Tyrannosauridae
Tyrannosauridés • Tyrannosaures

Sous-familles de rang inférieur
- † Albertosaurinae
- † Tyrannosaurinae
Les Tyrannosauridés (Tyrannosauridae) (ou parfois les tyrannosaures), terme qui signifie « lézard tyran », forment la famille de dinosaures théropodes cœlurosauriens comprenant le fameux Tyrannosaure et d'autres grand prédateurs. Ils sont classés en deux sous-familles dont le nombre de genres varient selon les auteurs. Leurs fossiles, datant de la fin du Crétacé, ont été retrouvés en Amérique du Nord, en Europe et en Asie.
Bien que descendant d'ancêtres de petite taille, les Tyrannosauridae étaient presque toujours les plus grands prédateurs de leurs écosystèmes respectifs, se trouvant ainsi au sommet de la chaîne alimentaire. La plus grande espèce a été Tyrannosaurus rex, un des plus grands prédateurs terrestres connus, qui mesurait jusqu'à 12,8 mètres de longueur[2] et pesait jusqu'à 6,4 tonnes[3]. Les Tyrannosauridae étaient des carnivores bipèdes à la tête massive et possédant de grandes dents. Malgré leur poids important, leurs membres postérieurs étaient longs et adaptés à une marche rapide. En revanche, les membres antérieurs étaient très petits et portaient seulement deux doigts fonctionnels.
Contrairement à la plupart des autres groupes de dinosaures, on a découvert des restes très complets de la plupart des Tyrannosauridae connus. Cela a permis nombre de recherches diverses sur leur biologie. Des études ont ainsi mis entre autres l'accent sur leur ontogénie, leur biomécanique et leur mode de vie. Des tissus mous, fossilisés et intacts, ont été retrouvés sur un spécimen de Tyrannosaurus rex.
Description
- Tyrannosaurus rex ;
- Tarbosaurus bataar ;
- Albertosaurus sarcophagus ;
- Daspletosaurus torosus ;
- Gorgosaurus libratus.
Tous les Tyrannosauridae connus étaient de grands animaux[4]. Le plus petit spécimen connu est un Alioramus dont la longueur est estimée entre 5 et 6 mètres du museau à l'extrémité de la queue[5] mais considéré comme un juvénile par Thomas R. Holtz et Philip J. Currie[4],[6]. Les divers squelettes d’Albertosaurus, Gorgosaurus et Daspletosaurus trouvés mesurent entre 8 et 10 mètres de long[7], tandis que Tarbosaurus atteignait une longueur de 12 mètres[8]. Tyrannosaurus, le plus grand des tyrannosaures connus, approchait 13 mètres de long pour les plus grands spécimens[9].
L'anatomie de la tête des Tyrannosauridae est bien décrite car on possède des crânes complets de tous les genres, à l'exception d’Alioramus, dont on n'a trouvé qu'un crâne partiel[10]. Tyrannosaurus, Tarbosaurus et Daspletosaurus avaient des têtes qui mesuraient plus d'un mètre de longueur[7] et le plus grand crâne découvert de Tyrannosaurus mesure plus de 1,75 mètre de long[11]. Les Tyrannosauridae adultes avaient des crânes massifs avec de nombreux os soudés entre eux et renforcés pour plus de solidité mais on trouve aussi des cavités creusées dans les os et de nombreuses grandes ouvertures qui contribuaient à réduire le poids du crâne. Plusieurs caractéristiques des crânes de Tyrannosauridae ont également été retrouvées chez leurs ancêtres immédiats, comme la présence de prémaxillaires et des os du nez soudés[4].
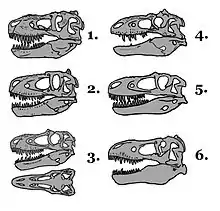
Les crânes des Tyrannosauridae présentent de nombreuses particularités, comme la présence d'une crête sur les os pariétaux, crête qui courait longitudinalement le long de la suture sagittale et séparait les deux fenêtres supratemporales sur le sommet du crâne. En arrière de ces fenêtres, les Tyrannosauridae avaient une autre crête à hauteur de la nuque, au niveau des os pariétaux, mais orientée dans un plan transversal plutôt que longitudinal. Cette crête était particulièrement bien développée chez Tyrannosaurus, Tarbosaurus et Alioramus. Albertosaurus, Daspletosaurus et Gorgosaurus avaient de hautes crêtes sur les os lacrymaux à l'angle interne des yeux, tandis que Tarbosaurus et Tyrannosaurus avaient des os postorbitaires extrêmement développés formant des crêtes osseuses très épaisses en forme de croissant au-dessus des yeux. Alioramus avait une rangée de six crêtes osseuses sur le dessus de son museau, au niveau de l'os du nez; des crêtes sur le dessous de la tête ont été signalées sur quelques spécimens de Daspletosaurus et de Tarbosaurus, ainsi que chez un Tyrannosauroidea, Appalachiosaurus[6],[12].
Les Tyrannosauridae, comme leurs ancêtres, étaient hétérodontes, avec les dents des prémaxillaires transversalement en forme de D et plus petites que les autres. Contrairement aux tyrannosaures primitifs et à la plupart des autres théropodes, les dents des maxillaires et des mandibules des Tyrannosauridae matures n'étaient pas tranchantes, mais très épaisses et souvent circulaires dans leur section transversale[4]. Le nombre de dents avait tendance à être fixe au sein des espèces et les espèces de grande taille semblent en avoir eu moins que les plus petites. Par exemple, Alioramus avait 76 à 78 dents tandis que Tyrannosaurus en avait entre 54 et 60[13].


.JPG.webp)
La tête était posée à l'extrémité d'un cou épais, en forme de S, et une longue et lourde queue faisait contrepoids pour équilibrer tête et torse, mettant le centre de gravité au niveau des hanches. Les Tyrannosauridae sont connus pour avoir eu, proportionnellement à leur taille, de petits membres antérieurs terminés par deux doigts, même si on a retrouvé quelquefois les restes d'un troisième doigt vestigial[4],[14]. Tarbosaurus est le genre qui avait les membres antérieurs les plus courts par rapport à la taille de son corps, tandis que Daspletosaurus avait les plus longs.
Les Tyrannosauridae marchaient exclusivement sur leurs membres postérieurs, de sorte que les os de leurs pattes arrière étaient massifs. Contrairement aux antérieurs, les membres postérieurs étaient plus longs, par rapport à la taille du corps, que chez la plupart des autres théropodes. Les jeunes et même certains adultes de petite taille, comme les Tyrannosauroidea primitifs, avaient des tibias plus longs que les fémurs, une caractéristique des dinosaures qui marchaient vite comme les Ornithomimidae. Les adultes les plus grands avaient des membres inférieurs caractéristiques d'animaux plus lents, mais pas autant que d'autres grands théropodes comme les Abelisauridae ou les Carnosauria. Les Tyrannosauridae avaient un petit troisième métatarsien coincé entre les deuxième et quatrième, constituant une structure connue sous le nom d'arctométatarse[4]. On ne sait pas quand ces arctométatarses sont apparus. Ils n'étaient pas présents chez les premiers Tyrannosauroidea comme Dilong[15] mais on les a retrouvés chez les derniers Appalachiosaurus[12]. Cette structure se retrouve également chez les Troodontidae, les Ornithomimidae et les Caenagnathidae[16], mais son absence chez les premiers Tyrannosauroidea indique qu'elle a été acquise par une évolution convergente[15].
Histoire de leur découverte

Les premiers restes de Tyrannosauridae ont été découverts lors d'expéditions menées par la Commission géologique du Canada qui trouva de nombreuses dents éparses. Joseph Leidy donna le nom de Deinodon (« dent épouvantable ») à ces dents de dinosaures en 1856. Les premiers spécimens de bonne qualité de Tyrannosauridae ont été trouvés dans la Formation de Horseshoe Canyon dans l'Alberta et se composaient de crânes presque complets et de parties de squelettes. Ces restes ont été étudiés par Edward Drinker Cope en 1876 qui les considéraient comme ceux d'une espèce de Tyrannosauroidea, le Dryptosaurus. En 1905, Henry Fairfield Osborn a reconnu que les restes trouvés dans l'Alberta étaient très différents de ceux du Dryptosaurus et a inventé un nouveau nom pour eux: Albertosaurus sarcophagus (« lézard mangeur de chair de l'Alberta »)[17]. Cope décrivit davantage de restes de Tyrannosauridae en 1892, avec des vertèbres isolées, et donna à cet animal le nom de Manospondylus gigas. Cette découverte a été négligée pendant plus d'un siècle, ce qui a causé une controverse au début des années 2000 quand on découvrit que ce matériel appartenait en fait à Tyrannosaurus rex alors que Manospondylus gigas aurait dû avoir l'antériorité[18].
Dans un article de 1905 intitulé Albertosaurus, Osborn décrivit deux spécimens de Tyrannosauridae supplémentaires qui avaient été découverts dans le Montana et le Wyoming lors d'une expédition en 1902 l’American Museum of Natural History, dirigée par Barnum Brown. Initialement, Osborn les considérait comme des espèces distinctes. Il nomma le premier Dynamosaurus imperiosus (« lézard puissant empereur »), et le second, Tyrannosaurus rex (« lézard tyran roi »). Un an plus tard, Osborn a reconnu que ces deux échantillons provenaient en fait de la même espèce. Bien que Dynamosaurus ait été trouvé en premier, le nom Tyrannosaurus figurait une page plus tôt dans l'article original décrivant les deux spécimens. Par conséquent, selon le Code international de nomenclature zoologique (CINZ), le nom de Tyrannosaurus a été utilisé[19].

Barnum Brown a ensuite recueilli plusieurs spécimens de nouveaux Tyrannosauridae dans l'Alberta, y compris le premier ayant encore ses deux pattes avant avec ses doigts caractéristiques (que Lawrence Lambe nomma Gorgosaurus libratus, « lézard féroce équilibré », en 1914). Une seconde découverte importante attribuée à un Gorgosaurus a été faite en 1942, sous la forme d'un ensemble bien conservé, même si, exceptionnellement, il avait un petit crâne complet. Le spécimen a dû attendre la fin de la Seconde Guerre mondiale pour être étudié par Charles W. Gilmore, qui le nomma Gorgosaurus lancesnis[17]. Ce crâne a été ré-étudié par Robert T. Bakker, Phil Currie, et Michael Williams en 1988, et affecté à un nouveau genre Nanotyrannus[20]. C'est également en 1946 que les paléontologues d'Union soviétique ont mené leurs premières expéditions en Mongolie et découvert les premiers restes de Tyrannosauridae d'Asie. En 1955, Evgeny Maleev décrivit de nouvelles espèces de Tyrannosauridae découvertes en Mongolie ainsi qu'un nouveau genre : Tarbosaurus (« lézard terrible »). Des études ultérieures, cependant, ont montré que toutes les espèces de Tyrannosauridae découvertes par Maleev étaient en fait une espèce de Tarbosaurus à différents stades de croissance. Une deuxième espèce de Tyrannosauridae mongol a été trouvée plus tard, décrite par Sergei Kurzanov en 1976, et a reçu le nom d'Alioramus remotus (« autre branche éloignée »), bien que son statut de vrai Tyrannosauridae et non pas de Tyrannosauroidea, plus primitif, soit encore controversé[5],[17],[21].
Répartition
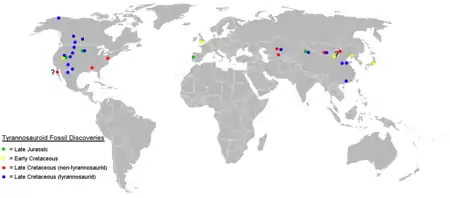
Alors qu'on trouve les premiers Tyrannosauroidea sur les trois continents de l'hémisphère nord, on n'a trouvé des fossiles de Tyrannosauridae qu'en Amérique du Nord et en Asie. On a parfois découvert des fragments de fossiles dans l'hémisphère sud qui ont été appelés « Tyrannosauridae de l'hémisphère sud » même si ceux-ci semblent avoir été confondus avec des fossiles d'Abelisauridae[22]. La date exacte d'apparition et le lieu d'origine de la famille restent inconnus en raison de la mauvaise qualité des fossiles trouvés sur les deux continents et datant du milieu du Crétacé, même si l'on sait que les premiers Tyrannosauridae ont vécu au début du Campanien sur l'île-continent Laramidia, correspondant à l'ouest de l'Amérique du Nord[4],[21].
On n'a jamais trouvé de restes de Tyrannosauridae dans l'est de l'Amérique du Nord, tandis que les plus primitifs des Tyrannosauroidea comme Dryptosaurus et Appalachiosaurus y ont persisté jusqu'à la fin du Crétacé, ce qui semble indiquer que les Tyrannosauridae doivent avoir évolué ou s'être dispersés dans l'ouest de l'Amérique du Nord après la division du continent par la Voie maritime intérieure de l'Ouest au milieu du Crétacé[12]. Des fossiles de Tyrannosauridae ont été trouvés en Alaska, qui reliait à l'époque l'est de l'Asie à l'ouest de l'Amérique et qui pourrait donc avoir servi de voie de passage entre l'Amérique du Nord et l'Asie[23]. Les genres Alioramus et Tarbosaurus qui se sont révélés être liés dans une analyse cladistique, forment l'unique branche asiatique de la famille[13].
Des deux sous-familles, les Tyrannosaurinae semblent avoir été les plus répandus. Les Albertosaurinae sont inconnus en Asie, qui a été le foyer des Tyrannosaurinae Tarbosaurus et Alioramus. Les deux sous-familles étaient présentes au Campanien et au début du Maastrichtien en Amérique du Nord, avec le Tyrannosaurinae Daspletosaurus vivant dans tout l'ouest intérieur tandis que les Albertosaurinae Albertosaurus et Gorgosaurus n'ont été actuellement trouvés que dans la partie nord-ouest du continent[24].
À la fin du Maastrichtien, les Albertosaurinae semblent avoir disparu d'Amérique alors que le Tyrannosaurinae Tyrannosaurus la parcourait de la Saskatchewan au Texas. Cette tendance se retrouve chez d'autres taxons de dinosaures d'Amérique du Nord. Au cours du Campanien et au début du Maastrichien, les sous-familles des Lambeosaurinae (famille des Hadrosauridae) et des Centrosaurinae (super-famille des Ceratopsia) étaient fréquents dans le nord-ouest, tandis que les Saurolophinae et les Chasmosaurinae étaient plus abondants dans le sud. À la fin du Crétacé, les Centrosaurinae ont disparu et les Lambeosaurinae sont devenus rares, tandis que les Saurolophinae et Chasmosaurinae sont communs à l'intérieur de la région occidentale[4].
Physiologie
Croissance

Le paléontologue Gregory Erickson et ses collègues ont étudié la croissance et les étapes de la vie des Tyrannosauridae. L'histologie osseuse permet de déterminer l'âge d'un spécimen au moment de sa mort. La vitesse de croissance peut être déterminée en reportant sur un graphique la taille des individus en fonction de leur âge. Erickson a ainsi démontré que, après une longue période de croissance lente, les Tyrannosauridae avaient une importante poussées de croissance durant environ quatre ans vers le milieu de leur vie. Cette phase de croissance rapide se terminait à la maturité sexuelle, la croissance se ralentissant considérablement chez les animaux adultes. Une courbe de croissance de Tyrannosauridae est en forme de S, avec une vitesse de croissance maximale autour de 14 ans[3].

Le plus petit Tyrannosaurus rex connu (LACM 28471, le théropode « Jordanie ») avait un poids estimé à seulement 30 kilogrammes à l'âge de deux ans, alors que le plus grand, (FMNH PR2081 « Sue ») devait peser environ 5 400 kilogrammes à 28 ans, un âge que l'on pense avoir été proche de l'âge limite pour cette espèce[3]. Les jeunes T. rex restaient en dessous de 1 800 kg jusqu'à environ 14 ans, puis la taille de leur corps commençait à augmenter de façon spectaculaire. Au cours de cette phase de croissance rapide, un jeune Tyrannosaurus rex prenait en moyenne 600 kg par an pendant les quatre années suivantes. La croissance commençait à ralentir après 16 ans et, à 18 ans, la courbe atteignait à nouveau un plateau, indiquant que la croissance avait considérablement ralenti[25]. Par exemple, seulement 600 kg séparaient « Sue » âgé de 28 ans du spécimen canadien RTMP 81.12.1 âgé de 22 ans[3]. Ce brusque changement de vitesse de croissance pouvait indiquer la maturité physique, une hypothèse qui est soutenue par la découverte d'une moelle osseuse particulière dans le fémur du T. Rex MOR 1125 trouvé dans le Montana également connu sous le nom de « B-Rex » et âgé de 18 ans[26]. Ce type de moelle osseuse, qui sert à produire le calcium nécessaire à la coquille des œufs se retrouve uniquement chez les oiseaux femelles en période d'ovulation, ce qui indique que B-rex était en âge de procréer[27].

Les autres Tyrannosauridae présentent des courbes de croissance similaires, bien que les vitesses de croissance soient plus faibles ce qui aboutit à un poids adulte inférieur[28]. Chez les Albertosaurinae, Daspletosaurus montrait une vitesse de croissance plus rapide en raison de son poids plus élevé à l'âge adulte. La vitesse de croissance maximale de Daspletosaurus était de 180 kilogrammes par an, basée sur un poids estimé de 1 800 kg chez les adultes. D'autres auteurs pensent que les Daspletosaurus adultes étaient plus lourds, ce qui changerait leur vitesse de croissance mais pas la tendance générale[3]. Le plus jeune Albertosaurus connu est un individu de deux ans trouvé à Dry Island dans l'Alberta et qui devait peser environ 50 kg et mesurer un peu plus de 2 mètres de longueur. Le spécimen de 10 mètres de long trouvé dans la même carrière, le plus vieux et le plus gros connu, avait 28 ans. Sa vitesse de croissance maximale rapide est estimée avoir eu lieu entre environ 12 et 16 ans, atteignant 122 kg par an, sur la base d'un poids adulte de 1 300 kg soit une vitesse environ cinq fois plus lente que pour T. rex. Pour Gorgosaurus la vitesse maximale de croissance calculée est d'environ 110 kg/an au cours de la phase de croissance rapide, ce qui est comparable à celle de l'Albertosaurus[3].
Vie et mort

La fin de la phase de croissance rapide semble s'être accompagnée du début de la maturité sexuelle chez Albertosaurus même si la croissance se poursuivait peut-être à un rythme plus lent tout au long de la vie des animaux[3],[28]. L'apparition de la maturité sexuelle en pleine période de croissance semble être un trait commun chez les petits[29] et les grands[30] dinosaures ainsi que chez les grands mammifères comme les humains et les éléphants[30]. Ce type de maturation relativement précoce diffère radicalement de celui observé chez les oiseaux, où la maturité sexuelle n'est atteinte qu'après la fin de la croissance[30],[31].
En réunissant plusieurs spécimens de chaque tranche d'âge, Erickson et ses collègues ont été en mesure de tirer des conclusions sur la vie des populations de Tyrannosauridae. Leur étude a montré que, alors que l'on retrouve rarement des fossiles de jeunes animaux, on en retrouve beaucoup plus de subadultes en phase de croissance rapide et d'adultes. Plus de la moitié des spécimens connus de T. rex semblent être morts dans les six années autour de la maturité sexuelle, une tendance que l'on retrouve également chez d'autres Tyrannosauridae et, aujourd'hui, chez certains grands oiseaux à longue durée de vie et certains mammifères. Ces espèces sont caractérisées par des taux élevés de mortalité infantile et, par la suite, par une mortalité relativement faible chez les jeunes. La mortalité augmente à nouveau après la maturité sexuelle, en raison notamment des aléas de la reproduction. Bien que ce constat puisse être dû à des biais de conservation ou de collecte d'ossements, Erickson émet l'hypothèse que la différence serait due à une faible mortalité chez les jeunes en dessous d'une certaine taille, ce qui est observé actuellement chez certains grands mammifères modernes comme les éléphants. Ce faible taux de mortalité pourrait être du à un manque de prédation, car les Tyrannosauridae dépassaient en taille tous les autres prédateurs contemporains dès l'âge de deux ans. Les paléontologues n'ont pas trouvé suffisamment de restes de Daspletosaurus pour faire une étude similaire, mais Erickson note que la même tendance générale semble s'appliquer[28].
Les Tyrannosauridae passaient la moitié de leur vie dans la période juvénile avant de grandir subitement jusqu'à leur taille maximale ou presque en quelques années seulement[3]. Ceci, associé à l'absence totale de prédateurs de taille intermédiaire entre les énormes Tyrannosauridae adultes et les autres petits théropodes suggère que ce créneau a pu être pourvu par de jeunes Tyrannosauridae. On voit un phénomène analogue chez les dragons de Komodo actuels, où les nouveau-nés commencent leur vie comme insectivores arboricoles et grandissent lentement avant de devenir des prédateurs capables de s'attaquer aux grands vertébrés[4]. On a ainsi, par exemple, découvert des groupes de fossiles d'Albertosaurus contenant des individus de tout âge[32],[33].
Locomotion

Les caractéristiques de déplacement des Tyrannosauridae ont été les mieux étudiées chez Tyrannosaurus et il y a deux questions principales auxquelles il fallait répondre: sa vitesse de rotation sur lui-même et sa vitesse maximale en ligne droite. Tyrannosaurus semble s'être retourné lentement, un quart de tour lui demandant une à deux secondes, un mouvement que les êtres humains, à la station verticale et dépourvus de queue, peuvent faire en une fraction de seconde[34]. La raison de cette différence est le moment d'inertie élevé du premier, dont une grande partie de la masse est située à distance de son centre de gravité, un peu comme un homme portant sur l'épaule un long et gros rondin de bois[35].
Les scientifiques ont trouvé un large éventail de vitesse maximale possible. La plupart se situent autour de 11 mètres par seconde (40 km/h), mais cela va de 5 à 11 mètres par seconde (18 à 40 km/h) à un peu plus de 20 mètres par seconde (70 km/h). Les chercheurs ont été conduits à utiliser diverses techniques détournées pour l'estimer car, bien qu'il existe de nombreuses empreintes de grands théropodes en marche, à ce jour on n'en a retrouvé aucune d'un grand théropode en train de courir et cette absence de type d'empreinte peut être due au fait que ces animaux ne couraient pas[36].

Jack Horner et Don Lessem ont fait valoir en 1993 que Tyrannosaurus rex était un animal lent qui ne pouvait probablement pas courir (pas de phase sans appui dans le pas)[37]. Toutefois, Holtz (1998) a conclu que les Tyrannosauridae et leurs proches ont été les plus rapides des grands Theropoda[38]. Christiansen (1998) estime que les os des membres postérieurs des Tyrannosauridae n'étaient pas significativement plus gros que ceux des éléphants, qui ont une vitesse de pointe relativement faible et qui ne courent pratiquement jamais et il en conclut que la vitesse maximale de ce dinosaure aurait été d'environ 11 mètres par seconde, soit à peu près la vitesse d'un coureur à pied humain[39]. Farlow et ses collègues (1995) ont fait valoir que, vu son poids de 6 à 8 tonnes, le Tyrannosaurus aurait été gravement ou mortellement blessé s'il était tombé en se déplaçant rapidement, car son torse aurait percuté le sol à une décélération de 6 g (six fois l'accélération de la pesanteur, soit environ 60 mètres par seconde carrée) et ses petites pattes antérieures ne lui auraient pas permis de réduire l'impact[40],[41]. Cependant, à l'inverse, les girafes galopent à 50 km/h bien qu'elles puissent se casser une patte ou pire, se tuer, et ce même sans nécessité comme dans un environnement sûr tel un zoo[42],[43]. Aussi il est fort possible que Tyrannosaurus ait pu courir également lorsque cela était nécessaire. Cette hypothèse a été étudiée aussi pour Allosaurus[44],[45]. La plupart des recherches récentes sur la vitesse de déplacement du Tyrannosaurus aboutissent à une plage de 17 à 40 kilomètres à l'heure c'est-à-dire celle correspondant à de la marche ou une course à vitesse modérée[36],[46],[47]. Une étude par modèle informatique réalisée en 2007 basée sur des données obtenues directement à partir de fossile, permettait de conclure que Tyrannosaurus rex avait une vitesse de pointe de 8 mètres par seconde (30 km/h)[48],[49] (probablement chez un juvénile)[50].
Vision

Les orbites de Tyrannosaurus sont orientées de façon que les yeux regardent devant, lui permettant ainsi d'avoir une vision binoculaire légèrement meilleure que celle des faucons actuels. Jack Horner a également souligné que les Tyrannosauridae avaient évolué au cours du temps en améliorant de façon constante leur vision binoculaire. Il est difficile de savoir comment la sélection naturelle a favorisé cette tendance au long cours chez des animaux qui, s'ils avaient été des charognards purs, n'auraient pas eu besoin de la perception binoculaire qui donne une vision stéréoscopique[51],[52]. Chez les animaux actuels, la vision binoculaire se trouve principalement chez les prédateurs (la principale exception est les primates qui en ont besoin pour sauter de branche en branche). Contrairement au Tyrannosaurus, Tarbosaurus avait un crâne proportionnellement plus étroit que les autres Tyrannosauridae avec les yeux regardant sur le côté. Cela suggère que Tarbosaurus comptait plus sur son odorat et son ouïe que sur sa vue[53]. Chez Gorgosaurus, l'orbite est circulaire plutôt qu'ovale ou allongée comme chez les autres genres de Tyrannosauridae[6]. Chez Daspletosaurus, elle forme un grand ovale, une forme intermédiaire entre la forme circulaire de Gorgosaurus et une forme allongée de Tyrannosaurus[4],[6],[54].
Crêtes osseuses

On trouve des crêtes osseuses sur les crânes de nombreux théropodes et notamment chez de nombreux Tyrannosauridae. Alioramus, une possible espèce de cette famille de Mongolie, porte une seule rangée de cinq bosses osseuses bien saillantes sur les os du nez; une ligne similaire mais beaucoup moins prononcée est présente sur le crâne d'Appalachiosaurus ainsi que quelques spécimens de Daspletosaurus, Albertosaurus et Tarbosaurus[12]. Chez Albertosaurus, Gorgosaurus et Daspletosaurus, on trouve une saillie proéminente, en dedans de chaque œil, sur l'os lacrymal. Cette saillie est absente chez Tarbosaurus et Tyrannosaurus qui avaient plutôt un bourrelet en forme de croissant au-dessus de chaque œil sur l'os postorbitaire. Ces crêtes peuvent avoir servi de repère, peut-être comme signe de reconnaissance d'espèce ou dans les parades amoureuses[4].
Thermorégulation
On a longtemps cru que Tyrannosaurus, comme la plupart des dinosaures, était un ectotherme (animal à « sang-froid ») au métabolisme analogue à celui des reptiles, mais cela a été contesté par des scientifiques comme Robert T. Bakker et John Ostrom dans les premières années de la Renaissance des Dinosaures, à la fin des années 1960[55],[56]. On a même affirmé que Tyrannosaurus rex était même endotherme (animal à « sang chaud »), ce qui suppose qu'il avait une vie très active[57]. Depuis lors, plusieurs paléontologues ont cherché à déterminer la capacité de Tyrannosaurus à réguler sa température corporelle. La preuve histologique d'une vitesse de croissance élevée chez les jeunes T. rex, comparable à celle des mammifères et des oiseaux, peut plaider en faveur de l'hypothèse d'un métabolisme élevé. Les courbes de croissance indiquent aussi que, comme chez les mammifères et les oiseaux, la croissance de T. rex s'arrêtait à l'arrivée à l'âge mature et n'était pas indéterminée comme chez la plupart des autres vertébrés[25]. On a cité une différence de température possible de 4 à 5 °C au plus entre les vertèbres thoraciques et le tibia. Ce faible écart de température entre le centre du corps et les extrémités a été revendiqué par le paléontologue Reese Barrick et le géochimiste William Douches pour indiquer que Tyrannosaurus rex maintenait une température corporelle interne constante (homéothermie) et qu'il avait un métabolisme intermédiaire entre celui des reptiles et des mammifères[58]. Plus tard, ils ont trouvé des résultats comparables pour des échantillons de Giganotosaurus, qui ont vécu sur un continent différent et des dizaines de millions d'années plus tôt[59]. Même si Tyrannosaurus rex montre des signes évidents d'homéothermie, cela ne signifie pas nécessairement qu'il était endotherme. Sa thermorégulation peut aussi s'expliquer simplement du fait de sa grande taille (gigantothermie), comme retrouvé chez certaines grandes tortues de mer actuelles[60],[61].
Plumage

De longues structures filamenteuses ont été retrouvées avec des restes de nombreux Coelurosauria de la Formation de Jehol datant du Crétacé inférieur et d'autres formations géologiques situées à proximité dans la province de Liaoning, en Chine[62]. Ces filaments ont généralement été interprétés comme étant des « protoplumes », homologues des plumes des oiseaux et de certains théropodes non-aviaires[63],[64] même si d'autres hypothèses ont été proposées notamment qu'il se soit agi de fibres de collagène[65]. On a ainsi décrit, en 2004, un squelette de Dilong qui était le premier exemple de Tyrannosauroidea porteur de protoplumes. De même on a trouvé chez cet animal ce qui pourrait être des protoplumes ramifiées mais pas pennées, analogues au duvet des oiseaux actuels et qui pourraient avoir servi à l'isolation thermique de l'animal[15].
On a supposé que les Tyrannosauridae avaient de telles protoplumes. Toutefois, les rares empreintes de peau de Tyrannosauridae montrent la présence d'écailles[66],[67]. Bien qu'il soit possible que des protoplumes aient existé sur des parties du corps qui n'ont pas été conservées, il se peut aussi que ces animaux n'aient pas eu de revêtement isolant, comme on le retrouve chez les mammifères modernes de plusieurs tonnes tels que les éléphants, les hippopotames et la plupart des espèces de rhinocéros. Lorsqu'un objet augmente de volume, sa capacité à retenir la chaleur fait de même en raison de la diminution de son ratio surface/volume. Par conséquent, comme ces grands animaux évoluaient dans des régions aux climats chauds, la présence d'un revêtement de fourrure ou de plumes perdait de son intérêt isolant et pouvait au contraire devenir un désavantage car il aurait pu provoquer une élévation excessive de température de l'animal. Ces protoplumes peuvent aussi avoir été perdues secondairement au cours de l'évolution des grands Tyrannosauridae, en particulier au cours des périodes chaudes du Crétacé[15].
Paléoécologie
Coexistence de Daspletosaurus et Gorgosaurus
Dans la Formation de Dinosaur Park, on a constaté que Gorgosaurus avait vécu à côté d'une autre espèce plus rare de Tyrannosaurinae Daspletosaurus. C'est l'un des rares exemples de coexistence de deux genres de Tyrannosauridae. Les prédateurs de taille similaire dans les guildes de prédateurs modernes sont séparés en différentes niches écologiques par des différences anatomiques, de comportement ou géographiques qui limitent la concurrence. La différenciation des niches entre les Tyrannosauridae du Dinosaur Park n'est pas bien comprise[68]. En 1970, Dale Russell a émis l'hypothèse que Gorgosaurus, le plus abondant, chassait de façon active les Hadrosauridae rapides à la course, tandis que Daspletosaurus, plus massif et plus rare, chassait les Ceratopsia et les Ankylosauria plus difficiles à chasser car armés de cornes et lourdement blindés[7]. On a toutefois retrouvé un spécimen de Daspletosaurus (OTM 200) de la même époque dans la formation de Two Medicine au Montana possédant encore les restes d'un jeune Hadrosauridae digéré dans son tube digestif[69].

Contrairement à certains autres groupes de dinosaures, il ne semble pas y avoir de genre de Tyrannosauridae plus abondant à une altitude qu'à une autre[68]. Toutefois, Gorgosaurus semble avoir été plus abondant dans les formations du Nord comme le Dinosaur Park alors que Daspletosaurus était plus abondant au sud. La même tendance est retrouvée chez les autres groupes de dinosaures. Les Ceratopsia et les Hadrosauridae sont également plus fréquents dans la formation de Two Medicine et dans le sud-ouest de l'Amérique du Nord durant le Campanien alors que les Centrosaurinae et les Lambeosaurinae dominaient dans les latitudes plus élevées. Holtz a suggéré cette répartition était due aux préférences écologiques entre Tyrannosaurinae, Chasmosaurinae et Saurolophinae. À la fin du Maastrichtien, Les Tyrannosaurinae comme Tyrannosaurus rex, Les Saurolophinae comme Edmontosaurus et les Chasmosaurinae comme Triceratops étaient répandus dans tout l'Ouest de l'Amérique du Nord, tandis que Les Albertosaurinae et les Centrosaurinae s'y étaient éteints et les Lambeosaurinae y étaient devenus rares[4].
Comportement social
On a peu de renseignements sur le comportement des Tyrannosauridae entre eux. Par exemple, le Tyrannosaurus rex « Sue » est mort apparemment d'une importante morsure à la tête qui ne peut lui avoir été infligée que par un autre Tyrannosaurus[70]. Des chercheurs ont fait remarquer qu'un squelette de subadulte et un squelette de juvénile ont été trouvés dans la même carrière que « Sue » et utilisé ce fait pour soutenir que les Tyrannosaurus ont peut-être vécu en groupes plus ou moins sociaux[71]. Bien qu'il n'y ait aucune preuve d'un comportement grégaire chez Gorgosaurus[32],[33] il existe des preuves de certains comportements sociaux chez Albertosaurus et Daspletosaurus.
Un jeune spécimen de Daspletosaurus (TMP 94.143.1) du Dinosaur Park montre des traces de morsures à la face qui lui ont été infligées par un autre Tyrannosauridae. Les marques de morsures sont cicatrisées ce qui prouve que l'animal a survécu à la morsure. Un adulte de Daspletosaurus (TMP 85.62.1) du même parc porte également des traces de morsure par un Tyrannosauridae, attestant que les attaques ne portaient pas uniquement sur des animaux très jeunes. Il est possible que ces morsures soient attribuables à d'autres espèces mais les agressions intraspécifiques, notamment les morsures à la face, sont très fréquentes chez les prédateurs. Ces types de morsures se retrouvent chez d'autres Tyrannosaures comme Gorgosaurus et Tyrannosaurus, ainsi que chez d'autres Théropodes comme Sinraptor et Saurornitholestes. Darren Tanke et Phil Currie ont émis l'hypothèse que ces morsures seraient dues à des compétitions intraspécifiques pour la possession d'un territoire ou de ressources, ou pour la domination au sein d'un groupe social[32].

La preuve que les Daspletosaurus vivaient en groupes provient d'un amas d'ossements trouvé dans la formation de Two Medicine. Cet amas comprend les restes de trois Daspletosaurus : un grand adulte, un petit jeune et un individu de taille intermédiaire. On a retrouvé au moins cinq squelettes d'Hadrosauridae au même endroit. Les indices géologiques montrent que les restes n'ont pas été rassemblés par un courant d'eau mais que tous les animaux ont été enterrés en même temps au même endroit. Les restes d'Hadrosauridae sont dispersés et portent de nombreuses traces de morsures par des Tyrannosauridae, indiquant que les Daspletosaurus se nourrissaient des Hadrosauria au moment de leur mort. La cause de leur décès est inconnue. Currie en conclut que les Daspletosaurus vivaient en groupe, bien que cela ne puisse pas être établi avec certitude[33]. D'autres scientifiques sont sceptiques quant à l'existence d'une vie sociale chez les Daspletosaurus et autres grands Theropoda[72], Brian Roach et Daniel Brinkman ont suggéré que les Daspletosaurus devaient mener une vie ressemblant davantage à celle des Dragons de Komodo actuels, où des individus sans vie sociale se partagent une même carcasse, s'attaquant et même se cannibalisant souvent dans l'affaire[73].
Le gisement de Dry Island découvert par Barnum Brown et son équipe contient les restes de vingt-deux Albertosaurus, le plus grand nombre de Theropoda du Crétacé trouvé en un même lieu, et le deuxième plus grand de Theropoda après le gisement d'Allosaurus de la carrière de dinosaures de Cleveland Lloyd. Le groupe semble être composé d'un adulte très vieux, de huit adultes entre 17 et 23 ans, de sept individus en phase de croissance rapide, donc entre 12 et 16 ans, et de six jeunes, âgés de 2 à 11 ans, qui n'avaient pas encore commencé leur phase de croissance accélérée[28]. La quasi-absence d'ossements d'herbivores et le même état de conservation des nombreux individus ont conduit Phil Currie à conclure que l'emplacement n'était pas un piège à prédateurs comme le site paléontologique de La Brea Tar Pits en Californie et que tous les animaux retrouvés étaient morts en même temps. Currie affirme que cela prouve l'existence d'un comportement social[74]. D'autres scientifiques sont sceptiques, observant que les animaux peuvent être morts de soif, noyés ou pour toute autre raison[28],[72],[73].
Stratégies alimentaires
Le paléontologue Philip J. Currie a émis une hypothèse sur les habitudes de chasse en groupe des Albertosaurus ; il suggère que les plus jeunes membres d'un groupe pouvaient être chargés de rabattre leurs proies vers les adultes, plus grands et plus puissants, mais aussi plus lents[74]. Les jeunes pouvaient avoir aussi des modes de vie différents de ceux des adultes, occupant un créneau de prédation entre celui des adultes et celui des Theropoda dont les plus gros pesaient environ deux fois moins qu'un Albertosaurus adulte[4]. Cependant, comme les traces de comportement dans les gisements de fossiles sont extrêmement rares, ces hypothèses ne peuvent pas être facilement vérifiées. Phil Currie estime que Daspletosaurus formait des groupes de chasse, bien que cela ne puisse pas être établi avec certitude[33]. Il n'existe aucune preuve d'un comportement grégaire des Gorgosaurus[32],[33].
Le débat quant à savoir si Tyrannosaurus rex était un prédateur ou un charognard pur est aussi vieux que celui sur sa locomotion. Lambe (1917) a décrit un squelette bien conservé de Gorgosaurus, proche parent du Tyrannosaurus, et a conclu que ce premier et donc aussi Tyrannosaurus rex était un charognard pur, car les dents de Gorgosaurus montraient peu d'usure[75]. Cet argument n'est plus pris au sérieux parce que les Theropoda remplaçaient leurs dents assez rapidement. Depuis la première découverte du Tyrannosaurus, la plupart des scientifiques sont convenus que c'était un prédateur, même si, comme les grands prédateurs modernes, il devait saisir l'opportunité de récupérer ou de voler la proie morte d'un autre prédateur s'il en avait l'occasion[76],[77].
Jack Horner, un expert connu des Hadrosauridae est actuellement le principal défenseur de l'idée que Tyrannosaurus rex était exclusivement un charognard et ne chassait jamais[37],[78],[79]. Horner a présenté plusieurs arguments pour étayer son hypothèse. La présence de grands bulbes et nerfs olfactifs suggère qu'il avait un odorat très développé, lui permettant de flairer les cadavres sur de grandes distances. Ses dents pouvaient broyer les os et, donc, extraire un maximum de nourriture (moelle osseuse) des restes de carcasses qui sont, généralement, les parties les moins nutritives. Enfin, au moins certaines de ses proies potentielles pouvaient se déplacer rapidement alors qu'on a presque la preuve qu'il ne savait pas courir[78],[80].
D'autres indices suggèrent au contraire que Tyrannosaurus était un chasseur. Par exemple, les orbites de Tyrannosaurus sont positionnées de façon que les yeux regardent en avant, lui permettant d'avoir une vision binoculaire légèrement meilleure que celle des faucons modernes. On a aussi retrouvé des blessures cicatrisées infligées par un Tyrannosaurus sur des squelettes d'Hadrosauridae et de Triceratops qui ont donc du survivre à une attaque[81],[82],[83]. Certains chercheurs soutiennent que si Tyrannosaurus avait été un charognard, un autre dinosaure aurait dû jouer le rôle de plus grand prédateur dans ces régions au Crétacé supérieur. Les plus grandes proies étaient les grands Marginocephalia et Ornithopoda. Les autres Tyrannosauridae avaient des caractéristiques physiques si proches de Tyrannosaurus que seuls les petits Dromaeosauridae auraient pu jouer ce rôle de grands prédateurs. Dans cette optique, les partisans d'un comportement de charognard ont émis l'hypothèse que la taille et la force des Tyrannosauridae leur permettaient de voler les proies des petits prédateurs[80]. La plupart des paléontologues s'accordent à dire que Tyrannosaurus rex était à la fois un prédateur actif et un charognard.
Classification
Le nom de Deinodontidae a été inventé par Edward Drinker Cope en 1866 pour cette famille[84] et a continué d'être utilisé à la place de celui de Tyrannosauridae jusque dans les années 1960[8]. Le genre type de la famille était Deinodon, nom donné d'après des dents isolées trouvées au Montana[85]. Toutefois, dans une étude de 1970 sur les Tyrannosaurus en Amérique du Nord, Dale Russell a conclu que Deinodon n'était pas un taxon valide, et a utilisé le nom de Tyrannosauridae à la place de celui de Deinodontidae, indiquant que cela était conforme aux règles de l'ICZN[7]. Par conséquent, le nom Tyrannosauridae est maintenant préféré par les experts modernes[17].
Tyrannosaurus était un nom créé par Henry Fairfield Osborn en 1905, appliqué à la famille des Tyrannosauridae de l'époque[86]. Le nom est dérivé de l'ancien grec τυραννος/tyrannos (« tyran ») et σαυρος/sauros (« lézard »). Le très commun suffixe -idae est ajouté aux noms de famille zoologique et est dérivé du suffixe grec -ιδαι/-idai, qui indique un nom pluriel[87].
Taxinomie
Tyrannosauridae est une famille dans la classification linnéenne. Elle fait partie de la super-famille des Tyrannosauroidea, du clade des Coelurosauria et du sous-ordre des Theropoda.
Les Tyrannosauridae sont divisés en deux sous-familles. Les Albertosaurinae d'Amérique du Nord comprennent les genres Albertosaurus et Gorgosaurus tandis que les Tyrannosaurinae comprennent les genres Daspletosaurus, Tarbosaurus, Nanuqsaurus et Tyrannosaurus. Certains auteurs incluent les espèces Gorgosaurus libratus dans le genre Albertosaurus et Tarbosaurus bataar dans le genre Tyrannosaurus, tandis que d'autres préfèrent conserver Gorgosaurus et Tarbosaurus dans des genres séparés. Les Albertosaurinae se caractérisent par une charpente osseuse plus fine, un crâne plus petit et des tibias proportionnellement plus grands que ceux des Tyrannosaurinae. De plus, chez les Tyrannosaurinae, la crête sagittale trouvée sur les pariétaux continue en avant sur les frontaux.
Liste des genres selon Brusatte & Carr (2016)[88] :
- sous-famille † Albertosaurinae Currie et al. 2003 :
- † Albertosaurus Osborn, 1905
- † Gorgosaurus Lambe, 1914
- sous-famille † Tyrannosaurinae † Osborn 1906 :
- † Alioramus Kurzanov, 1976
- † Daspletosaurus Russell, 1970
- † Lythronax Loewen & al., 2013
- † Nanuqsaurus Fiorillo & Tykoski, 2014
- † Qianzhousaurus Lü, 2014
- † Tarbosaurus Maleev, 1955
- † Teratophoneus Carr & al., 2011
- † Tyrannosaurus Osborn, 1905
- † Zhuchengtyrannus Hone & al., 2011











Phylogénie
| Cladogrammes des Tyrannosauridae : | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
*Note: Carr et al. considèrent Gorgosaurus libratus comme une espèce d’Albertosaurus et Tarbosaurus bataar comme une espèce de Tyrannosaurus
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Avec l'avènement de la systématique phylogénétique en paléontologie des vertébrés, on a donné plusieurs classifications des Tyrannosauridae. La première, due à Paul Sereno en 1998, inclut tous les Tyrannosauroidea les plus proches de Tyrannosaurus autres que Alectrosaurus, Aublysodon ou Nanotyrannus[89]. Toutefois, Nanotyrannus est souvent considéré comme un jeune Tyrannosaurus rex tandis que Aublysodon est généralement considéré comme un nomen dubium impropre à l'usage dans la définition d'un clade[4]. Les nouvelles définitions sont basées sur des genres plus précis.
Une tentative de classification faite par Christopher Brochu en 2003 inclut Albertosaurus, Alectrosaurus, Alioramus, Daspletosaurus, Gorgosaurus, Tarbosaurus et Tyrannosaurus dans la définition[2]. Holtz a redéfini le clade en 2004 pour utiliser tout ce qui précède sauf Alioramus et Alectrosaurus, qu'ils ne pouvaient pas classer avec certitude. Toutefois, dans le même journal, Holtz a également fourni une définition complètement différente, comprenant tous les Theropoda plus apparentés à Tyrannosaurus que Eotyrannus[4]. La définition la plus récente est celle de Sereno en 2005, qui définit les Tyrannosauridae comme le clade contenant au moins Albertosaurus, Gorgosaurus et Tyrannosaurus[90].
Les analyses cladistiques de la phylogénie des Tyrannosauridae trouvent souvent que Tarbosaurus et Tyrannosaurus sont des taxons sœur, alors que Torosus est plus primitif. Une relation étroite entre Tarbosaurus et Tyrannosaurus est soutenue par de nombreuses caractéristiques du crâne, y compris le modèle de sutures entre les os de certains, la présence d'une crête en forme de croissant sur l'os postorbitaire au-dessus de chaque œil et un maxillaire très profond avec une courbe notable à la baisse sur le bord inférieur entre autres[4],[12]. Une autre hypothèse a été présentée dans une étude réalisée en 2003 par Phil Currie et ses collègues, qui ont trouvé peu d'arguments favorables pour que Daspletosaurus soit un membre basal d'un clade comprenant également Tarbosaurus et Alioramus, ces deux derniers provenant d'Asie, basé sur l'absence d'un volet osseux reliant les os du nez et les os lacrymaux[3]. Alioramus s'est avéré être le plus proche parent de Tarbosaurus dans cette étude, basée sur un modèle de répartition similaire des contraintes dans le crâne.
Une étude connexe a également noté un mécanisme de verrouillage de la mâchoire inférieure partagé entre les deux genres[91]. Dans un document séparé, Currie a noté la possibilité que Alioramus puisse être un jeune Tarbosaurus mais il a déclaré que le nombre de dents beaucoup plus élevé et les crêtes nasales plus importantes d'Alioramus suggèrent qu'il s'agit d'un genre distinct. De même, Currie utilise le nombre de dents élevé de Nanotyrannus pour penser qu'il s'agit peut-être d'un genre distinct[6], plutôt que d'un jeune Tyrannosaurus comme la plupart des autres experts le croient[4],[54]. Cependant, la découverte puis la description de Qianzhousaurus en 2014 a révélé qu'Alioramus ne serait pas un proche parent de Tarbosaurus, mais qu'il appartiendrait à une nouvelle branche de tyrannorauridés, la tribu des Alioramini. Le découverte de Qianzhousaurus a révélé en outre que de tels tyrannosauridés possédant un long museau étaient largement répandus dans toute l'Asie et auraient partagé le même environnement, tout en évitant la concurrence avec les tyrannosaurinés plus grands et plus robustes en chassant des proies différentes[92].
Phylogénie des genres de Tyrannosauridés d'après Brusatte & Carr (2016)[88] :
| Tyrannosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Annexes
Articles connexes
Bibliographie
- (en) Kenneth Carpenter, The Carnivorous Dinosaurs, Bloomington, Indiana University Press, , 392 p. (ISBN 978-0-253-34539-4, lire en ligne)
- (en) Peter L. Larson et Kenneth Carpenter, Tyrannosaurus rex, the Tyrant King (Life of the Past), Bloomington, Indiana University Press, , Har/Cdr éd., 456 p. (ISBN 978-0-253-35087-9, lire en ligne)
- (en) David B. Weishampel, Peter Dodson et Halszka Osmólska, The Dinosauria, University of California Press, , 2e éd., 880 p. (ISBN 978-0-520-25408-4, lire en ligne)
Références taxinomiques
- (en) Référence Paleobiology Database : Tyrannosauridae Osborn 1906 (éteint)
- (en) Référence Tree of Life Web Project : Tyrannosauridae
- (en) Référence BioLib : Tyrannosauridae Osborn, 1906 †
- (en) Référence uBio : Tyrannosauridae Osborn 1906
- (en) Référence NCBI : Tyrannosauridae (taxons inclus)
Liens externes
- • Bibliothèque du Congrès
- (en) Référence Tree of Life Web Project : Tyrannosauridae
- (en) Référence Paleobiology Database : Tyrannosauridae Osborn 1906
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Tyrannosauridae » (voir la liste des auteurs).
- (en) H. F. Osborn, « Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (Second communication) », Bulletin of the American Museum of Natural History, vol. 22, no 16, , p. 281-296 (lire en ligne)
- (en) C.R. Brochu, « Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull », Memoirs of the Society of Vertebrate Paleontology, vol. 7, , p. 1–138 (DOI 10.2307/3889334, lire en ligne)
- (en) Gregory M. Erickson, Peter J. Makovicky, Philip J. Currie, Mark A. Norell, Scott A. Yerby et Christopher A. Brochu, « Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs », Nature, vol. 430, no 7001, , p. 772–775 (PMID 15306807, DOI 10.1038/nature02699)
- (en) Thomas R. Holtz, « Tyrannosauroidea », dans David B. Weishampel, Peter Dodson et Halszka Osmólska, The Dinosauria, Berkeley, University of California Press, (ISBN 0-520-24209-2), p. 111–136
- (ru) Sergei M. Kurzanov, « A new carnosaur from the Late Cretaceous of Nogon-Tsav, Mongolia », The Joint Soviet-Mongolian Paleontological Expedition Transactions, vol. 3, , p. 93–104
- (en) Philip J. Currie, « Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta », Acta Palaeontologica Polonica, vol. 48, no 2, , p. 191–226 (lire en ligne [PDF])
- (en) Dale A. Russell, « Tyrannosaurs from the Late Cretaceous of western Canada », National Museum of Natural Sciences Publications in Paleontology, vol. 1, , p. 1–34
- (ru) Evgeny A. Maleev (trad. (en) F. J. Alcock), « New carnivorous dinosaurs from the Upper Cretaceous of Mongolia », Doklady Akademii Nauk SSSR, vol. 104, no 5, , p. 779–783 (lire en ligne [PDF])
- (en) « Sue's vital stats », Sue at the Field Museum, sur http://archive.fieldmuseum.org/, Field Museum of Natural History (consulté le )
- (en) Philip J. Currie, « Theropods from the Cretaceous of Mongolia », dans The Age of Dinosaurs in Russia and Mongolia, Cambridge, Cambridge University Press, (ISBN 978-0-521-54582-2), p. 434–455
- (en) « Museum unveils world's largest T-rex skull », sur Montana State University, (consulté le )
- (en) Thomas D. Carr, Thomas E. Williamson et David R. Schwimmer, « A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama », Journal of Vertebrate Paleontology, vol. 25, no 1, , p. 119–143 (DOI 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2)
- (en) Philip J. Currie, Jørn H. Hurum et Karol Sabath, « Skull structure and evolution in tyrannosaurid phylogeny », Acta Palaeontologica Polonica, vol. 48, no 2, , p. 227–234 (lire en ligne [PDF])
- (en) Elizibeth D. Quinlan, Kraig Derstler et Mercedes M. Miller, « Anatomy and function of digit III of the Tyrannosaurus rex manus », Geological Society of America Annual Meeting — Abstracts with Programs, , p. 77 (résumé)
- (en) Xing Xu, Mark A. Norell, Xuewen Kuang, Xiaolin Wang, Zhao Qi et Chengkai Jia, « Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids », Nature, vol. 431, no 7009, , p. 680–684 (PMID 15470426, DOI 10.1038/nature02855)
- Thomas R. Holtz, « The phylogenetic position of the Tyrannosauridae: implications for theropod systematics », Journal of Palaeontology, vol. 68, no 5, , p. 1100–1117 (lire en ligne)
- (en) K. Carpenter, « Tyrannosaurs (Dinosauria) of Asia and North America », dans N. Mateer et P.-J. Chen (eds.), Aspects of Nonmarine Cretaceous Geology, Beijing, China Ocean Press, , p. 250-268
- (en) B. H. Breithaupt, E. H. Southwell et N. A. Matthews (18 octobre 2005) « In Celebration of 100 years of Tyrannosaurus rex: Manospondylus gigas, Ornithomimus grandis, and Dynamosaurus imperiosus, the Earliest Discoveries of Tyrannosaurus rex in the West » dans 2005 Salt Lake City Annual Meeting 37: 406 p., Geological Society of America. Consulté le 8 octobre 2008.
- (en) B. H. Breithaup, E.H. Southwell et N. A. Matthews, « Dynamosaurus Imperiosus and the earliest discoveries of Tyrannosaurus Rex in Wyoming and the West », New Mexico Museum of Natural History and Science Bulletin, vol. 35, , p. 257–258 (lire en ligne [PDF], consulté le )
- (en) R. T. Bakker, M. Williams et P. Currie, « Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana », Hunteria, vol. 1, , p. 1–30
- (en) M. A. Loewen, R. B. Irmis, J. J. W. Sertich, P. J. Currie et S. D. Sampson, « Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Océans », PLoS One, vol. 8, no 11, , e79420 (DOI 10.1371/journal.pone.0079420)
- (en) Peter Dodson, Brooks Britt et al., Age of Dinosaurs, Lincolnwood, Publications International, LTD, , 192 p. (ISBN 978-0-7853-0443-2, OCLC 29824077, LCCN 93084891), « Abelisaurus », p. 105
- (en) Anthony R. Fiorillo et Roland A. Gangloff, « Theropod teeth from the Prince Creek Formation (Cretaceous) of northern Alaska, with speculations on arctic dinosaur paleoecology », Journal of Vertebrate Paleontology, vol. 20, no 4, , p. 675–682 (DOI 10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2)
- (en) Thomas R. Holtz, Jr., « The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics », Journal of Paleontology, vol. 68, no 5, , p. 1100–1117 (lire en ligne)
- (en) J. R. Horner et K. Padian, « Age and growth dynamics of Tyrannosaurus rex », Proceedings of the Royal Society of London B, vol. 271, no 1551, , p. 1875–1880 (PMID 15347508, PMCID 1691809, DOI 10.1098/rspb.2004.2829)
- (en) Andrew H. Lee et Sarah Werning, « Sexual maturity in growing dinosaurs does not fit reptilian growth models », Proceedings of the National Academy of Sciences, vol. 105, no 2, , p. 582–587 (PMID 18195356, PMCID 2206579, DOI 10.1073/pnas.0708903105)
- (en) M.H. Schweitzer, J. L. Wittmeyer et J. R. Horner, « Gender-specific reproductive tissues in ratites and Tyrannosaurus rex », Science, vol. 308, no 5727, , p. 1456–1460 (PMID 15933198, DOI 10.1126/science.1112158)
- (en) G. M. Erickson, P. J. Currie, B. D. Inouye et A. A. Winn, « Tyrannosaur life tables: an example of nonavian dinosaur population biology », Science, vol. 313, no 5784, , p. 213–217 (PMID 16840697, DOI 10.1126/science.1125721)
- (en) Gregory M. Erickson, Kristi Curry Rogers, David J. Varricchio, Mark A. Norell et Xing Xu, « Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition », Biology Letters, vol. 3, no 5, , p. 558–561 (PMID 17638674, PMCID 2396186, DOI 10.1098/rsbl.2007.0254)
- (en) Andrew H. Lee et Sarah Werning, « Sexual maturity in growing dinosaurs does not fit reptilian growth models », PNAS, vol. 105, no 2, , p. 582–587 (PMID 18195356, PMCID 2206579, DOI 10.1073/pnas.0708903105)
- (en) Robert E. Ricklefs, « Tyrannosaur ageing », Biology Letters, vol. 3, no 2, , p. 214–217 (PMID 17284406, PMCID 2375931, DOI 10.1098/rsbl.2006.0597)
- (en) Darren H. Tanke et Philip J. Currie, « Head-biting behavior in theropod dinosaurs: paleopathological evidence », Gaia, vol. 15, , p. 167–184 (lire en ligne [PDF])
- (en) Philip J. Currie, David Trexler, Eva B. Koppelhus, Kelly Wicks et Nate Murphy, « An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA) », dans The Carnivorous Dinosaurs, Bloomington, Indiana University Press, (ISBN 978-0-253-34539-4, LCCN 2004023225), p. 313–324
- (en) Janette Ellis, « Tyrannosaurus had poor turning circle », sur Cosmos Magazine, (consulté le )
- (en) D. R. Carrier, R. M. Walter et D. V. Lee, « Influence of rotational inertia on turning performance of theropod dinosaurs: clues from humans with increased rotational inertia », Journal of Experimental Biology, The Company of Biologists Limited, vol. 204, , p. 3917–3926 (PMID 11807109, lire en ligne)
- (en) J. R. Hutchinson, « Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa », Journal of Morphology, vol. 262, no 1, , p. 441–461 (PMID 15352202, DOI 10.1002/jmor.10240, lire en ligne [PDF])
- (en) John R. Horner et Don Lessem, The complete T. rex, New York, Simon & Schuster, , 239 p. (ISBN 978-0-671-74185-3, LCCN 93000211)
- (en) Thomas R. Holtz, « Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda) », Journal of Paleontology, vol. 70, no 3, , p. 536–538 (JSTOR 1306452, résumé)
- (en) P. Christiansen, « Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential », Gaia, vol. 15, , p. 241–255 (ISSN 0871-5424, lire en ligne [PDF])
- (en) James O. Farlow, Matthew B. Smith et John M. Robinson, « Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex », Journal of Vertebrate Paleontology, vol. 15, no 4, , p. 713–725 (DOI 10.1080/02724634.1995.10011257, lire en ligne)
- (en) « The bigger they come, the harder they fall », sur New Scientist,
- (en) « Giraffe », WildlifeSafari.info (consulté le )
- « The History of Woodland Park Zoo — Chapter 4 » (consulté le )
- (en) R. M. Alexander, « Dinosaur biomechanics », Proc Biol Sci., The Royal Society, vol. 273, no 1596, , p. 1849–1855 (PMID 16822743, PMCID 1634776, DOI 10.1098/rspb.2006.3532)
- (en) Rebecca R. Hanna, « Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd dinosaur quarry collection », Journal of Vertebrate Paleontology, vol. 22, no 1, , p. 76–90 (DOI 10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2) attribue les blessures de l'Allosaurus connu sous le nom de « Big Al » – du moins une au moins – à une chute.
- (en) John R. Hutchinson et Mariano Garcia, « Tyrannosaurus was not a fast runner », Nature, vol. 415, no 6875, , p. 1018–1021 (PMID 11875567, DOI 10.1038/4151018a, résumé)
- R. Hajdul (1997). Tendons. "Dinosaur Cards". Orbis Publishing Ltd. D36044311.
- (en) W. I. Sellers et P. L. Manning, « Estimating dinosaur maximum running speeds using evolutionary robotics », Proc. R. Soc. B, The Royal Society, vol. 274, no 1626, , p. 2711 (PMID 17711833, PMCID 2279215, DOI 10.1098/rspb.2007.0846, lire en ligne)
- (en) Liz Seward, « T. rex 'would outrun footballer' », L'article cite Dr Bill Sellers, université de Manchester, coauteur d'un article publié dans Proceedings of the Royal Society B, sur BBC News, .
- (en) G. Callison et H. M. Quimby, « Tiny dinosaurs: Are they fully grown? », Journal of Vertebrate Paleontology, vol. 3, , p. 200–209 (DOI 10.1080/02724634.1984.10011975)
- (en) Kent A. Stevens, « Binocular vision in theropod dinosaurs », Journal of Vertebrate Paleontology, vol. 26, no 2, , p. 321–330 (DOI 10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2, lire en ligne)
- (en) Eric Jaffe, « Sight for 'Saur Eyes: ' T. rex vision was among nature’s best », Science News, vol. 170, no 1, , p. 3 (DOI 10.2307/4017288, JSTOR 4017288, lire en ligne, consulté le )
- (en) Sergei V. Saveliev et Vladimir R. Alifanov, « A new study of the brain of the predatory dinosaur Tarbosaurus bataar (Theropoda, Tyrannosauridae) », Paleontological Journal, vol. 41, no 3, , p. 281–289 (DOI 10.1134/S0031030107030070)
- (en) Thomas D. Carr, « Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria) », Journal of Vertebrate Paleontology, vol. 19, no 3, , p. 497–520 (DOI 10.1080/02724634.1999.10011161)
- (en) Robert T. Bakker, « The superiority of dinosaurs », Discovery, vol. 3, no 2, , p. 11–12
- (en) Robert T. Bakker, « Anatomical and ecological evidence of endothermy in dinosaurs », Nature, vol. 238, , p. 81–85 (DOI 10.1038/238081a0)
- (en) Robert T. Bakker, The Dinosaur Heresies, New York, Kensington Publishing, , 1re éd., 481 p. (ISBN 978-0-688-04287-5, OCLC 13699558, LCCN 86012643)
- (en) Reese E. Barrick et William J. Showers, « Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes », Science, vol. 265, no 5169, , p. 222–224 (PMID 17750663, DOI 10.1126/science.265.5169.222, résumé)
- (en) Reese E. Barrick et William J. Showers, « Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus », Palaeontologia Electronica, vol. 2, no 2, (lire en ligne, consulté le )
- (en) Frank V. Paladino, James R. Spotila et Peter Dodson, The Complete Dinosaur, Bloomington, Indiana University Press, , 752 p., poche (ISBN 978-0-253-21313-6, lire en ligne), « A blueprint for giants: modeling the physiology of large dinosaurs », p. 491–504
- (en) Anusuya Chinsamy et Willem J. Hillenius, The dinosauria, Berkeley, University of California Press, , 2e éd., 861 p. (ISBN 978-0-520-24209-8, LCCN 2004049804, lire en ligne), « Physiology of nonavian dinosaurs », p. 643–659
- (en) Zhonghe Zhou, Paul M. Barrett et Jason Hilton, « An exceptionally preserved Lower Cretaceous ecosystem », Nature, vol. 421, no 6925, , p. 807–814 (PMID 12594504, DOI 10.1038/nature01420)
- (en) Pei-ji Chen, Zhi-ming Dong et Shuo-nan Zhen, « An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China », Nature, vol. 391, no 6663, , p. 147–152 (DOI 10.1038/34356)
- (en) Xing Xu, Zhonghe Zhou et Richard O. Prum, « Branched integumental structures in Sinornithosaurus and the origin of feathers », Nature, vol. 410, no 6825, , p. 200–204 (PMID 11242078, DOI 10.1038/35065589)
- (en) Theagarten Lingham-Soliar, Alan Feduccia et Xiaolin Wang, « A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres », Proceedings of the Royal Society of London. Series B, Biological Sciences, vol. 270, no 1620, , p. 1823–1829 (PMID 17521978, PMCID 2270928, DOI 10.1098/rspb.2007.0352)
- (en) Brian Switek, « Palaeontology: The truth about T. rex », Nature, vol. 502, no 7472, , p. 424-426 (DOI 10.1038/502424a, lire en ligne)
- (en) Phil R. Bell, Nicolás E. Campione, W. Scott Persons, Philip J. Currie, Peter L. Larson, Darren H. Tanke et Robert T. Bakker, « Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution », Biology Letter, (DOI 10.1098/rsbl.2017.0092, lire en ligne).
- (en) James O. Farlow et Eric R. Pianka, « Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs », Historical Biology, vol. 16, no 1, , p. 21–40 (DOI 10.1080/0891296031000154687)
- (en) David J. Varricchio, « Gut contents from a Cretaceous tyrannosaurid: implications for theropod dinosaur digestive tracts », Journal of Paleontology, vol. 75, no 2, , p. 401–406 (DOI 10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2)
- (en) C. A. Brochu, « Lessons From A Tyrannosaur: The Ambassadorial Role Of Paleontology », PALAIOS, vol. 18, no 6, , p. 475 (DOI 10.1669/0883-1351(2003)018<0475:LFATTA>2.0.CO;2)
- Guinness World Records Ltd. (2003). 2003 Guinness World Records. pg 90.
- (en) David A. Eberth et Richard T. McCrea, « Were large theropods gregarious? », Journal of Vertebrate Paleontology, vol. 21, no (Supplement to 3 - Abstracts of Papers, 61st Annual Meeting of the Society of Vertebrate Paleontology), , p. 46A
- (en) Brian T. Roach et Daniel L. Brinkman, « A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs », Bulletin of the Peabody Museum of Natural History, vol. 48, no 1, , p. 103–138 (DOI 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2)
- (en) Philip J. Currie, « Possible evidence of gregarious behavior in tyrannosaurids », Gaia, vol. 15, , p. 271–277 (lire en ligne [PDF])
- (en) L. B. Lambe, « The Cretaceous theropodous dinosaur Gorgosaurus », Memoirs of the Geological Survey of Canada, vol. 100, , p. 1–84 (lire en ligne [PDF])
- J. O. Farlow and T. R. Holtz, « The Fossil Record of Predation » [PDF], The Paleontological Society Papers, M. Kowalewski and P. H Kelley, , p. 251–266
- M. Dorey. (1997). Tyrannosaurus. "Dinosaur Cards". Orbis Publishing Ltd. D36045907.
- (en) J.R. Horner, « Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger) », The Paleontological Society Special Publication, vol. 7, , p. 157–164
- (en) J. Amos, « T. rex goes on trial », BBC, (lire en ligne)
- (en) M. Walters et J. Paker, Dictionary of Prehistoric Life, Claremont Books, (ISBN 1-85471-648-4).
- (en) G. M. Erickson et K. H. Olson, « Bite marks attributable to Tyrannosaurus rex: preliminary description and implications », Journal of Vertebrate Paleontology, vol. 16, no 1, , p. 175–178 (DOI 10.1080/02724634.1996.10011297).
- (en) K. Carpenter, « Evidence of predatory behavior by theropod dinosaurs », Gaia, vol. 15, , p. 135–144 (lire en ligne [PDF]).
- (en) Denver W. Fowler et Robert M. Sullivan, « A ceratopsid pelvis with toothmarks from the Upper Cretaceous Kirtland Formation, New Mexico: evidence of late Campanian tyrannosaurid feeding behavior », New Mexico Museum of Natural History and Science Bulletin, vol. 35, , p. 127–130 (lire en ligne, consulté le ).
- (en) E. D. Cope, « Discovery of a gigantic dinosaur in the Cretaceous of New Jersey », Proceedings of the Academy of Natural Sciences of Philadelphia, vol. 18, , p. 275-279.
- (en) Joseph Leidy, « Notice of remains of extinct reptiles and fishes, discovered by Dr. F.V. Hayden in the badlands of the Judith River, Nebraska Territory », Proceedings of the Academy of Natural Sciences of Philadelphia, vol. 8, , p. 72–73
- (en) Henry F. Osborn, « Tyrannosaurus and other Cretaceous carnivorous dinosaurs », Bulletin of the American Museum of Natural History, vol. 21, , p. 259–265 (DOI 10.1111/j.1468-5965.2007.00735_17.x, lire en ligne)
- A. Bailly (1901) Abrégé du dictionnaire GREC-FRANÇAIS
- (en) Stephen Brusatte et Thomas D. Carr, « The phylogeny and evolutionary history of tyrannosauroid dinosaurs », Scientific Reports, no 20252, (DOI 10.1038/srep20252)
- (en) Paul C. Sereno, « A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria », Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, vol. 210, , p. 41–83
- Paul C. Sereno, « Stem Archosauria—TaxonSearch », (consulté le )
- (en) Jørn H. Hurum et Sabath Karol, « Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared », Acta Palaeontologica Polonica, vol. 48, no 2, , p. 161–190 (lire en ligne)
- (en) Junchang Lü, Laiping Yi, Stephen L. Brusatte, Ling Yang, Hua Li et Liu Chen, « A new clade of Asian Late Cretaceous long-snouted tyrannosaurids », Nature Communications, vol. 5, (DOI 10.1038/ncomms4788).
 Portail des dinosaures
Portail des dinosaures  Portail de l’herpétologie
Portail de l’herpétologie  Portail de la paléontologie
Portail de la paléontologie