Microbiote cutané humain
Le microbiote cutané humain (ou microflore de la peau humaine) est la communauté de micro-organismes opportunistes, saprophytes, souvent symbiotes[1] et parfois pathogènes qui composent la « flore cutanée ». C'est la partie externe du microbiote de l'organisme humain. De manière générale, la biodiversité bactérienne limite le risque de colonisation de la peau par une bactérie pathogène et constitue une protection contre l'inflammation de la peau [2]. Une partie du microbiote cutané est partagé avec le microbiote des muqueuses, notamment respiratoires[3].
Éléments de description
En raison de la taille microscopique des organismes qui la composent, la microflore humaine est invisible à nos yeux.
La peau d'un adulte héberge en moyenne 1012 (mille milliards) bactéries (soit un million par cm2 de peau)[4] de plus de 500 espèces différentes[5].
La peau supporte et entretient naturellement son propre « micro-écosystème ». Celui-ci se forme à la naissance, puis évolue jusqu'à la mort. Cet « écosystème » est étudié depuis quelques décennies en tant que tel.
Pour partie anaérobie[6] et caché dans les profondeurs de la peau, il est encore mal connu, mais on sait que :
- il est organisé en biofilm et s'alimente à la fois de molécules et de composés excrétés par la peau elle-même, et de composés sécrétés par des communautés de microorganismes plus ou moins symbiotes ;
- ces organismes sont spécialisés ou opportunistes. Ce sont surtout des bactéries mais aussi des micro-champignons, des acariens (dont Demodex brevis et Demodex folliculorum, des micro-nématodes. Ils forment notre « flore cutanée » (commensale et transitoire) ;
- la composition de cette « flore cutanée » varie selon les individus, leur âge, leur sexe, leurs activités, leur comportement et l'environnement[7] ;
- cet écosystème cutané est organisé en biofilm[8] ;
- sa composition et son « épaisseur physique » varie selon l'individu, mais aussi selon les parties du corps ; La microflore du cuir chevelu diffère[9] de celle du visage, du dos, des aisselles, des pieds, des dessous d'ongles ou des organes génitaux, de l'anus, etc. Nulle part sur la peau n'existe une flore qui rassemblerait la totalité de la flore cutanée (Marples, 1969). Les zones corporelles les plus riches en bactéries sont (en ordre décroissant) : les mains, le cuir chevelu, les aisselles, le front, les membres et le dos[9]. Cette microflore est aérobie en surface de la peau et anaérobie dans ses parties profondes[10] ;
- cette flore est pour une petite partie en continuité avec celles des muqueuses épithéliales internes qui prolongent la peau à l'intérieur du corps, mais ces muqueuses accueillent surtout d'autres communautés d’organismes, très différentes et qui, pour ce qui du nombre et du volume, constituent la majeure partie du microbiote de l'organisme humain. Ce microbiote « interne » forme également des biofilms qui jouent un rôle essentiel pour la digestion. Une déficience du biofilm de la muqueuse colique joue un rôle dans les maladies inflammatoires de l’intestin[11] ;
- le biofilm cutané doit se renouveler constamment, pour s'adapter à la desquamation[12] naturelle et à l'érosion différenciée de la peau, ainsi qu'aux pratiques d'hygiène corporelle (qui ne doivent pas être excessives pour protéger les fonctions "barrière" de la peau ni réduire la diversité du microbiote qui protège de l'inflammation[2]) ;
- chez l'enfant, le biofilm bactérien est d'abord constitué de staphylocoques dont Staphylococcus aureus, très peu de Escherichia coli et corynébactéries. À l’adolescence, cette flore se complète d'autres coques[9] comme les Staphylocoques. Cette microflore joue un rôle dans l'odeur de la peau humaine, qui évolue de la naissance à la mort[9].
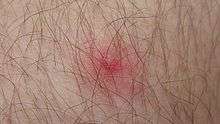
Fonctions du biofilm cutané

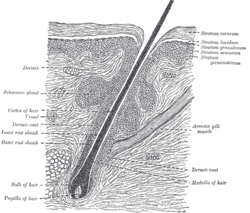
_Dyshidrotic.JPG)
Il joue un rôle tantôt positif, tantôt négatif :
- les micro-organismes qui vivent sur et dans la peau humaine et leur diversité participent au contrôle de l'équilibre qui caractérise une peau « saine »[9]. Ce biofilm commensal « joue un véritable rôle de protection de l’organisme face à des infections. Elle n’est, à l’état normal chez un individu sain, à l’origine d’aucune infection. Ceci est dû la présence d’une véritable homéostasie bactérienne[8] » ;
- Ils participent à l'apparition des odeurs corporelles qui sont dues essentiellement à des acides gras courts, tels que l'acide caprilique, caprique, valérianique, ou l'acide propionique, produit de dégradation des acides aminés et des acides gras à chaîne carbonée longue par les bactéries du genre Brevibacterium et Propionibacterium [13].
- en cas de déséquilibre ou de blessure, le biofilm peut évoluer en une situation pathologique[14].
Il est positif pour la cicatrisation et l'homéostasie de la peau. Indirectement, ces bactéries entretiennent notre système immunitaire, stimulent la phagocytose et la production d'anticorps ainsi que de cytokines (Chiller, 2001)[réf. insuffisante]. De plus la microflore symbiote sécrète des bactériocines, polypeptides bactériens aux propriétés bactéricides et bactériostatiques qui empêchent d'autres bactéries, plus souvent pathogènes (Gram-positives) de coloniser la peau. L'activité bactérienne peut cependant, quand elle est dérégulée, contribuer à empêcher la cicatrisation (plaie chronique)[15] ou à l'apparition ou à la perpétuation de diverses dermatoses, d'abcès. Les bactéries du biofilm cutané ne deviennent pathogènes que chez les immunodéprimés (SIDA/VIH, toxicomanes, traitement immunosuppresseur, nouveau-né…), ou parfois chez les patients sains, en cas rupture de la barrière cutanée ou des muqueuses[16] : blessures, brûlures, etc. Ce sont souvent des bactéries de la peau, naturellement plutôt résistantes, qui sont sources de maladies nosocomiales en pénétrant l'organisme via les cathéter, ou par colonisation de zones vulnérables dont l'œil[17] ; par exemple, l'endophtalmie provient généralement de bactéries originaires de la peau péri-oculaire ou des cils[18] ; - le biofilm cutané humain est un sujet d'étude encore émergent[19], mais il y a consensus scientifique sur le fait que « la flore cutanée résidente, organisée sous forme de biofilms, a une présence bénéfique pour l’hôte, puisqu’elle le préserve de l’invasion et de la colonisation de bactéries pathogènes. Toute modification de l’équilibre de la flore bactérienne, autrement dit toute rupture de l’homéostasie bactérienne, entraîne des troubles cutanés et est à l’origine de dermatoses[8]. »
Bactéries
Les bactéries semblent jouer un rôle majeur parmi les micro-organismes du biofilm cutané.
Des évaluations quantitatives ont mesuré des populations bactériennes allant jusqu'à plus de cent mille bactéries par centimètre carré de peau humaine (taux variant fortement selon les zones de peau humaine (environ 1,9 m2). Les zones plus riches en sébum (visage notamment), accueillent plus d'un million de bactéries par centimètre carré. Ce nombre paraît élevé, mais l'ensemble de ces bactéries tiendraient toutes dans le volume d'un pois[20] .
Les bactéries les plus communes sur la peau humaine sont de type Gram-positives et appartiennent principalement à 3 genres[9],[21].
On trouve les bactéries suivantes:
- Staphylococcus, souvent opportunistes comme Staphylococcus epidermidis[16], qui constitue plus de 90 % de la flore résidente aérobie présente sur le stratum corneum[22]. D'autres staphylocoques qui ont été trouvés dans le biofilm cutané sont Staphylococcus aureus (commun, trouvé par exemple en portage asymptomatique dans les cavités nasales dans 20 % des cas[23]) et Staphyloccocus hominis ;
- Corynebacterium ;
- Propionibacterium (par exemple Propionibacterium acnes, Propionibacterium granulosum, Propionibacterium avidum). Propionibacterium acnes produit des acides gras issus en lipolysant le sébum. Ce faisant elle acidifie le milieu cutané, ce qui inhibe la croissance des colonies de Streptococcus pyogenes[24];
- Lactobacillus[10];
- Streptococcus ;
Les bactéries sont très présentes aussi sur les phanères (cheveux, poils, dessous des ongles). À partir du biofilm qui se forme naturellement sur les poils, elles peuvent pénétrer l'intérieur de la peau (et causer des problèmes inflammatoires en cas d'ongle incarné ou de poil incarné).
Champignons cutanés
- Ce sont par exemple Malassezia furfur et Candida albicans considérés comme symbiotes de la peau et qui se nourrissent du sébum et de la peau morte[9] ;
- Ils sont moins présents que les bactéries, mais peuvent causer des mycoses en cas de déséquilibre de l'hydratation ou du pH cutanés.
- En situation de déséquilibre pathogène de la peau et/ou de son biofilm protecteur, d'autres champignons, saprophytes notamment, peuvent coloniser la peau.
Acariens
Les plus typiques et sans doute les moins connus du grand public sont les demodex.
- Découverts en 1841, ils ont depuis été trouvés sur la peau de presque tous les humains (pour deux espèces) et pour d'autres espèces sur la peau de 11 des 18 ordres de mammifères euthériens)[25], acariens allongés qui semblent se nourrir du sébum produit par les glandes sébacées. Au moins deux espèces sont commensales de l'Homme : D. folliculorum et D. brevis.
- Selon le Dr Kui Zun, l'analyse de 48 années d'observations cliniques et d'études épidémiologiques (à partir de 1955 et correspondant à 905,801 participants de diverses origines ethniques, âges et professions) laisse penser que 97,68 % des humains adultes en abritent sans le savoir.
- Chez certaines personnes (ex. : immunodéprimées[26] ou après une chimiothérapie[26]…), ils pullulent en élargissant l'entrée des folicules pileux, en provoquant un blocage physique des follicules, ou probablement en agissant en tant que vecteurs pour les micro-organismes[27]. Ils peuvent alors favoriser la chute des cheveux et certaines affections de la peau dont l'acné rosacée[27] ; cette forme d'acné correspond toujours à des micropullulations (quadruplement en moyenne[27] avec une densité maximale d'acariens dans les pores des joues[27] et des pullulations plus marquées dans le cas des rosacées induites par la prise de stéroïdes[27]. La prise de tétracycline par voie orale durant un mois n'induit aucun changement dans le nombre des demodex comptés sur la peau touchée par une acné rosacée[27],[28]. Le demodex, bisexuel[réf. nécessaire], se reproduit au bout de 14 jours environ, avec un métabolisme et une viabilité dépendant notamment de la température[29]).
- Les espèces commensales de l'Homme ne semblent pas avoir d'hôtes intermédiaires (c'est-à-dire qu'ils passent directement d'Humain à humain) et sont dits « cosmopolites »[29] (Le nombre de demodex varie selon l'âge et la personne, ou sa condition de santé, mais on n'a pas trouvé de facteur ethnique, régional, climatique, professionnel ou sexuel qui puisse expliquer les différences dans les infestations naturelles ou pathologiques de demodex chez les humains).
- Un des moyens d'en collecter pour observation (au microscope en raison de leur petite taille) est d'appliquer un morceau de scotch sur la peau ou sur le cuir chevelu[29]. Selon des tests d'exposition à des températures de 5 à 37 °C, hors de la peau, plus la température est proche de 5 °C et plus ils vivent longtemps[29]. En moyenne, ils se meuvent en dessous de 0 °C, vivent moins bien au-dessus de 37 °C et meurent au-dessus de 54 °C (tous meurent au-dessus de 58 °C)[29]. La température optimale de survie de D. brevis est de 5 °C et elle est de 16 à 20 °C pour leur développement[29].
Sécrétions et signature olfactive de la peau humaine
La peau des mammifères est aussi un organe sécrétoire complexe.
Une phase aqueuse est sécrétée avec la sueur en plus de l'humidification transcutanée (« perte insensible en eau transcutanée » ou TEWL pour « Transepidermal water loss »[30]).
Un film hydrolipidique est constamment sécrété et renouvelé (émulsion sébum-sueur ; le sébum est excrété par les glandes sébacées. Les lipides (mélange d'acides gras polyinsaturés (25 %), de cholestérol (20 %) et de plus de 40 % de céramides[31] sont produits par les kératinocytes (et régulés par des enzymes dont la phospholipase A2 qui hydrolyse les phospholipides cutanés ; le dérèglement du fonctionnement de cet enzyme participe aux troubles[32] tels que psoriasis, acné, dermatite atopique, érythème dû aux rayons ultra-violets, eczéma, etc. C'est ce « film gras » qui s’étale au niveau du stratum corneum sur la phase aqueuse et produit une couche protectrice et bactériostatique[33] comparable au film oculaire[34]). Il rend toute la peau hydrophobe[34].
Une grande variété de particules (dont débris de peau morte) et de composés chimiques odorants s'en dégagent naturellement, ainsi que de la sueur et des sébums ; une part des composés participent directement à l'entretien de la peau et notamment à l'équilibrage de son pH (régulé par l’excrétion sudorale de composés chimiques acides comme l’acide lactique, l’acide undécylénique, l’acide urocanique, l'acide arachidonique, céramides et acides gras oméga-6 à longue chaîne[34] ; ils entretiennent le pouvoir tampon de la peau qui doit être assez acide pour repousser certains micro-organismes pathogènes[35] ; L’alcalinisation de la peau associée à certaines dermatoses permet une surinfection bactérienne, qui aggrave la dermatose existante[35]. Chez le chien, le stress peut rapidement alcaliniser la peau.
D'autres composés ou les mêmes sont des hormones et/ou des kairomones. Ils varient selon l'âge et selon le sexe de l'émetteur[36].
Les scientifiques ont à ce jour identifié environ 350 composés chimiques différents sécrétés par la peau et libérés dans l'air ou l'environnement, dont par exemple l’acide lactique, des acides gras à courte et longue chaîne, des aldéhydes, des alcools, des composés aromatiques, des amines, des acétates et des cétones, et des composés complexes, notamment sous les aisselles[37]. Ceux de ces composés qui sont volatils sont utilisés par de nombreux insectes ou acariens hématophages (poux, puces, tiques, moustiques, punaises, taons, mouches…) pour repérer leur hôte[38],[39].
Les odeurs de blessures, de pus, la gangrène peuvent également attirer certains insectes, dont mouches (quelques espèces produisent des asticots qui nettoient les plaies (voir asticothérapie).
Une fois le corps humain mort, son odeur change et attire très rapidement les insectes nécrophages.
Classification : on peut distinguer ces composés selon leur quantité, selon leurs caractéristiques physicochimiques ou biochimiques ou encore selon leur origine et caractère « primaire » ou « secondaire » (en deux grandes catégories et quatre sous-catégories) :
- composés endogènes : c'est le cas de la plupart de ces molécules. Elles résultent du métabolisme de la peau ou du corps (sécrétions des glandes de l’épiderme essentiellement). Il peut s'agir de composés primaires (émis tels quels) ou secondaires (dégradés par la microflore, par l'oxydation à l'air, par la température, la lumière ou les rayons UV solaires ;
- composés exogènes : ce sont les molécules indirectement produites par la peau, en réalité par les organismes (la « microflore ») qui y vivent, à parti de leur métabolisme propre. Il peut aussi s'agir de composés primaires (émis tels quels) ou secondaires. Les composés secondaires sont très nombreux, et pas toujours propres à l'espèce humaine, ni mêmes caractéristiques d'une origine animale ;
Le mélange de ces différents types de molécules forme une « signature odorante corporelle » propre à l'espèce et aux individus (signature que certains chiens peuvent finement différentier et suivre grâce à leur flair). C'est aussi ainsi que chez de nombreuses espèces, une mère reconnait avec certitude son petit et inversement. Il existe aussi une signature propre à chaque espèce ou type de vertébrés[40].
L'être humain dispose de sa « signature odorante » ; la peau humaine semble unique pour son niveau très élevé de triglycérides, en grande partie différent de ceux produits dans les tissus internes, ce qui laisse supposer qu'ils y ont une fonction. Ces triglycérides sont principalement transformés par des bactéries du genre propionibacterium en un large nombre d’acides gras libres à courtes et longues chaines.
Nombre de ces molécules pourraient jouer un rôle de kairomones - messagers / stimuli susceptible d'activer l'attention et l'attraction d'arthropodes hématophages (se nourrissant de sang ou de chair). C'est le cas par exemple de l'acide lactique[41] et de plusieurs acides gras[42],[43].
Principaux composés de l'odeur humaine
Ce sont notamment[37]:
Des acides :
- n-hexanoïque[37]
- 2-méthylhéxanoïque[37]
- 3-méthylhéxanoïque[37]
- 4-éthylpentanoïque[37]
- (Z)-3-méthyl-2-hexanoïque[37]
- 2-éthylhexanoïque[37]
- n-éthylheptanooïque[37]
- 2-méthylheptanoïque[37]
- (E)-3-méthyl-2-hexanoïque[37]
- n-octanoïque[37]
- 2-méthyloctanoïque[37]
- 4-éthylheptanoïque[37]
- n-nonanoïque[37]
- 2-méthylnonanoïque[37]
- 4-éthyloctanoïque[37]
- n-décanoïque[37]
- 2-méthyldecanoïque[37]
- 4-éthylnonanoïque[37]
- 9-décénoïque[37]
- n-undécanoïque[37]
- 4-éthyldécanoïque[37]
Des carbonyls
Des alcools
Des hormones stéroïdes
Rôle des populations bactériennes dans la signature odorante du corps
- Les espèces du genre Corynebacteriumsont responsables de la modification des sécrétions apocrines (initialement inodores, incluant l’androstérone sulfate et la deshydroépiandostérone) en composés odorants (5-androst-16-en-3-one et acides gras à chaine courte).
- La bactérie Brevibacterium epidermidis contribue à l'odeur de pieds, plus forte chez certains individus en raison d'un taux élevé de méthanethiol et d’acide isovalérique. En laboratoire, ou à l'extérieur, le moustique Anopheles gambiae se montre très attiré par une odeur proche, produite par une bactérie parente Brevibacterium linens[44]. Meijerink et Van Loon (1999) ont montré[45] que l'odeur du fromage « Limburger », due à B. linens (et proche de l'odeur de pieds), attire les moustiques (2 à 3 fois plus de captures dans les pièges olfactifs en présence de fractions acides de ce fromage), de même pour des mélanges artificiels d’acides gras à chaine courte produits sur ces fromages, et pour les vraies odeurs de pieds[46],[47].
- La bactérie Bacillus cereus, naturellement présente sur la peau humaine produit des composés qui attirent le moustique Aedes aegypti[48].
Enjeux sanitaires et épidémiologiques
Les dermatoses sont presque toutes des pathologies du biofilm cutané. Elles sont difficiles à traiter, et souvent antibiorésistantes voire nosocomiales. Cette résistance accrue est acquise par plusieurs moyens[49]. Mieux comprendre la formation des biofilms pathogènes ainsi que la manière dont les bactéries y entretiennent des interactions synergiques[50] utiles ou pathogènes (et permettant éventuellement l'apparition d'antibiorésistance)[50] est donc un enjeu de santé publique.
Une peau saine est recouverte de biofilms adaptés aux différentes parties du corps. Ces biofilms naturels sont plus stables, résistants et protecteurs contre d'autres bactéries quand ils abritent une diversité d'espèces et de groupes de micro-organismes[50].
Inversement, la peau malade est généralement uniformément colonisée par une ou quelques espèces qui ont accidentellement trouvé des conditions leur permettant de proliférer anormalement et parfois d'empêcher toute cicatrisation[51]. Par exemple, un biofilm monospécifique de Staphylococcus aureus (qui peut atteindre son extension maximale en 3 jours) cause une dermatite atopique sèche difficile à soigner[52] et un biofilm monospécifique de Propionibacterium acnes est source d'acné[52],[53].
Un biofilm pathogène hétérogène (plurispécifique, c'est-à-dire formé par plusieurs pathogènes qui sont généralement des bactéries et/ou des champignons) est plus résistant (entre autres aux antibiotiques) qu'une biofilm pathogène monospécifique[50]. Cette résistance provient généralement d'intéractions entre polymères sécrétés par les microorganismes et/ou d'autres modifications de la matrice extracellulaire du biofilm qui devient plus dense, visqueuse voire quasi-imperméable[54],[50].
Les molécules (dont CO2 et vapeur d'eau) et divers composés émis par la peau et ses biofilms dans l'air (ou l'eau) sont des kéromones qui attirent de nombreux parasites hématophages (poux, puces, tiques, moustiques, mouches, punaises, taon, etc.), toutes espèces pouvant véhiculer et inoculer des parasites ou microbes pathogènes. À titre d'exemple :
- Le moustique Aedes aegypti (vecteur du paludisme) peut "remonter" une piste odorante via un courant d’air enrichi en composés émis par Bacillus cereus ;
- Plusieurs insectes hématophages testés en laboratoire (moustiques, mouches des sables) sont dotées de sensilles chémoréceptrices répondant à des stimuli tels que des acides carboxyliques, alcools, cétone et d'autres composés organiques produits par la peau humaine[réf. nécessaire], et le moustique Anopheles gambiae répond aux stimulations par des acides gras à chaine courte[réf. nécessaire] qui sont émis par la peau de l'Homme (comme par celle de canidés tels que chiens, renards, coyotes…[55],[56],[39],[57];
- les mouches hématophages Glossina morsitans[58] et Stomoxys calcitrans ou encore de la mouche tsétsé[58] se dirigent toutes vers une source (humaine ou autre) émettrice d'acétone (molécule émise par la digestion et rejeté à l'expiration ou par la peau[59]);
- plusieurs diptères hématophages (en laboratoire comme dans la nature) sont également attirés par certaines molécules libérées par l'urine de bovins[60],[61],[62],[63],[64]. C'est notamment le cas de L'ammoniac et l’urée qui se dégagent aussi de l'urine humaine (et moindrement de la peau et des poumons via l'expiration, à l'état de traces).
L’urée de la peau est rapidement décomposée en ammonium par la microflore[44].
L'ammoniac est une kairomone connue, repérée par les moustiques qu'elle attire[65],[66]. - Beaucoup d'invertébrés piqueurs et suceurs de sang détectent le CO2 émis par les animaux et l'Homme et peuvent remonter sa trace vers la source, notre haleine et expiration nasale qui en contiennent environ 45 000 ppm, alors que l'air n'en contient que 300 à 400 ppm.
Les cellules de la peau et les bactéries associées produisent aussi du CO2 mais à l'état de traces (0,25 % de ce qui est expiré par les poumons selon Frame et al., 1972). Ce CO2 est très attractif (et activateur du comportement de quête d'un hôte pour presque tous les hématophages. Les moustiques Ae. aegypti s'orientent par exemple vers une telle source de CO2 dès 500 ppm[67]) et volent plus activement que la teneur de l'air en CO2 est élevée. À partir de 800 ppm de CO2 [68], son activité de vol est positivement corrélée avec la teneur de l'air en CO2. De même pour la mouche S. calcitrans[69],[70]. Les récepteurs antennaires de Glossina palpalis réagissent dès 300 ppm[71] et ceux d'Ae. aegypti dès 100 ppm[72],[73] même si chez les moustiques, à grande distance, ce sont d'abord les odeurs corporelles qui sont perçues (ce qui peut être expliqué par une dilution plus rapide du CO2 dans les courants d’air[74],[75]. Il a été montré que la combinaison du CO2 à d'autres facteurs d'attraction déclenche ou améliore beaucoup l'orientation des insectes vers la source émettrice[76],[77],[78],[79]. - les tiques Amblyomma variegatum (qui sont des acariens et non des insectes) se montrent sensibles à des traces de CO2 dans l'air[80], mais aussi d'H2S (hydrogène sulfuré) ou d'aldéhydes (hexanal, nonanal, benzaldéhyde…) émises par l'haleine humaine ou d'autres vertébrés[81]. Ces composés sont identifiés par un organe dit Organe de Haller, situé sur le 1er tarse de la tique[82].
Interactions microflore-cosmétiques
Les parfums, produits de beauté et de soins parfumés et a fortiori certains répulsifs ou autres produits involontairement ou volontairement mis en contact avec la peau (lors d'activité professionnelle, de contacts avec des animaux ou certains matériaux...) peuvent interférer avec les odeurs de la peau et le biofilm, de même que les solvants organiques et détergents qui peuvent endommager ou provisoirement détruire le film hydrolipidique cutané[9].
Trop ou pas assez de lipides sur la couche cornée est signe de déséquilibre. Un manque de lipides entraîne une déshydratation de la peau (accroissement de la « perte insensible en eau ») qui indique que la fonction de barrière cutanée est dégradée[34].
Modalités d'étude
La microscopie a longtemps été la principale voie d'observation du microbiote cutané. Les progrès des techniques biomoléculaires ont ouvert de nouveaux champs d'étude via la génomique, la génétique moléculaire, la métagénomique[83],[84], le séquençage haut débit[85]et la « culturomique »[86].
Notes et références
- Sansonetti P (2011) La relation hôte-microbiote: une insondable symbiose ?. La lettre du Collège de France, (32), 14-15.
- Hacard, F., Nosbaum, A., Huynh, V. A., Nicolas, J. F., & Bérard, F. (2015). Plus il y a de bactéries différentes, moins il y a d’inflammation: la révolution microbiotique. In Annales de Dermatologie et de Vénéréologie (Vol. 142, pp. S13-S17, janvier 2015). Elsevier Masson (résumé)
- Michon A.L & Marchandin H (2015) Diversité physiopathologique du microbiote respiratoire. Revue Francophone des Laboratoires, 2015(469), 37-49
- (en) Kenneth Todar, « The Normal Bacterial Flora of Humans », sur Todar's Online Textbook of Bacteriology (consulté le 5 octobre 2013)
- Docteur Françoise Rodhain, La peau, la beauté et le temps, Cherche Midi, , p. 17.
- Evaldson G, Heimdahl A, Kager L, Nord CE (1982), The normal human anaerobic microflora. Scand J Infect Dis Suppl. 1982; 35:9-15.
- (en) Goller CC, Romeo T, « Environmental influences on biofilm development » Curr. Top. Microbiol. Immunol. 2008;322: 37- 66]
- Alice de Chalvet de Rochemonteix, Les biofilms et la peau, thèse vétérinaire, École vétérinaire de Maison-Alfort, octobre 2009, sous la direction de Geneviève Marignac (résumé (SUDOC))
- Marie-Claude Martini, Introduction à la dermopharmacie et à la cosmétologie, Lavoisier, , 3e éd., 500 p. (ISBN 9782743012700, lire en ligne)
- Nielsen ML, Raahave D, Stage JG, Justesen T (1975) Anaerobic and aerobic skin bacteria before and after skin-disinfection with chlorhexidine: An experimental study in volunteers. J. Clin. Path. 28 : 793-797 (résumé)
- (en) Macfarlane S, Dillon JF, « Microbial biofilms in the human gastrointestinal tract » J. Appl. Microbiol. 2007;102(5):1187- 1196
- (en) Feingold DS, « Bacterial adherence, colonization, and pathogenicity » Arch. Dermatol. 1986;122:161-163 (résumé)
- (en) Jeffrey C. Pommerville, Fundamentals of Microbiology, Jones & Bartlett Publishers, , p. 427-428.
- (en) Hall-Stoodley L, Costerton JW, Stoodley P « Bacterial biofilms: from the natural environment to infectious diseases » Nat. Rev. Microbiol. 2004;2(2):95-108.
- (en) Bjarnsholt T, Kirketerp-Møller K, Jensen PØ et al. « Why chronic wounds will not heal: a novel hypothesis » Wound Rep. Reg. 2008;16(1):2-10. (résumé)
- (en) Otto M, « Staphylococcal biofilms » Curr. Top. Microbiol. Immunol. 2008;322:207-28.
- (en) Elder MJ, Stapleton F, Evans E, Dart JK, « Biofilm-related infections in ophthalmology » Eye (Lond). 1995; 9 (Pt 1):102-9. (Résumé)
- (en) Zegans ME, Becker HI, Budzik J & O’Toole G, « The role of bacterial biofilms inocular infections » DNA and Cell Biology 2002;21:415-420 (résumé)
- (en) Parsek MR, Fuqua C, « Biofilms 2003: emerging themes and challenges in studies of surface- associated microbial life » J. Bacteriol. 2004;186:4427-40.
- Theodor Rosebury. Life on Man: Secker & Warburg, 1969 (ISBN 0-670-42793-4)
- Chiller K, Selkin BA, Murakawa GJ (2001) Skin microflora and bacterial infections of the skin. J. Investig. Dermatol. Symp. Proc. 6(3): 170-174
- (en) Cogen AL, Nizet V, Gallo RL, « Skin microbiota: a source of disease or defence ? », British Journal of Dermatology, vol. 158, no 3, , p. 442-455 (DOI 10.1111/j.1365-2133.2008.08437.x)
- Iwatsuki K, Yamasaki O, Morizane S, Oono T (2006) Staphylococcal cutaneous infections : Invasion, evasion and aggression. J. Dermatol. Sci. 42: 203-214 (résumé)
- Hentges DJ (1993) The anareobic microflora of the human body. Clin. Infect. Dis. 16 : 175- 180 (Résumé)
- (en) CE Desch « Human hair follicle mites and forensic acarology » Experimental and Applied Acarology. 2009;49(1-2):143-146. DOI:10.1007/s10493-009-9272-0 (résumé)
- Eleanor E. Sahn & Diane M. Sheridan, « Demodicidosis in a child with leukemia » Journal of the American Academy of Dermatology. 1992;27(5-Part. 2):799–801 (résumé)
- (en) E. Bonnar, P. Eustace, F.C. Powell, « The Demodex mite population in rosacea » Journal of the American Academy of Dermatology 1993;28(3):443–448. (résumé)
- (en) JL Diaz-Perez, « Demodex mites in rosacea » Journal of the American Academy of Dermatology 1994;30(5, Part 1):812–813. DOI:10.1016/S0190-9622(08)81529-0
- (en) Ya E Zhao, Na Guo and Li Ping Wu, « The effect of temperature on the viability of Demodex folliculorum and Demodex brevis » Parasitology Research. 2009;105(6):1623-1628.
- (en) Francesco Gioia et Leonardo Celleno (2002) « The dynamics of TEWL from hydrated skin » Skin Research and Technology 2002, vol. 8 p. 178-186
- (en) L Baumann, Basic Science of the epidermis. Cosmetic dermatology. Principles and Practice, Mc Graw-Hill. Medical Division Publishing, 2002, p. 3-8
- Maury E, Julié S, Charvéron M, Gall Y, Chap H, « Lipides et inflammation cutanée : place des phospholipases A2. Colloque sur les lipides de la peau » Pathologie Biologie 2003;51:248-252 (résumé)
- (en) Aly R, Maibach HI, Shinefield HR, Strauss WG, « Survival of pathogenic microorganisms on human skin » J. Investig. Dermatol. 1972;58:205-210
- Philippe Humbert, « Conséquences fonctionnelles des perturbations des lipides cutanés : Colloque sur les lipides de la peau », Pathologie Biologie, vol. 51, no 5, , p. 271-274. (résumé)
- (en) Matousek JL, Campbell KL, « A comparative review of cutaneous pH » Vet. Dermatol. 2002;13(6):293-300. (résumé)
- (en) Dustin J Penn, Elisabeth Oberzaucher, Karl Grammer, Gottfried Fischer, Helena A Soini, Donald Wiesler, Milos V Novotny, Sarah J Dixon, Yun Xu et Richard G Brereton, « Individual and gender fingerprints in human body odour » J. R. Soc. Interface 2007;4(13):331-340. DOI:10.1098/rsif.2006.0182 (Résumé)
- (en) X. Zeng, L. Leyden, H. Lawley, K. Sawano, I. Nohara, G. Preti, « Analysis of characteristic odors from human male axillae » J. Chem. Ecol. 1991;17:1469.
- (en) Braks MAH, Anderson RA et Knols BGJ. « Infochemicals in mosquito host selection human skin microflora and Plasmodium parasites » Parasitology Today 1999;15:409-413.
- (en) Bernier UR, Kline DL, Barnard DR, Schreck CE et Yost RA. « Analysis of human skin emanations by gas chromatography/mass spectrometry. 2. Identification of volatile compounds that are candidate attractants for the yellow fever mosquito (Aedes aegypti) » Analytical Chemistry 2000;72:747-756.
- Nicolaides N (1974)« Skin lipids: their biochemical uniqueness » Science 1974;186:19–26. DOI:10.1126/science.186.4158.19 (ISSN 0036-8075) (print), 1095-9203 (online) (résumé)
- (en) Acree F Jr. Turner RB, Gouck HK, Beroza M et Smith N. « L-Lactic acid: a mosquito attractant isolated from humans » Science 1968;161:1346-1347.
- (en) A Cork et Park KC, « Identification of electrophysiologically-active compounds for the malaria mosquito, Anopheles gambiae, in human sweat extracts » Medical and Veterinary Entomology 1996;10:269-276.
- (en) M Enserink, « What Mosquitoes Want: Secrets of Host Attraction » Science 2002;298:90-92.
- Braks, M.A.H., Anderson, R.A. et Knols, B.G.J. (1999). Infochemicals in mosquito host selection human skin microflora and Plasmodium parasites. Parasitology Today, 15: 409-413.
- Meijerink, J. et Van Loon, J.J.A. (1999) Sensitivities of antennal olfactory neurons of the malaria mosquito, Anopheles gambiae, to carboxylic acids. Journal of Insect Physiology, 45: 365-373.
- De Jong, R. et Knols, B.G.J. (1995). Olfactory responses of host-seeking Anopheles gambiae s.s. Giles (Diptera: Culicidae). Acta Tropica, 59: 333-335
- Knols, B.G.J., Van Loon, J.J.A., Cork, A., Robinson, R.D., Adam, W., Meijerink, J., De Jong, R. et Takken, W. (1997). Behavioural and electrophysiological responses of the female malaria mosquito Anopheles gambiae s. s. Giles (Diptera: Culicidae) towards Limburger cheese volatiles. Bulletin of Entomological Research, 87: 151-159
- Schreck, C.E. et James, J. (1968), Broth cultures of bacteria that attract female mosquitoes ; Mosquito News, 28: 33-38
- Mah TL, O’Toole GA (2001) Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 9: 34- 39]
- Burmolle M, Webb JS & al (2006) Enhanced biofilm formation and increased resistance to antimicrobial agents and bacterial invasion are caused by synergistic interactions in multispecies biofilms. Applied and environmental Microbiology, 3916-3923
- Costerton JW, Stewart PS & al. (1999) Bacterial biofilms: a common cause of persistent infections. Science, 284: 1318-1322
- Alice de Chalvet de Rochemonteix, Les biofilms et la peau, thèse vétérinaire, École vétérinaire de Maison-Alfort, octobre 2009. Voir p 114 et 122
- Burkhart (2003) Microbiology’s principle of biofilms as a major factor in the pathogenesis of acne vulgaris. International Journal of Dermatology. 42: 925-927
- Skillman LC, Sutherland IW, Jones MV (1999) The role of exopolysaccharides in dual species biofilm development. J.Appl.Microbiol. Symp. Suppl. 85:13S-18S
- Schreck, C.E. et James, J. (1968). Broth cultures of bacteria that attract female mosquitoes. Mosquito News, 28: 33-38
- Dougherty, M.J., Guerin, P.M., Ward, R.D. et Hamilton, J.G.C. (1999). Behavioural and electrophysiological responses of the phlebotomine sandfly Lutzomyia longipalpos (Diptera: Psychodidae) when exposed to canid host odour kairomones. Physiological Entomology, 24: 252-262.
- Meijerink, J., Braks, M.A.H., Brack, A., Adam, W. et Dekker, T. (2000). Identification of olfactory stimulants for Anopheles gambiae from human sweat samples. Journal of Chemical Ecology, 26: 1367-1382
- Bursell, E. (1984). Effects of host odour on the behaviour of tse-tse. Insect Science and Its Application, 5: 345-349
- Krotoszynski, B., Gabriel, G. et O'Neil, H. (1977). Characterization of human expired air: a promising investigative and diagnostic technique. Journal of Chromatographic Science, 15: 239-244
- Hassanali, A., Mc Dowell, P.G., Owaga, M.L.A. et Saini, R.K. (1986). Identification of tsetse attractants from excretory products of a wild host, Syncerus caffer. Insect Science and Its Application, 7: 5-9.
- Saini, R.K. (1986). Antennal responses of Glossina morsitans morsitans to buffalo urine, a potent olfactory attractant of tse-tse. Insect Science and Its Application, 7: 771-775
- Bursell, E. (1984). Effects of host odour on the behaviour of tse-tse. Insect Science and Its Application, 5: 345-349.
- Park, K.C. et Cork, A. (1999). Electrophysiological responses of antennal receptor neurons in female Australian sheep blowflies, Lucilia cuprina, to host odours. Journal of Insect Physiology, 45: 85-91.
- Van der Broek, I.V.F. et Den Otter, C.J. (1999). Olfactory sensitivities of mosquitoes with different host preferences (Anopheles gambiae s.s., An. arabiensis, An. quadriannulatus, An. m. atroparvus) to synthetic host odours. Journal of Insect Physiology, 45: 1001-1010.
- Geier, M., Bosch, O.J. et Boeckh, J. (1999). Ammonia as an attractive component of host odour for the yellow fever mosquito, Aedes aegypti. Chemical Senses, 24: 647-653
- Braks, M.A.H., Meijerink, J. et Takken, W. (2001). The response of the malaria mosquito, Anopheles gambiae, to two components of human sweat, ammonia and l-lactic acid, in an olfactometer. Physiological Entomology, 26: 142-148
- Geier, M., Bosch, O.J. et Boeck, J. (1999a). Influence of odour plume structure on upwind flight of mosquitoes towards hosts. Journal of Experimetal Biology, 202: 1639-1648
- Eiras, A.E. et Jepson, P.C. (1991). Host location by Aedes aegypti (Diptera: Culicidae): a wind tunnel study of chemical cues. Bulletin of Entomological Research, 81: 151-160
- Warnes, M.L. et Finlayson, L.H. (1986). Electroantennogram responses of the stable fly, Stomoxys calcitrans, to carbon dioxide and other host odours. Physiological Entomology, 11: 469-473
- Schofield, S., Witty, C. et Brady, J. (1997). Effects of carbon dioxide, acetone and 1-octen-3- ol on the activity of the stable fly, Stomoxys calcitrans. Physiological Entomology, 22: 256-260.
- Bogner, F. (1992). Response properties of CO2-sensitive receptors in tse-tse flies (Diptera: Glossina palpalis). Physiological Entomology, 17: 19-24
- Kellogg, F.E. (1970). Water vapour and carbon dioxide receptors in Aedes aegypti. Journal of Insect Physiology, 16: 99-108.
- Gillies, M.T. et Wilkes, T.J. (1972). The range of attraction of animal baits and carbon dioxide for mosquitoes. Studies in a freshwater area of West Africa. Bulletin of Entomological Research, 61: 389-404
- Warnes, M.L. et Finlayson, L.H. (1985). Responses of the stable fly, Stomoxys calcitrans (L.) (Diptera: Muscidae), to carbon dioxide and host odours. I. Activation. Bulletin of Entomological Research, 75: 519-527.
- Barrozo, R.B. et Lazzari, C.R. (2004a). The response of the blood-sucking bug Triatoma infestans to carbon dioxide and other host odours. Chemical Senses, 29: 319-329
- Gillies, M.T. (1980). The role of carbon dioxide in host-finding by mosquitoes (Diptera: Culicidae): a review. Bulletin of Entomological Research, 70: 525-532
- Eiras, A.E. et Jepson, P.C. (1991). Host location by Aedes aegypti (Diptera: Culicidae): a wind tunnel study of chemical cues. Bulletin of Entomological Research, 81: 151-160
- Steib, B.M., Geier, M. et Boeckh, J. (2001). The effect of lactic acid on odour-related host preference of yellow fever mosquitoes. Chemical Senses, 26: 523-528
- Dekker, T., Steib, B., Carde, R.T. et Geier, M. (2002). L-lactic acid: a human-signifying host cue for the anthropophilic mosquito Anopheles gambiae. Medical and Veterinary Entomology, 16: 91-98
- Steullet, P. et Guerin, P.M. (1992). Perception of breath components by the tropical bont tick, Amblyomma variegatum Fabricius (Ixodidae). I. CO2-excited and CO2-inhibited receptors. Journal of Comparative Physiology A, 170: 665-676
- Steullet, P. et Guerin, P.M. (1992). Perception of breath components by the tropical bont tick, Amblyomma variegatum Fabricius (Ixodidae). II. Sulfide-receptors. Journal of Comparative Physiology A, 170: 677-685
- Steullet, P. et Guerin, P.M. (1994). Identification of vertebrate volatiles stimulating olfactory receptors on tarsus I of the tick Amblyomma variegatum Fabricius (Ixodidae). II. Receptors outside the Haller's organ capsule. Journal of Comparative Physiology A, 174: 39-47
- Mathieu A (2014) Etude fonctionnelle de la communauté microbienne de la peau par une approche métagénomique (Doctoral dissertation, Ecully, Ecole centrale de Lyon).
- Guet-Revillet H (2014) Métagénomique bactérienne de l'hidrosadénite suppurée (Doctoral dissertation, Paris 5).
- Foulongne V, Sauvage V, Hebert C, Dereure O & Eloit M (2013) Virome cutané: étude systématique par séquençage haut débit. In Annales de Dermatologie et de Vénéréologie (Vol. 140, No. 12S1, pp. 581-582), décembre 2013 (résumé).
- La Scola, B. (2015). Nouvelle technique d’étude du microbiote: la culturomique. Revue Francophone des Laboratoires, 2015(469), 83-87
Voir aussi
Articles connexes
- Système digestif
- Peau
- Dermatologie
- Derme
- Épiderme
- Biofilm
- Ultra-violet
- Cosmétique
- Micrococcus
- Culturomique
Lien externe
Bibliographie
- Dunyach-Remy C, Sotto A & Lavigne J.P (2015) Le microbiote cutané: étude de la diversité microbienne et de son rôle dans la pathogénicité. Revue Francophone des Laboratoires, 2015(469), 51-58.
- Foulongne V, Sauvage V, Hebert C, Dereure O & Eloit M (2013) Virome cutané: étude systématique par séquençage haut débit. In Annales de Dermatologie et de Vénéréologie (Vol. 140, No. 12S1, pp. 581-582), décembre 2013 (résumé).
- Hillion M (2013). Interactions peau/microbiote cutané: étude du microbiote cutané cultivable et influence de produits cosmétiques sur la virulence bactérienne. Apports de la technique de spectrométrie de masse MALDI-TOF (Doctoral dissertation, Rouen) (résumé).
- Laborel-Préneron E (2015). Effets des sécrétomes de Staphylococcus aureus et Staphylococcus epidermidis du microbiote cutané d'enfants atopiques sur la réponse immunitaire T CD4 (Doctoral dissertation, Université de Toulouse, Université Toulouse III-Paul Sabatier).
- Lacour J.P (2015, January). Microbiote cutané et dermatite atopique: vers une nouvelle prise en charge thérapeutique?. In Annales de Dermatologie et de Vénéréologie (Vol. 142, pp. S18-S22). Elsevier Masson (résumé).
- Mijouin L (2012). Interactions peau/microbiote cutané: effet d'un neuropeptide cutané, la Substance P, sur l'adaptation et la virulence bactériennes (Doctoral dissertation, Rouen) (résumé).
- Mathieu, A. (2014). Étude fonctionnelle de la communauté microbienne de la peau par une approche métagénomique (Doctoral dissertation, Ecully, Ecole centrale de Lyon)
- Mokni M & Abdelhak S (2014). Flore cutanée, microbiote et microbiome.
- Romano-Bertrand S, Licznar-Fajardo P, Parer S & Jumas-Bilak E (2015) Impact de l’environnement sur les microbiotes: focus sur l’hospitalisation et les microbiotes cutanés et chirurgicaux. Revue Francophone des Laboratoires, 2015(469), 75-82.
- Sansonetti P (2011) La relation hôte-microbiote: une insondable symbiose ?. La lettre du Collège de France, (32), 14-15.
- Sansonetti P (2013) Des microbes et des hommes. Guerre et paix aux surfaces muqueuses (Leçon inaugurale prononcée le jeudi 20 novembre 2008). Collège de France.

