Chaîne respiratoire
En biochimie, on appelle chaîne respiratoire une chaîne de transport d'électrons réalisant l'oxydation des coenzymes réduites issues de la dégradation de composés organiques ou minéraux. Ces coenzymes sont notamment le NADH et le Q10H2 produits dans les mitochondries par le cycle de Krebs et par la β-oxydation des acides gras. Les chaînes respiratoires sont formées d'enzymes membranaires et de transporteurs d'électrons — cytochromes, quinones, flavoprotéines — organisés autour de la membrane plasmique chez les procaryotes et des crêtes mitochondriales de la membrane mitochondriale interne chez les eucaryotes.
• Dans ce processus, la chaîne respiratoire est une chaîne de transport d'électrons qui assure l'oxydation des coenzymes réduites, telles que le NADH, issues du catabolisme, par exemple du cycle de Krebs.
• Cette oxydation fournit l'énergie nécessaire au pompage de protons hors de la matrice mitochondriale vers l'espace intermembranaire mitochondrial, ce qui génère un gradient de concentration de protons et un gradient électrochimique à travers la membrane mitochondriale interne.
• Le reflux des protons vers la matrice à travers la membrane interne fournit l'énergie chimiosmotique nécessaire aux ATP synthases pour phosphoryler l'ADP en ATP.
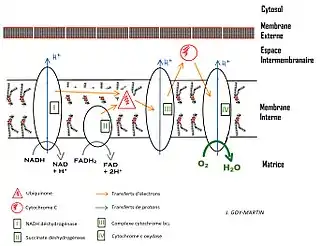
La chaîne respiratoire des eucaryotes a été intensivement étudiée. Elle est constituée essentiellement de quatre complexes protéiques dénombrés de I à IV. Les électrons à haut potentiel de transfert du NADH y entrent au niveau de la NADH déshydrogénase, ou complexe I, pour redonner du NAD+, tandis que ceux du succinate y entrent au niveau de la succinate déshydrogénase, ou complexe II, pour donner du fumarate. Un troisième point d'entrée existe au niveau de l'ETF déshydrogénase pour les électrons des electron-transferring flavoproteins (ETF). Dans ces trois cas, les électrons proviennent de la matrice mitochondriale et sont transférés à la coenzyme Q10 pour former de l'ubiquinol Q10H2, liposoluble, qui diffuse dans la bicouche lipidique constituant la membrane mitochondriale interne. Puis la coenzyme Q-cytochrome c réductase, ou complexe III, transfère les électrons du Q10H2 au cytochrome c dans l'espace intermembranaire, et la cytochrome c oxydase, ou complexe IV, transfère les électrons de quatre cytochromes c réduits à une molécule d'oxygène O2 pour former deux molécules d'eau H2O.
La circulation des électrons le long d'une chaîne respiratoire génère un gradient de concentration de protons à travers la membrane biologique où elle a lieu. Chez les eucaryotes, des protons sont pompés au niveau des complexes I, III et IV. Le gradient électrochimique qui en résulte emmagasine l'énergie libérée par les oxydations de la chaîne respiratoire et permet par la suite de phosphoryler l'ADP en ATP par couplage chimiosmotique avec des ATP synthases dans le cadre d'un processus global appelé phosphorylation oxydative.
Molécules de transfert d'électrons et de protons
La chaîne respiratoire assure le transport à la fois des protons et des électrons. Ces derniers circulent de donneurs en accepteurs d'électrons, tandis que les protons circulent à travers la membrane. Ces processus utilisent des molécules de transfert dont certaines sont solubles tandis que d'autres sont liées à des protéines. Dans les mitochondries, les électrons sont transférés dans l'espace intermembranaire mitochondrial par le cytochrome c, une petite protéine soluble dans l'eau assurant le transfert d'un électron par molécule[1]. Seuls les électrons sont transportés par ce cytochrome, et les transferts d'électrons de et vers cette protéine font intervenir l'oxydation et la réduction d'un atome de fer situé dans un groupe héminique lié au cytochrome. Certaines bactéries contiennent également le cytochrome c, chez lesquelles il est situé dans l'espace périplasmique[2].
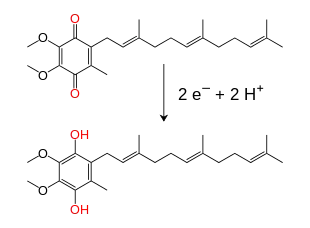
Dans la membrane mitochondriale interne, les électrons sont transportés par des ubiquinones, dont la forme principale chez les mammifères est la coenzyme Q10. Celle-ci est un transporteur d'électrons et de protons liposoluble faisant intervenir un cycle rédox[3]. Cette petite benzoquinone est très hydrophobe, ce qui lui permet de diffuser à l'intérieur de la membrane. Lorsque la coenzyme Q10 accepte deux électrons et deux protons, elle donne un ubiquinol Q10H2, qui en est la forme réduite ; ce dernier est susceptible de libérer deux électrons et deux protons pour redonner la coenzyme Q10, qui en est la forme oxydée. Par conséquent, si deux enzymes sont disposées de telle sorte que la coenzyme Q10 soit réduite d'un côté de la membrane et que l'ubiquinol Q10H2 soit oxydé de l'autre, l'ubiquinone joue le rôle d'une navette transportant les protons à travers la membrane[4]. Certaines chaînes respiratoires bactériennes utilisent des quinones différentes, comme la ménaquinone, en plus de l'ubiquinone[5].
Au niveau des protéines, les électrons circulent à travers une série de cofacteurs : flavine[6], protéines fer-soufre[7] et cytochromes. Il existe plusieurs types de centres fer-soufre. Le plus simple de ceux présents dans les chaînes de transfert d'électrons est formé de deux atomes de fer liés à deux atomes de soufre ; ce sont les clusters [2Fe–2S]. L'autre variété de centre fer-soufre est appelée [4Fe–4S] et contient un cube formé de quatre atomes de fer et quatre atomes de soufre. Chaque atome de fer de tels clusters est coordonné à la chaîne latérale d'un résidu d'acide aminé, généralement à l'atome de soufre d'un résidu de cystéine. Les cofacteurs à ions métalliques subissent des réactions sans fixer ni libérer de protons, de sorte que, dans la chaîne de transport d'électrons, ils interviennent uniquement dans le transport des électrons à travers des protéines. Les électrons peuvent parcourir d'assez grandes distances en sautant de proche en proche le long de chaînes de tels cofacteurs[8]. Ce mécanisme fait intervenir l'effet tunnel, qui est efficace sur des distances inférieures à 1,4 nm[9].
Chaîne respiratoire mitochondriale
La plupart des cellules d'eucaryotes, possèdent des mitochondries, lesquelles produisent de l'ATP à partir des produits de voies métaboliques telles que le cycle de Krebs, la β-oxydation, ou encore la dégradation des acides aminés. Dans la membrane mitochondriale interne, les électrons issus du NADH et du succinate circulent à travers la chaîne respiratoire jusqu'à réduire une molécule d'oxygène O2, ce qui libère une molécule d'eau H2O. Chaque donneur d'électrons de la chaîne respiratoire transmet ses électrons à un accepteur plus électronégatif que lui selon une cascade de réactions qui se termine par la réduction de l'oxygène, le plus électronégatif des accepteurs d'électrons de la chaîne respiratoire.
Le transfert d'électrons entre un donneur et un accepteur plus électronégatif libère de l'énergie, utilisée pour pomper des protons hors de la matrice mitochondriale, ce qui génère un gradient de concentration de protons, et donc un gradient électrochimique, à travers la membrane mitochondriale interne. Ce gradient électrochimique est ensuite utilisé par l'ATP synthase pour produire de l'ATP : on dit que la chaîne respiratoire et la phosphorylation de l'ADP en ATP sont couplées par chimiosmose au moyen d'un gradient de concentration d'ions H+ à travers la membrane interne de la mitochondrie.
| Complexe | Enzyme respiratoire | Couple rédox | Potentiel standard |
|---|---|---|---|
| I | NADH déshydrogénase | NAD+ / NADH | −0,32 V |
| II | Succinate déshydrogénase | FMN ou FAD / FMNH2 ou FADH2 | −0,20 V |
| III | Cytochrome c réductase | Coenzyme Q10 ox / Q10H2 | 0,06 V |
| Cytochrome b ox / Cytochrome b réd | 0,12 V | ||
| IV | Cytochrome c oxydase | Cytochrome c ox / Cytochrome c réd | 0,22 V |
| Cytochrome a ox / Cytochrome a réd | 0,29 V | ||
| O2 / HO− | 0,82 V | ||
| Conditions : pH = 7 | |||
Complexe I : NADH déshydrogénase

La NADH déshydrogénase (EC ), également appelée complexe I, est la première enzyme de la respiration cellulaire[11]. Il s'agit d'une protéine de très grande taille qui, chez les mammifères, possède 46 sous-unités et une masse d'environ 1 000 kDa[12]. Le détail de sa structure a été déterminé à partir de Thermus thermophilus (en)[13]. Chez la plupart des organismes, elle a une forme rappelant grossièrement une botte[14],[15]. Les gènes qui encodent cette protéine se trouvent à la fois dans le noyau et dans le génome mitochondrial, comme c'est le cas pour de nombreuses enzymes mitochondriales.
Le complexe I catalyse la réduction de la coenzyme Q10, une ubiquinone liposoluble présente dans la membrane mitochondriale interne, par les deux électrons à haut potentiel de transfert du NADH pour donner de l'ubiquinol, ou Q10H2 :
- NADH + coenzyme Q10 + 5 H+matriciels → NAD+ + Q10H2 + 4 H+intermembranaires.
L'amorçage de la réaction, et en fait de toute la chaîne de transport d'électrons, a lieu avec la liaison d'une molécule de NADH au complexe I et la cession de deux électrons du premier à un groupe prosthétique FNM du second. Les électrons du FNMH2 circulent ensuite à travers une série de centres fer-soufre[13], à la fois de type [2Fe-2S] et [4Fe-4S].
Alors que les électrons circulent à travers le complexe I, quatre protons sont pompés hors de la matrice mitochondriale vers l'espace intermembranaire à travers la membrane interne. Le mécanisme exact de ce processus demeure obscur, mais il semble faire intervenir des changements conformationnels du complexe I conduisant à lier des protons du côté de la face N de la membrane pour les libérer du côté de la face P[16]. Les électrons sont finalement cédés par la chaîne de centres fer-soufre à une molécule d'ubiquinone (coenzyme Q10) dans la membrane[11]. La formation d'ubiquinol (Q10H2) contribue également à la génération du gradient de concentration de protons à travers la membrane interne dans la mesure où les deux protons nécessaires à cette réaction proviennent de la matrice mitochondriale.
Complexe II : succinate déshydrogénase

La succinate déshydrogénase (EC ), également appelée complexe II, est le deuxième point d'entrée des électrons dans la chaîne respiratoire[17]. Cette enzyme présente la particularité d'appartenir à la fois au cycle de Krebs et à la chaîne respiratoire. Elle est constituée de quatre sous-unités et de plusieurs cofacteurs : FAD, centres fer-soufre, et un groupe héminique qui ne participe pas au transfert des électrons à la coenzyme Q10 mais jouerait un rôle important en limitant la production de dérivés réactifs de l'oxygène[18],[19]. Elle catalyse l'oxydation du succinate en fumarate et la réduction de la coenzyme Q10. Cette réaction libère moins d'énergie que l'oxydation du NADH, ce qui fait que le complexe II ne contribue pas à la génération du gradient de concentration de protons.
- Succinate + coenzyme Q10 → fumarate + Q10H2.
Chez certains eucaryotes tels que le ver parasite Ascaris suum (en), une enzyme semblable au complexe II, la fumarate réductase à ménaquinone, catalyse la réaction inverse pour réduire le fumarate en oxydant l'ubiquinol, ce qui permet à ce parasite de survivre dans l'environnement anaérobie du gros intestin en réalisant une phosphorylation oxydative avec le fumarate comme accepteur d'électrons[20]. Le parasite du paludisme, Plasmodium falciparum, utilise quant à lui le complexe II dans le sens inverse pour régénérer l'ubiquinol dans le cadre de la biosynthèse de pyrimidines[21].
ETF déshydrogénase
L’ETF déshydrogénase (EC ) est un troisième point d'entrée des électrons dans la chaîne respiratoire. Il s'agit d'une enzyme capable d'utiliser des électrons de flavoprotéines — les electron-transferring flavoproteins (ETF) — de la matrice mitochondriale pour réduire la coenzyme Q10[22]. Cette enzyme contient une flavine et un cluster [4Fe-4S] mais, contrairement aux autres complexes respiratoires, elle est liée à la surface de la membrane mitochondriale interne et ne traverse pas la bicouche lipidique[23]. Le potentiel standard du couple rédox ETFoxydée/ETFréduite vaut –0,197 V chez Methylophilus methylotrophus[24], une bactérie méthylotrophe (en).
- ETFréduite + coenzyme Q10 → ETFoxydée + Q10H2.
Cette enzyme est importante chez les mammifères car elle permet aux électrons issus de l'oxydation des acides gras, des acides aminés ou encore de la choline, d'entrer dans la chaîne respiratoire à partir de nombreuses acétyl-CoA déshydrogénases[25],[26]. Elle joue également un rôle important chez les plantes dans les réponses métaboliques aux périodes d'obscurité prolongées[27].
Complexe III : coenzyme Q-cytochrome c réductase

La coenzyme Q-cytochrome c réductase (EC ) est également appelée complexe III, ou complexe cytochrome bc1[28],[29]. Chez les mammifères, il s'agit d'un dimère de deux complexes formés chacun de 11 sous-unités, un cluster [2Fe-2S] et trois cytochromes : un cytochrome c1 et deux cytochromes b[30]. Un cytochrome est une variété de protéines de transfert d'électrons contenant au moins un groupe héminique. L'atome de fer du complexe III oscille entre l'état d'oxydation +2 (ferreux) et l'état d'oxydation +3 (ferrique) lors de la circulation des électrons à travers la protéine.
Le complexe III catalyse l'oxydation d'une molécule d'ubiquinol Q10H2 et la réduction de deux molécules de cytochrome c. Contrairement à la coenzyme Q10, qui transporte deux électrons par molécule, le cytochrome c ne transporte qu'un électron par molécule.
- Q10H2 + 2 cytochrome coxydé + 2 H+matriciel → coenzyme Q10 + 2 cytochrome créduit + 4 H+intermembranaire.
Dans la mesure où un seul électron peut être transféré d'un Q10H2 donneur à un cytochrome c accepteur à chaque fois, le mécanisme réactionnel du complexe III est plus subtil que celui des autres complexes respiratoires et se déroule en deux étapes formant ce qu'on appelle le cycle Q[31] :
- À la première étape, l'enzyme se lie à trois substrats : d'abord Q10H2, qui est oxydé en coenzyme Q10 ; puis un cytochrome c, qui reçoit l'un des deux électrons de Q10H2 ; puis enfin une coenzyme Q10, qui reçoit le second électron pour former une semiquinone Q10• –, laquelle est un radical libre. Les deux protons issus de Q10H2 passent dans l'espace intermembranaire mitochondrial. Les deux premiers substrats sont libérés, tandis que la semiquinone demeure liée.
- À la seconde étape, une seconde molécule de Q10H2 est liée et cède un électron à un cytochrome c accepteur et l'autre électron à la semiquinone liée. Celle-ci redonne Q10H2 en absorbant deux protons, provenant cette fois de la matrice mitochondriale. Les deux protons issus de la seconde molécule de Q10H2 passent à leur tour dans l'espace intermembranaire mitochondrial, tandis que la molécule Q10H2 formée à partir de la semiquinone est libérée de l'enzyme[32].
Deux protons sont injectés dans l'espace intermembranaire à chacune des deux étapes du cycle Q, ce qui contribue à la génération d'un gradient de concentration de protons à travers la membrane mitochondriale interne. Ce mécanisme complexe en deux étapes permet ainsi d'injecter quatre protons dans l'espace intermembranaire par molécule de Q10H2 oxydée, alors que seulement deux protons le seraient si Q10H2 cédait directement ses deux électrons à deux cytochromes c[6].
Complexe IV : cytochrome c oxydase

La cytochrome c oxydase (EC ), également appelée complexe IV, est la protéine terminant la chaîne respiratoire mitochondriale[33]. Chez les mammifères, la structure de cette enzyme est extrêmement compliquée : elle est formée de 13 sous-unités, deux groupes héminiques et plusieurs cofacteurs renfermant des ions métalliques, plus précisément trois de cuivre, un de magnésium et un de zinc[34].
Cette enzyme catalyse le transfert d'électrons à une molécule d'oxygène O2 tout en pompant des protons à travers la membrane mitochondriale interne[35]. Deux molécules d'eau H2O sont formées par molécule d'oxygène réduite, tandis que quatre protons sont expulsés vers l'espace intermembranaire mitochondrial, ce qui contribue à la génération du gradient de concentration de protons à travers la membrane interne :
- 4 cytochrome créduit + O2 + 8 H+matriciel → 4 cytochrome coxydé + 2 H2O + 4 H+intermembranaire.
Réductases et oxydases alternatives
De nombreux eucaryotes ont une chaîne respiratoire différente de celle des mammifères décrite ci-dessus. Par exemple, les plantes ont des NADH oxydases qui oxydent le NADH dans le cytosol plutôt que dans la matrice mitochondriale en transférant les électrons correspondants à des ubiquinones[36]. Cependant, ces enzymes cytosoliques ne participent pas à la génération du gradient de concentration de protons à travers la membrane mitochondriale interne[37].
Un autre exemple d'enzyme respiratoire divergente est l'oxydase alternative (AOX), ou ubiquinol oxydase non électrogène, présente chez les plantes, les champignons, les protistes et peut-être également chez certains animaux[38],[39]. Cette enzyme transfère directement les électrons de l'ubiquinol à l'oxygène[40].
Les chaînes respiratoires impliquant ces oxydases alternatives présentent un plus faible rendement en ATP que la chaîne respiratoire complète des mammifères. Les avantages procurés par ces chaînes raccourcies ne sont pas complètement élucidés. Cependant, l'oxydase alternative est produite en réponse au stress tel que le froid, la présence de dérivés réactifs de l'oxygène et d'agents infectieux, ainsi qu'à d'autres facteurs qui inhibent la chaîne respiratoire complète[41],[42]. Les chaînes respiratoires alternatives pourraient ainsi contribuer à la résistance des organismes au stress oxydant[43].
Supercomplexes ou respirasomes
Les différents complexes de la chaîne respiratoire ont été initialement présentés comme diffusant librement et indépendamment les uns des autres dans la membrane mitochondriale interne[44]. Cependant, au fur et à mesure que les observations et les données s'accumulent, il apparaît de plus en plus en clair que les complexes respiratoires tendent à s'organiser dans les membranes en structures supramoléculaires de grande taille appelées supercomplexes ou « respirasomes »[45]. Toutes les études expérimentales ne corroborent cependant pas toujours l'observation de supercomplexes, de sorte que leur présence dans les membranes n'est pas systématique[12],[46].
Dans les respirasomes, les complexes s'organisent en ensembles d'enzymes en interaction[47]. Ces associations pourraient agir en canalisant les flux de substrats entre les complexes successifs, ce qui optimiserait l'efficacité des transferts d'électrons entre les différentes protéines qui les constituent[48]. Chez les mammifères, certains complexes seraient plus représentés que d'autres au sein de ces ensembles de supercomplexes, certaines données suggérant un ratio entre les cinq composantes de la phosphorylation oxydative I / II / III / IV / V (où V représente l'ATP synthase) d'environ 1:1:3:7:4[49].
Chaîne respiratoire bactérienne
Contrairement aux eucaryotes, dont les chaînes de transport d'électrons sont très semblables, les bactéries possèdent une grande variété d'enzymes de transfert d'électrons, qui utilisent également une grande variété de substrats[50]. Le transport d'électrons des procaryotes — bactéries et archées — partage cependant avec les eucaryotes le fait d'utiliser l'énergie libérée au cours de l'oxydation des substrats pour pomper des ions à travers une membrane et produire un gradient électrochimique à travers cette membrane. Chez les bactéries, la chaîne respiratoire d’Escherichia coli est la mieux comprise, tandis que celle des archées demeure mal connue[51].
La principale différence entre la chaîne respiratoire des procaryotes et celle des eucaryotes est que les bactéries et les archées utilisent une grande variété de substances comme donneurs et accepteurs d'électrons. Ceci permet aux procaryotes de croître dans des conditions environnementales très diverses[52]. Chez Escherichia coli, par exemple, la chaîne respiratoire peut être le fait d'un grand nombre de couples oxydant / réducteur, dont le tableau ci-dessous dresse une liste en indiquant leur potentiel standard ; ce dernier mesure l'énergie libérée lorsqu'une substance est oxydée ou réduite, les réducteurs ayant un potentiel négatif tandis que les oxydants ont un potentiel positif.
| Enzyme respiratoire | Couple rédox | Potentiel standard |
|---|---|---|
| Formiate déshydrogénase | HOCOO– / HCOO– | −0,43 V |
| Hydrogénase | H+ / H2 | −0,42 V |
| NADH déshydrogénase | NAD+ / NADH | −0,32 V |
| Glycérol-3-phosphate déshydrogénase | DHAP / Gly-3-P | −0,19 V |
| Pyruvate oxydase | Acétate + CO2 / Pyruvate | ? |
| Lactate déshydrogénase | Pyruvate / Lactate | −0,19 V |
| D-Aminoacide déshydrogénase | α-Cétoacide + NH3 / Acide aminé D | ? |
| Glucose déshydrogénase | Gluconate / Glucose | −0,14 V |
| Succinate déshydrogénase Fumarate réductase (sens inverse) |
Fumarate / Succinate | 0,03 V |
| Triméthylamine N-oxyde réductase | TMAO / TMA | 0,13 V |
| DMSO réductase | (CH3)2S=O / (CH3)2S | 0,16 V |
| Nitrite réductase | NO2– / NH3 | 0,36 V |
| Nitrate réductase | NO3– / NO2– | 0,42 V |
| Ubiquinol oxydase | O2 / H2O | 0,82 V |
Comme le montre ce tableau, E. coli peut croître en présence de réducteurs tels que le formiate, l'hydrogène ou le lactate comme donneurs d'électrons, et d'oxydants tels que le nitrate, le diméthylsulfoxyde (DMSO) ou l'oxygène comme accepteurs d'électrons[52]. Plus la différence entre le potentiel standard de l'oxydant et celui du réducteur est élevée, plus l'énergie libéré est élevée. Parmi ces composés, le couple rédox succinate / fumarate est inhabituel dans la mesure où son potentiel standard est proche de zéro. Le succinate peut ainsi être oxydé en fumarate en présence d'un oxydant fort comme l'oxygène, tandis que le fumarate peut être réduit en succinate en présence d'un réducteur fort comme le formiate. Ces réactions alternatives sont respectivement catalysées par la succinate déshydrogénase et la fumarate réductase[54].
Certains procaryotes utilisent des couples rédox qui ne présentent qu'une faible différence de potentiel standard. Ainsi, les bactéries nitrifiantes telles que Nitrobacter oxydent le nitrite en nitrate en cédant leurs électrons à l'oxygène. La faible quantité d'énergie libérée par cette réaction suffit pour pomper des protons et synthétiser de l'ATP, mais est insuffisante pour produire directement du NADH ou du NADPH à destination des biosynthèses (anabolisme)[55]. Ce problème est résolu à l'aide d'une nitrite réductase qui génère une force proton-motrice suffisante pour inverser une partie de la chaîne de transport d'électrons afin que le complexe I produise du NADH[56],[57].
Les procaryotes contrôlent l'utilisation de leurs donneurs et accepteurs d'électrons en ajustant la biosynthèse des enzymes en fonction des fluctuations des conditions environnementales[58]. Cette flexibilité est rendue possible par le fait que les différentes oxydases et réductases utilisent le même pool d'ubiquinones. Ceci permet à de nombreuses combinaisons d'enzymes de fonctionner ensemble en échangeant de l'ubiquinol[53]. Ces chaînes respiratoires présentent de ce fait une nature modulaire formée de groupes d'enzymes facilement interchangeables.
En plus de cette diversité métaboliques, les procaryotes possèdent également un ensemble d'isoenzymes , c'est-à-dire de protéines différentes qui catalysent la même réaction chimique. Il existe ainsi deux ubiquinol oxydases utilisant l'oxygène comme accepteur d'électrons : sous environnement fortement aérobie, la cellule utilise une oxydase ayant une faible affinité pour l'oxygène pouvant transporter deux protons par électron, tandis que, si le milieu tend à devenir anaérobie, la même cellule utilise une oxydase ne pouvant transporter qu'un seul proton par électron mais ayant une affinité supérieure pour l'oxygène[59].
Formes réactives de l'oxygène
L'oxygène moléculaire O2 est un oxydant fort, ce qui en fait un excellent accepteur final d'électrons. Cependant, la réduction de l'oxygène fait intervenir des intermédiaires potentiellement dangereux[60]. Même si le transfert de quatre protons et quatre électrons à une molécule d'O2 produit une molécule d'eau H2O, laquelle est sans danger et même bénéfique à toutes les formes de vies connues, le transfert d'un ou deux électrons produit respectivement l'ion superoxyde O2• – et l'ion peroxyde O22–, qui sont très réactifs et par conséquent dangereux pour la plupart des êtres vivants. Ces formes réactives de l'oxygène, ainsi que leurs dérivés tels que le radical hydroxyle HO•, sont très nocifs pour les cellules car ils oxydent les protéines et provoquent des mutations génétiques en altérant l'ADN. L'accumulation de telles dégradations dans les cellules pourrait contribuer à l'apparition de maladies et serait l'une des causes de la sénescence[61],[62] (vieillissement).
La cytochrome c oxydase catalyse très efficacement la réduction de l'oxygène en eau, et elle ne libère qu'une très faible quantité d'intermédiaires partiellement réduits. Cependant, de petites quantités d'ions superoxyde et peroxyde sont malgré tout produits par la chaîne de transport d'électrons[63]. La réduction de la coenzyme Q10 par le complexe III joue à cet égard un rôle important, car il se forme un radical libre semiquinone comme intermédiaire réactionnel. Cette espèce chimique instable est susceptible de laisser fuir des électrons directement sur l'oxygène pour former l'ion superoxyde O2• –[64]. Dans la mesure où la production de dérivés réactifs de l'oxygène par la chaîne respiratoire croît avec le potentiel de membrane, on pense que les mitochondries régulent leur activité pour maintenir leur potentiel de membrane dans une plage de valeurs étroite qui optimise la production d'ATP tout en minimisant la production de dérivés réactifs de l'oxygène[65]. Les oxydants pourraient ainsi activer des protéines de découplage qui réduiraient le potentiel de membrane[66].
Pour contrecarrer les effets néfastes de ces dérivés réactifs de l'oxygène, les cellules contiennent de nombreux systèmes antioxydants, parmi lesquels des vitamines antioxydantes telles que la vitamine C et la vitamine E, et des enzymes antioxydantes telles que la superoxyde dismutase, la catalase et les peroxydases[60].
Inhibiteurs
Plusieurs médicaments et toxines bien connus sont des inhibiteurs de la chaîne respiratoire. Bien que chacune de ces toxines n'inhibe qu'une seule enzyme de la chaîne de transport d'électrons, l'inhibition d'une étape quelconque de cette chaîne bloque l'ensemble du processus.
| Complexe | Inhibiteur | Utilisation | Mode d'action sur la chaîne respiratoire |
|---|---|---|---|
| I | Roténone | Pesticide | Bloque le transfert d'électrons du complexe I vers la coenzyme Q10 en bloquant le site de liaison à l'ubiquinone[67]. |
| II | Malonate Oxaloacétate SDHI |
Poisons | Ce sont des inhibiteurs compétitifs de la succinate déshydrogénase (complexe II)[68]. |
| III | Antimycine A | Piscicide | Elle se lie au site Qi de la coenzyme Q-cytochrome c réductase, ce qui bloque l'oxydation de l'ubiquinol. |
| IV | CN– CO N3– H2S |
Poisons | Inhibent la chaîne respiratoire en se liant plus fortement que l'oxygène au centre Fe–Cu de la cytochrome c oxydase, ce qui empêche la réduction de l'oxygène[69]. |
Dans la mesure où la chaîne respiratoire fonctionne de pair avec la phosphorylation de l'ADP en ATP par l'ATP synthase dans le cadre du processus global de phosphorylation oxydative, cette chaîne peut également être bloquée par l'arrêt du reflux des protons vers la matrice mitochondriale : c'est ce qui se produit lorsque l'ATP synthase est inhibée par l'oligomycine[70] ; les pompes à protons deviennent ainsi incapables d'expulser des protons hors de la matrice lorsque les électrons circulent à travers la chaîne respiratoire car le gradient de concentration en protons devient trop élevé à travers la membrane, de sorte qu'elles ne peuvent plus contrer le gradient électrochimique. Le NADH cesse d'être oxydé en NAD+, et le cycle de Krebs cesse à son tour de fonctionner faute d'une concentration suffisante en NAD+.
| Cible | Composé | Utilisation | Mode d'action indirect à travers le couplage chimiosmotique avec l'ATP synthase |
|---|---|---|---|
| ATP synthase | Oligomycine | Antibiotique | Inhibe l'ATP synthase en bloquant le flux de protons à travers la région FO[70], ce qui bloque en retour toutes les pompes à protons de la membrane (complexe I, complexe III, et complexe IV) en raison de l'accroissement excessif du gradient électrochimique. |
| Membrane interne | Carbonylcyanure m-chlorophénylhydrazone 2,4-Dinitrophénol |
Poisons, amaigrissement[71] | Il s'agit d'ionophores qui brisent le gradient de concentration en laissant les protons diffuser librement à la travers la membrane mitochondriale interne[72]. |
Tous les inhibiteurs de la phosphorylation oxydative ne sont cependant pas des toxines. Dans le tissu adipeux brun, des protéines découplantes permettent de réguler la diffusion des protons à travers la membrane mitochondriale interne, ce qui a pour effet de découpler la chaîne de transport d'électrons — dont le fonctionnement n'est pas altéré — de la production d'ATP[73]. Cette respiration découplée a pour effet de produire de la chaleur au lieu de l'ATP, ce qui joue un rôle vital dans la thermorégulation notamment des animaux en hibernation, bien que ces protéines puissent également avoir une fonction plus générale de réponse cellulaire au stress[74].
Notes et références
- (en) F. Scott Mathews, « The structure, function and evolution of cytochromes », Progress in Biophysics and Molecular Biology, vol. 45, no 1, , p. 1-56 (PMID 3881803, DOI 10.1016/0079-6107(85)90004-5, lire en ligne)
- (en) Paul M. Wood, « Why do c-type cytochromes exist? », FEBS Letters, vol. 164, no 2, , p. 223-226 (PMID 6317447, DOI 10.1016/0014-5793(83)80289-0, lire en ligne)
- (en) Frederick L. Crane, « Biochemical Functions of Coenzyme Q10 », Journal of the American College of Nutrition, vol. 20, no 6, , p. 591-598 (PMID 11771674, DOI 10.1080/07315724.2001.10719063, lire en ligne)
- (en) Peter Mitchell, « Keilin's respiratory chain concept and its chemiosmotic consequences », Science, vol. 206, no 4423, , p. 1148-1159 (PMID 388618, DOI 10.1126/science.388618, Bibcode 1979Sci...206.1148M, lire en ligne)
- (en) Britta Søballe et Robert K. Poole, « Microbial ubiquinones: multiple roles in respiration, gene regulation and oxidative stress management », Microbiology, vol. 145, no Pt 8, , p. 1817-1830 (PMID 10463148, DOI 10.1099/13500872-145-8-1817, lire en ligne)
- (en) Brian E. Schultz et Sunney I. Chan, « Structures and Proton-Pumping Strategies of Mitochondrial Respiratory Enzymes », Annual Review of Biophysics and Biomolecular Structure, vol. 30, , p. 23-65 (PMID 11340051, DOI 10.1146/annurev.biophys.30.1.23, lire en ligne)
- (en) Deborah C. Johnson, Dennis R. Dean, Archer D. Smith et Michael K. Johnson, « STRUCTURE, FUNCTION, AND FORMATION OF BIOLOGICAL IRON-SULFUR CLUSTERS », Annual Review of Biochemistry, vol. 74, , p. 247-281 (PMID 15952888, DOI 10.1146/annurev.biochem.74.082803.133518, lire en ligne)
- (en) David Leys et Nigel S Scrutton, « Electrical circuitry in biology: emerging principles from protein structure », Current Opinion in Structural Biology, vol. 14, no 6, , p. 642-647 (PMID 15582386, DOI 10.1016/j.sbi.2004.10.002, lire en ligne)
- (en) Christopher C. Page, Christopher C. Moser, Xiaoxi Chen et P. Leslie Dutton, « Natural engineering principles of electron tunnelling in biological oxidation–reduction », Nature, vol. 402, no 6757, , p. 47-52 (PMID 10573417, DOI 10.1038/46972, Bibcode 1999Natur.402...47P, lire en ligne)
- Anders Overgaard Pedersen et Henning Nielsen, « Medical CHEMISTRY Compendium », Aarhus University, 2008.
- (en) J. Hirst, « Energy transduction by respiratory complex I--an evaluation of current knowledge », Biochemical Society Transactions, vol. 33, no 3, , p. 525-529 (PMID 15916556, DOI http://www.biochemsoctrans.org/content/33/3/525, 10.1042/BST0330525)
- (en) Giorgio Lenaz, Romana Fato, Maria Luisa Genova, Christian Bergamini, Cristina Bianchi et Annalisa Biondi, « Mitochondrial Complex I: Structural and functional aspects », Biochimica et Biophysica Acta (BBA) - Bioenergetics, vol. 1757, nos 9-10, , p. 1406-1420 (PMID 16828051, DOI 10.1016/j.bbabio.2006.05.007, lire en ligne)
- (en) Leonid A. Sazanov et Philip Hinchliffe, « Structure of the Hydrophilic Domain of Respiratory Complex I from Thermus thermophilus », Science, vol. 311, no 5766, , p. 1430-1436 (PMID 16469879, DOI 10.1126/science.1123809, Bibcode 2006Sci...311.1430S, lire en ligne)
- (en) Ekaterina A. Baranova, Peter J. Holt et Leonid A. Sazanov, « Projection Structure of the Membrane Domain of Escherichia coli Respiratory Complex I at 8 Å Resolution », Journal of Molecular Biology, vol. 366, no 1, , p. 140-154 (PMID 17157874, DOI 10.1016/j.jmb.2006.11.026, lire en ligne)
- (en) Thorsten Friedrich et Bettina Böttcher, « The gross structure of the respiratory complex I: a Lego System », Biochimica et Biophysica Acta (BBA) - Bioenergetics, vol. 1608, no 1, , p. 1-9 (PMID 14741580, DOI 10.1016/j.bbabio.2003.10.002, lire en ligne)
- (en) Judy Hirst, « Towards the molecular mechanism of respiratory complex I », Biochemical Journal, vol. 425, no 2, , p. 327-339 (PMID 20025615, DOI 10.1042/BJ20091382, lire en ligne)
- (en) Gary Cecchini, « Function and Structure of Complex II of the Respiratory Chain », Annual Review of Biochemistry, vol. 72, , p. 77-109 (PMID 14527321, DOI 10.1146/annurev.biochem.72.121801.161700, lire en ligne)
- (en) Victoria Yankovskaya, Rob Horsefield, Susanna Törnroth, César Luna-Chavez, Hideto Miyoshi, Christophe Léger, Bernadette Byrne, Gary Cecchini et So Iwata, « Architecture of Succinate Dehydrogenase and Reactive Oxygen Species Generation », Science, vol. 299, no 5607, , p. 700-704 (PMID 12560550, DOI 10.1126/science.1079605, Bibcode 2003Sci...299..700Y, lire en ligne)
- (en) Rob Horsefield, So Iwata et Bernadette Byrne, « Complex II from a Structural Perspective », Current Protein & Peptide Science, vol. 5, no 2, , p. 107-118 (PMID 15078221, DOI 10.2174/1389203043486847, lire en ligne)
- (en) Kiyoshi Kita, Hiroko Hirawake, Hiroko Miyadera, Hisako Amino et Satoru Takeo, « Role of complex II in anaerobic respiration of the parasite mitochondria from Ascaris suum and Plasmodium falciparum », Biochimica et Biophysica Acta (BBA) - Bioenergetics, vol. 1553, nos 1-2, , p. 123-139 (PMID 11803022, DOI 10.1016/S0005-2728(01)00237-7, lire en ligne)
- (en) Heather J. Painter, Joanne M. Morrisey, Michael W. Mather et Akhil B. Vaidya, « Specific role of mitochondrial electron transport in blood-stage Plasmodium falciparum », Nature, vol. 446, no 7131, , p. 88-91 (PMID 17330044, DOI 10.1038/nature05572, Bibcode 2007Natur.446...88P, lire en ligne)
- (en) R. R. Ramsay, D. J. Steenkamp, M. Husain, « Reactions of electron-transfer flavoprotein and electron-transfer flavoprotein: ubiquinone oxidoreductase », Biochemical Journal, vol. 241, no 3, , p. 883-892 (PMID 3593226, PMCID 3593226)
- (en) Jian Zhang , Frank E. Frerman et Jung-Ja P. Kim, « Structure of electron transfer flavoprotein-ubiquinone oxidoreductase and electron transfer to the mitochondrial ubiquinone pool », Proceedings of the National Academy of Sciences of the United States of America, vol. 103, no 44, , p. 16212-16217 (PMID 17050691, PMCID 1637562, DOI 10.1073/pnas.0604567103, Bibcode 2006PNAS..10316212Z, lire en ligne)
- (en) Colleen M. Byron, Marian T. Stankovich, Mazhar Husain et Victor L. Davidson, « Unusual redox properties of electron-transfer flavoprotein from Methylophilus methylotrophus », Biochemistry, vol. 28, no 21, , p. 8582-8587 (PMID 2605209, DOI 10.1021/bi00447a047, lire en ligne)
- (en) Yasuyuki Ikeda, Christine Dabrowski et Kay Tanaka, « Separation and properties of five distinct acyl-CoA dehydrogenases from rat liver mitochondria. Identification of a new 2-methyl branched chain acyl-CoA dehydrogenase », Journal of Biological Chemistry, vol. 258, no 2, , p. 1066-1076 (PMID 6401712, lire en ligne)
- (en) Frank J. Ruzicka et Helmut Beinert, « A new iron-sulfur flavoprotein of the respiratory chain. A component of the fatty acid beta oxidation pathway », Journal of Biological Chemistry, vol. 252, no 23, , p. 8440-8445 (PMID 925004, www.jbc.org/cgi/reprint/252/23/8440.pdf)
- (en) Kimitsune Ishizaki, Tony R. Larson, Nicolas Schauer, Alisdair R. Fernie, Ian A. Graham et Christopher J. Leaver, « The Critical Role of Arabidopsis Electron-Transfer Flavoprotein:Ubiquinone Oxidoreductase during Dark-Induced Starvation », American Society of Plant Biologists, vol. 17, no 9, , p. 2587-2600 (PMID 16055629, PMCID 1197437, DOI 10.1105/tpc.105.035162, lire en ligne)
- (en) Edward A. Berry, Mariana Guergova-Kuras, Li-shar Huang et Antony R. Crofts, « Structure and Function of Cytochrome bc Complexes », Annual Review of Biochemistry, vol. 69, , p. 1005-1075 (PMID 10966481, DOI 10.1146/annurev.biochem.69.1.1005, lire en ligne)
- (en) Antony R. Crofts, « The Cytochrome bc1 Complex: Function in the Context of Structure », Annual Review of Physiology, vol. 66, , p. 689-733 (PMID 14977419, DOI 10.1146/annurev.physiol.66.032102.150251, lire en ligne)
- (en) So Iwata, Joong W. Lee, Kengo Okada, John Kyongwon Lee, Momi Iwata, Bjarne Rasmussen, Thomas A. Link, S. Ramaswamy et Bing K. Jap, « Complete Structure of the 11-Subunit Bovine Mitochondrial Cytochrome bc1 Complex », Science, vol. 281, no 5373, , p. 64-71 (PMID 9651245, DOI 10.1126/science.281.5373.64, Bibcode 1998Sci...281...64I, lire en ligne)
- (en) Bernard L. Trumpower, « The protonmotive Q cycle. Energy transduction by coupling of proton translocation to electron transfer by the cytochrome bc1 complex », Journal of Biological Chemistry, vol. 265, no 20, , p. 11409-11412 (PMID 2164001, lire en ligne)
- (en) Carola Hunte, Hildur Palsdottir et Bernard L Trumpower, « Protonmotive pathways and mechanisms in the cytochrome bc1 complex », FEBS Letters, vol. 545, no 1, , p. 39-46 (PMID 12788490, DOI 10.1016/S0014-5793(03)00391-0, lire en ligne)
- (en) Melissa W. Calhoun, Jeffrey W. Thomas et Robert B. Gennis, « The cytochrome oxidase superfamily of redox-driven proton pumps », Trends in Biochemical Sciences, vol. 19, no 8, , p. 325-330 (PMID 7940677, DOI 10.1016/0968-0004(94)90071-X, lire en ligne)
- (en) Tomitake Tsukihara, Hiroshi Aoyama, Eiki Yamashita, Takashi Tomizaki, Hiroshi Yamaguchi, Kyoko Shinzawa-Itoh, Ryosuke Nakashima, Rieko Yaono et Shinya Yoshikawa, « The Whole Structure of the 13-Subunit Oxidized Cytochrome c Oxidase at 2.8 Å », Science, vol. 272, no 5265, , p. 1136-1144 (PMID 8638158, DOI 10.1126/science.272.5265.1136, Bibcode 1996Sci...272.1136T, lire en ligne)
- (en) Shinya Yoshikawa, Kazumasa Muramoto, Kyoko Shinzawa-Itoh, Hiroshi Aoyama, Tomitake Tsukihara, Kunitoshi Shimokata, Yukie Katayama et Hideo Shimada, « Proton pumping mechanism of bovine heart cytochrome c oxidase », Biochimica et Biophysica Acta (BBA) - Bioenergetics, vol. 1757, nos 9-10, , p. 1110-1116 (PMID 16904626, DOI 10.1016/j.bbabio.2006.06.004, lire en ligne)
- (en) Allan G. Rasmusson, Kathleen L. Soole et Thomas E. Elthon, « Alternative NAD(P)H dehydrogenases of plant mitochondria », Annual Review of Plant Biology, vol. 55, , p. 23-39 (PMID 15725055, DOI 10.1146/annurev.arplant.55.031903.141720, lire en ligne)
- (en) R. Ian Menz et David A. Day, « Purification and Characterization of a 43-kDa Rotenone-insensitive NADH Dehydrogenase from Plant Mitochondria », Journal of Biological Chemistry, vol. 271, no 38, , p. 23117-23120 (PMID 8798503, DOI 10.1074/jbc.271.38.23117, lire en ligne)
- (en) Allison E. McDonald et Greg C. Vanlerberghe, « Branched Mitochondrial Electron Transport in the Animalia: Presence of Alternative Oxidase in Several Animal Phyla », IUBMB Life, vol. 56, no 6, , p. 333-341 (PMID 15370881, DOI 10.1080/1521-6540400000876, lire en ligne)
- (en) F. E. Sluse et W. Jarmuszkiewicz, « Alternative oxidase in the branched mitochondrial respiratory network: an overview on structure, function, regulation, and role », Brazilian Journal of Medical and Biological Research, vol. 31, no 6, , p. 733-747 (PMID 9698817, DOI 10.1590/S0100-879X1998000600003, lire en ligne)
- (en) Anthony L. Moore et James N. Siedow, « The regulation and nature of the cyanide-resistant alternative oxidase of plant mitochondria », Biochimica et Biophysica Acta (BBA) - Bioenergetics, vol. 1059, no 2, , p. 121-140 (PMID 1883834, DOI 10.1016/S0005-2728(05)80197-5, lire en ligne)
- (en) Greg C. Vanlerberghe et Lee McIntosh, « ALTERNATIVE OXIDASE: From Gene to Function », Annual Review of Plant Physiology and Plant Molecular Biology, vol. 48, , p. 703-734 (PMID 15012279, DOI 10.1146/annurev.arplant.48.1.703, lire en ligne)
- (en) Yusuke Ito, Daisuke Saisho, Mikio Nakazono, Nobuhiro Tsutsumi et Atsushi Hirai, « Transcript levels of tandem-arranged alternative oxidase genes in rice are increased by low temperature », Gene, vol. 203, no 2, , p. 121-129 (PMID 9426242, DOI 10.1016/S0378-1119(97)00502-7, lire en ligne)
- (en) Denis P. Maxwell, Yong Wang et Lee McIntosh, « The alternative oxidase lowers mitochondrial reactive oxygen production in plant cells », Proceedings of the National Academy of Sciences of the United States of America, vol. 96, no 14, , p. 8271-8276 (PMID 10393984, PMCID 22224, DOI 10.1073/pnas.96.14.8271, Bibcode 1999PNAS...96.8271M, lire en ligne)
- (en) Giorgio Lenaz, « A critical appraisal of the mitochondrial coenzyme Q pool », FEBS Letters, vol. 502, no 9, , p. 151-155 (PMID 11741580, DOI 10.1016/S0014-5793(01)03172-6, lire en ligne)
- (en) Jesco Heinemeyer, Hans-Peter Braun, Egbert J. Boekema et Roman Kouřil, « A Structural Model of the Cytochrome c Reductase/Oxidase Supercomplex from Yeast Mitochondria », Journal of Biological Chemistry, vol. 282, no 16, , p. 12240-12248 (PMID 17322303, DOI 10.1074/jbc.M610545200, lire en ligne)
- (en) S. Gupte, E. S. Wu, L. Hoechli, M. Hoechli, K. Jacobson, A. E. Sowers et C. R. Hackenbrock, « Relationship between lateral diffusion, collision frequency, and electron transfer of mitochondrial inner membrane oxidation-reduction components », Proceedings of the National Academy of Sciences of the United States of America, vol. 81, no 9, , p. 2606-2610 (PMID 6326133, PMCID 345118, DOI 10.1073/pnas.81.9.2606, Bibcode 1984PNAS...81.2606G, lire en ligne)
- (en) Hermann Schägger et Kathy Pfeiffer, « Supercomplexes in the respiratory chains of yeast and mammalian mitochondria », The EMBO Journal, vol. 19, no 8, , p. 1777-1783 (PMID 10775262, PMCID 302020, DOI 10.1093/emboj/19.8.1777, lire en ligne)
- (en) Hermann Schägger, « Respiratory chain supercomplexes of mitochondria and bacteria », Biochimica et Biophysica Acta (BBA) - Bioenergetics, vol. 1555, nos 1-3, , p. 154-159 (PMID 12206908, DOI 10.1016/S0005-2728(02)00271-2, lire en ligne)
- (en) Hermann Schägger et Kathy Pfeiffer, « The Ratio of Oxidative Phosphorylation Complexes I, II, III, IV, and V in Bovine Heart Mitochondria, and the Composition of Respiratory Chain Supercomplexes », Journal of Biological Chemistry, vol. 276, no 41, , p. 37861-37867 (PMID 11483615, DOI 10.1074/jbc.M106474200, lire en ligne)
- (en) Kenneth H. Nealson, « Post-Viking Microbiology: New Approaches, New Data, New Insights », Origins of life and evolution of the biosphere, vol. 29, no 1, , p. 73-93 (PMID 11536899, DOI 10.1023/A:1006515817767, lire en ligne)
- (en) Günter Schäfer, Martin Engelhard et Volker Müller, « Bioenergetics of the Archaea », Microbiology and Molecular Biology Reviews, vol. 63, no 3, , p. 570-620 (PMID 10477309, PMCID 103747, lire en ligne)
- (en) W. John Ingledew et Robert K. Poole, « The respiratory chains of Escherichia coli », Microbiology and Molecular Biology Reviews, vol. 48, no 3, , p. 222-271 (PMID 6387427, PMCID 373010, lire en ligne)
- (en) G. Unden et J. Bongaerts, « Alternative respiratory pathways of Escherichia coli: energetics and transcriptional regulation in response to electron acceptors », Biochimica et Biophysica Acta (BBA) - Bioenergetics, vol. 1320, no 3, , p. 217-234 (PMID 9230919, DOI 10.1016/S0005-2728(97)00034-0, lire en ligne)
- (en) Gary Cecchini, Imke Schröder, Robert P Gunsalus et Elena Maklashina, « Succinate dehydrogenase and fumarate reductase from Escherichia coli », Biochimica et Biophysica Acta (BBA) - Bioenergetics, vol. 1553, nos 1-2, , p. 140-157 (PMID 11803023, DOI 10.1016/S0005-2728(01)00238-9, lire en ligne)
- (en) A. Freitag et E. Bock, « Energy conservation in Nitrobacter », FEMS Microbiology Letters, vol. 66, nos 1-3, , p. 157-162 (DOI 10.1016/0378-1097(90)90275-U, lire en ligne)
- (en) Shawn R. Starkenburg, Patrick S. G. Chain, Luis A. Sayavedra-Soto, Loren Hauser, Miriam L. Land, Frank W. Larimer, Stephanie A. Malfatti, Martin G. Klotz, Peter J. Bottomley, Daniel J. Arp et William J. Hickey, « Genome Sequence of the Chemolithoautotrophic Nitrite-Oxidizing Bacterium Nitrobacter winogradskyi Nb-255 », Applied and Environmental Microbiology, vol. 72, no 3, , p. 2050-2063 (PMID 16517654, PMCID 1393235, DOI 10.1128/AEM.72.3.2050-2063.2006, lire en ligne)
- (en) Tateo Yamanaka et Yoshihiro Fukumori, « The nitrite oxidizing system of Nitrobacter winogradskyi », FEMS Microbiology Reviews, vol. 4, no 4, , p. 259-270 (PMID 2856189, DOI 10.1111/j.1574-6968.1988.tb02746.x, lire en ligne)
- (en) S. Iuchi et E. C. C. Lin, « Adaptation of Escherichia coli to redox environments by gene expression », Molecular Microbiology, vol. 9, no 1, , p. 9-15 (PMID 8412675, DOI 10.1111/j.1365-2958.1993.tb01664.x, lire en ligne)
- (en) M. W. Calhoun, K. L. Oden, R. B. Gennis, M. J. de Mattos et O. M. Neijssel, « Energetic efficiency of Escherichia coli: effects of mutations in components of the aerobic respiratory chain », Journal of Bacteriology, vol. 175, no 10, , p. 3020-3025 (PMID 8491720, PMCID 204621, lire en ligne)
- (en) Kelvin J.A. Davies, « Oxidative stress: the paradox of aerobic life », Biochemical Society Symposium, vol. 61, , p. 1-31 (PMID 8660387, DOI 10.1042/bss0610001, lire en ligne)
- (en) Suresh I.S. Rattana, « Theories of biological aging: Genes, proteins, and free radicals », Free Radical Research, vol. 40, no 12, , p. 1230-1238 (PMID 17090411, DOI 10.1080/10715760600911303, lire en ligne)
- (en) Marian Valko, Dieter Leibfritz, Jan Moncol, Mark T.D. Cronin, Milan Mazur et Joshua Telser, « Free radicals and antioxidants in normal physiological functions and human disease », The International Journal of Biochemistry & Cell Biology, vol. 39, no 1, , p. 44-84 (PMID 16978905, DOI 10.1016/j.biocel.2006.07.001, lire en ligne)
- (en) Sandeep Raha et Brian H. Robinson, « Mitochondria, oxygen free radicals, disease and ageing », Trends in Biochemical Science, vol. 25, no 10, , p. 502-508 (PMID 11050436, DOI 10.1016/S0968-0004(00)01674-1, lire en ligne)
- (en) « Oxidants, oxidative stress and the biology of ageing », Nature, vol. 408, no 6809, , p. 239-247 (PMID 11089981, DOI 10.1038/35041687, lire en ligne)
- (en) Bernhard Kadenbach, Rabia Ramzan, Li Wen et Sebastian Vogt, « New extension of the Mitchell Theory for oxidative phosphorylation in mitochondria of living organisms », Biochimica et Biophysica Acta (BBA) - General Subjects, vol. 1800, no 3, , p. 205-212 (PMID 19409964, DOI 10.1016/j.bbagen.2009.04.019, lire en ligne)
- (en) Karim S. Echtay, Damien Roussel, Julie St-Pierre, Mika B. Jekabsons, Susana Cadenas, Jeff A. Stuart, James A. Harper, Stephen J. Roebuck, Alastair Morrison, Susan Pickering, John C. Clapham et Martin D. Brand, « Superoxide activates mitochondrial uncoupling proteins », Nature, vol. 415, no 6867, , p. 96-99 (PMID 11780125, DOI 10.1038/415096a, Bibcode 2002Natur.415...96E, lire en ligne)
- (en) Adrian J. Lambert et Martin D. Brand, « Inhibitors of the quinone-binding site allow rapid superoxide production from mitochondrial NADH:ubiquinone oxidoreductase (complex I) », Journal of Biological Chemistry, vol. 279, no 38, , p. 39414-39420 (PMID 15262965, DOI 10.1074/jbc.M406576200, lire en ligne)
- (en) D. V. Dervartanian et C. Veeger, « Studies on succinate dehydrogenase: I. Spectral properties of the purified enzyme and formation of enzyme-competitive inhibitor complexes », Biochimica et Biophysica Acta (BBA) - Specialized Section on Enzymological Subjects, vol. 92, no 2, , p. 233-247 (PMID 14249115, DOI 10.1016/0926-6569(64)90182-8, lire en ligne)
- (en) Motonari Tsubaki et Shinya Yoshikawa, « Fourier-transform infrared study of cyanide binding to the Fea3-CuB binuclear site of bovine heart cytochrome c oxidase: implication of the redox-linked conformational change at the binuclear site », Biochemistry, vol. 32, no 1, , p. 164-173 (PMID 8380331, DOI 10.1021/bi00052a022, lire en ligne)
- (en) Saroj Joshi et Yougou Huang, « ATP synthase complex from bovine heart mitochondria: The oligomycin sensitivity conferring protein is essential for dicyclohexyl carbodiimide-sensitive ATPase », Biochimica et Biophysica Acta (BBA) - Biomembranes, vol. 1067, no 2, , p. 255-258 (PMID 1831660, DOI 10.1016/0005-2736(91)90051-9, lire en ligne)
- Le DNP a été largement prescrit contre l'obésité dans les années 1930 mais a par la suite été abandonné en raison de ses effets secondaires dangereux. Son utilisation illégale se poursuit cependant de nos jours.
- (en) Peter G. Heytler, « Uncouplers of oxidative phosphorylation », Methods in Enzymology, vol. 55, , p. 462-472 (PMID 156853, DOI 10.1016/0076-6879(79)55060-5, lire en ligne)
- (en) Daniel Ricquier et Frédéric Bouillaud, « The uncoupling protein homologues: UCP1, UCP2, UCP3, StUCP and AtUCP », Biochemical Journal, vol. 345, no Pt é, , p. 161-179 (PMID 10620491, PMCID 1220743, DOI 10.1042/bj3450161, lire en ligne)
- (en) Jiří Borecký et Aníbal E. Vercesi, « Plant uncoupling mitochondrial protein and alternative oxidase: energy metabolism and stress », Bioscience Reports, vol. 3, no 4, , p. 271-286 (PMID 16283557, DOI 10.1007/s10540-005-2889-2, lire en ligne)
Voir aussi
Articles connexes
Liens externes
 Portail de la biochimie
Portail de la biochimie  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire