplaste joue aussi un rôle dans la fixation du carbone, de l'azote, du soufre ou encore de la biosynthèse des lipides.
L'une des protéines impliquées dans la protection du chloroplaste est la protéine OsCEST[4].
Transmission
Chez les gymnospermes, les chloroplastes se transmettent à la descendance par le pollen, alors que chez la majorité des plantes à fleurs la transmission se fait par l'ovule. Cependant, il y a beaucoup de cas documentés actuellement, d'angiospermes héritant leurs chloroplastes de leur parent mâle, donc par le pollen.
Origine
Les chloroplastes sont le résultat d'une endosymbiose, c’est-à-dire que des cellules eucaryotes primitives ont ingéré des cyanobactéries il y a 1,5 ou 1,6 milliard d'années, puis ont vécu en symbiose avec ces dernières[5]. Par comparaison les mitochondries proviennent de l'endosymbiose d'une alphaprotéobactérie par une cellule primitive il y a environ 2 milliards d'années.
Il y a deux types d'endosymbiose :
- endosymbiose primaire : une cellule eucaryote ingère une bactérie, celle-ci devenant un chloroplaste avec deux membranes ayant pour origine la membrane de la bactérie pour la membrane interne, la membrane cytoplasmique pour la membrane externe (Rhodophyta et Chlorobionta),
- endosymbiose secondaire : une cellule eucaryote phagocyte une autre cellule eucaryote possédant un chloroplaste ; le cytoplasme et le noyau dégénèrent pour ne laisser que le chloroplaste à quatre membranes (deux membranes issues de l'endosymbiose primaire, la membrane cytoplasmique de la cellule phagocytée, et la membrane invaginée de la cellule qui phagocyte).
À la suite d'une réduction du nombre de membranes, on connaît aussi des chloroplastes à trois membranes chez certains Dinophytes.
On distingue une lignée verte et une lignée rouge selon que les plastes d'endosymbiose secondaire sont issus d'algues respectivement verte ou rouge.
Comparaison


- Proplaste
- Plaste
- Chloroplaste et étioplaste
- Chromoplaste
- Leucoplaste
Notes et références
- Pilarski, J. (1993). Ultrastructure of chloroplasts in the bark and leaves of lilac (Syringa vulgaris L.). Acta physiologiae plantarum, 15(4), 241-248.
- (en) Bendich AJ, « Why do chloroplasts and mitochondria contain so many copies of their genome? », BioEssays, vol. 6, no 6, , p. 279-82 (PMID 3619888)
- (en) Bendich AJ, « Circular chloroplast chromosomes: the grand illusion. », the Plant Cell, vol. 16, no 7, , p. 1661-6 (PMID 15235123, lire en ligne)
- A novel chloroplast protein, CEST induces tolerance to multiple environmental stresses and reduces photooxidative damage in transgenic Arabidopsis, 1. Naoki Yokotani1,*, 2. Mieko Higuchi2, 3. Youichi Kondou2,†, 4. Takanari Ichikawa2,‡, 5. Masaki Iwabuchi1, 6. Hirohiko Hirochika3, 7. Minami Matsui2 and 8. Kenji Oda1,$
- (en) McFadden, G., Gilson, P., Hofmann, C., Adcock, G. and Maier, U, « Evidence that an Amoeba Acquired a Chloroplast by Retaining Part of an Engulfed Eukaryotic Alga », PNAS, vol. 91, , p. 3690-3694
Voir aussi
Bibliographie
- Henri Camefort, Morphologie des végétaux vasculaires : cytologie.anatomie.adaptations, Doin, 1996, 2e éd. (7e tirage), 432 p., p. 32-49 (ISBN 2-7040-0395-5)
- cours de DEUG en ligne.[PDF]
- (en) Targeting of nucleus-encoded proteins to chloroplasts in plantsArticle scientifique de synthèse
Articles connexes
Liens externes
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire  Portail de la botanique
Portail de la botanique