Aldolase
L’aldolase, ou fructose-bisphosphate aldolase, est une lyase qui catalyse les réactions :
- (1) D-fructose-1-phosphate D-glycéraldéhyde + dihydroxyacétone phosphate ;
- (2) D-fructose-1,6-bisphosphate D-glycéraldéhyde-3-phosphate + dihydroxyacétone phosphate.


| N° EC | EC |
|---|---|
| N° CAS | |
| Cofacteur(s) | Zinc |
| IUBMB | Entrée IUBMB |
|---|---|
| IntEnz | Vue IntEnz |
| BRENDA | Entrée BRENDA |
| KEGG | Entrée KEGG |
| MetaCyc | Voie métabolique |
| PRIAM | Profil |
| PDB | RCSB PDB PDBe PDBj PDBsum |
| GO | AmiGO / EGO |
Cette enzyme intervient à la fois dans la glycolyse, la néoglucogenèse et le cycle de Calvin. Elle intervient notamment à la 4e étape de la glycolyse pour favoriser la réaction inverse d'une aldolisation, ce qui revient à cliver le fructose-1,6-bisphosphate en glycéraldéhyde-3-phosphate (G3P) et dihydroxyacétone phosphate (DHAP). Elle est également susceptible de produire du DHAP à partir d'autres (3S,4R)-cétose-1-phosphates tels que le fructose-1-phosphate et le sédoheptulose-1,7-bisphosphate.
Il en existe trois isoformes, correspondant à trois isoenzymes, exprimées de façon variable selon les tissus :
- l’aldolase A, encodée chez l'homme par le gène ALDOA sur le chromosome 16 et exprimée lors du développement de l'embryon, dans le muscle strié adulte, les lymphocytes et les érythrocytes, avec une préférence pour le fructose-1,6-bisphosphate comme substrat et donc une préférence pour la réaction (2) ;
- l’aldolase B, encodée chez l'homme par le gène ALDOB sur le chromosome 9 et exprimée dans le foie, avec une affinité comparable pour le fructose-1-phosphate et le fructose-1,6-bisphosphate, d'où une efficacité équivalente pour les réactions (1) et (2) ;
- l’aldolase C, encodée chez l'homme par le gène ALDOC sur le chromosome 17 et exprimée essentiellement dans l'hippocampe et les cellules de Purkinje du cerveau et, dans une moindre mesure, dans d'autres types de cellules telles que les thrombocytes et les mastocytes, avec une orientation préférentielle pour la réaction (2).
Structure et mécanisme
| Pfam | PF00274 |
|---|---|
| InterPro | IPR000741 |
| PROSITE | PDOC00143 |
| SCOP | 1ald |
| SUPERFAMILY | 1ald |
| CDD | cd00344 |
| Pfam | PF01116 |
|---|---|
| Clan Pfam | CL0036 |
| InterPro | IPR000771 |
| PROSITE | PDOC00523 |
| SCOP | 1dos |
| SUPERFAMILY | 1dos |
| CDD | cd00453 |
On distingue deux classes de protéines, dont les séquences peptidiques partagent peu de points communs, dotées d'activité enzymatique aldolase :
- les protéines de classe I forment une base de Schiff protonée entre un résidu de lysine hautement conservé du site actif et l'atome de carbone du carbonyle du dihydroxyacétone phosphate (DHAP);
- les protéines de classe II polarisent le groupe carbonyle à l'aide d'un cation divalent tel que celui de zinc Zn2+.
Par exemple, la protéine gatY de l'opéron dulcitol et la protéine agaY de l'opéron N-acétylgalactosamine d’Escherichia coli sont homologues de l'aldolase de classe II. Il a été établi que deux résidus d'histidine de la première moitié de la séquence de ces homologues interviennent dans la liaison avec le zinc[2].
Les sous-unités protéiques de ces deux classes possèdent chacune un domaine α/β repliées en tonneau TIM dans lequel se trouve le site actif. Plusieurs sous-unités sont assemblées pour constituer les protéines complètes et fonctionnelles.
Hormis quelques exceptions, seules des aldolases de classe I ont été observées chez les animaux, les plantes et les algues vertes[3], tandis que seules des aldolases de classe II ont été observées chez les mycètes. En revanche, les deux classes ont été abondamment observées chez d'autres eucaryotes et chez les bactéries. Elles sont alors souvent observées ensemble dans le même organisme. Les plantes et les algues possèdent également des aldolases dans leurs plastes, en plus de leur aldolase cytosolique. Une enzyme bifonctionnelle fructose-bisphosphate aldolase/phosphatase possédant un mécanisme de classe I a été souvent observée chez les archées ainsi que chez quelques bactéries[4] ; son site actif se trouve également dans un tonneau TIM.
Action dans la glycolyse et la néoglucogenèse
La glycolyse et la néoglucogenèse partagent une suite de six réactions chimiques réversibles. Au cours de la néoglucogenèse, le phosphoénolpyruvate (PEP) est réduit en fructose-1,6-bisphosphate, avec une aldolase du cytoplasme à la dernière étape, qui catalyse l'aldolisation du glycéraldéhyde-3-phosphate (G3P) avec le dihydroxyacétone phosphate (DHAP). Réciproquement, au cours de la glycolyse, le fructose-1,6-bisphosphate est oxydé en PEP avec une aldolase du cytoplasme à la première étape. La néoglucogenèse, qui est une voie métabolique anabolique, utilise la réaction réciproque (ci-dessous de droite à gauche) de celle de la glycolyse (de gauche à droite), qui est une réaction catabolique :


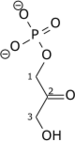
Il existe trois isoenzymes de l'aldolase — l'aldolase A, l'aldolase B et l'aldolase C — codées par des gènes différents dont l'expression change au cours du développement. L'aldolase A est ainsi produite chez l'embryon ainsi que dans les muscles adultes et les érythrocytes, un déficit en aldolase A ayant été associé aux myopathies et aux anémies hémolytiques, tandis que l'aldolase B est exprimée dans le foie, où elle peut également métaboliser le fructose[5], et que l'aldolase C est exprimée dans le cerveau. Les aldolases A et C interviennent essentiellement dans la glycolyse tandis que l'aldolase B intervient également dans la néoglucogenèse[6]. On pense que l'enzyme bifonctionnelle archéenne interviendrait dans la néoglucogenèse car elle forme du fructose-6-phosphate[7].
Action dans le cycle de Calvin
Le cycle de Calvin est une voie métabolique de fixation du carbone. Elle partage avec la néoglucogenèse une suite de quatre réactions chimiques réversibles. Au cours de ces deux voies métaboliques, le 3-phosphoglycérate est réduit en fructose-1,6-bisphosphate, dont la dernière étape est catalysée par l'aldolase. Le fructose-1,6-bisphosphate est hydrolysé au cours d'une cinquième réaction, catalysée par la fructose-1,6-bisphosphatase, pour produire du fructose-6-phosphate. La variation d'enthalpie libre très négative de cette réaction rend cette dernière irréversible.
Au cours du cycle de Calvin, l'aldolase catalyse également la formation du sédoheptulose-1,7-bisphosphate à partir du dihydroxyacétone phosphate et de l'érythrose-4-phosphate. Les produits principaux du cycle de Calvin sont des trioses phosphates — dihydroxyacétone phosphate et glycéraldéhyde-3-phosphate — et le fructose-6-phosphate, qui sont également nécessaires pour régénérer les ribulose-1,5-bisphosphate.
Les aldolases des plantes et des algues sont généralement des protéines codées par le génome nucléaire et importées dans les plastes.
Autres propriétés
L'aldolase assure également diverses fonctions non catalytiques en raison de son affinité pour plusieurs autres protéines et molécules telles que l'actine F, la tubuline α, les chaînes légères de la dynéine, la protéine du syndrome de Wiskott-Aldrich (en) (WASP), l'échangeur d'anions 1 (en), la phospholipase D (protéine PLD2 (en)), le transporteur de glucose GLUT4, l'inositol trisphosphate, la V-ATPase et l'ARNO (un facteur d'échange nucléotidique (en) du facteur d'ADP-ribosylation 6 (en)). On pense que ces associations assurent principalement la stabilité de la structure cellulaire, mais peuvent également assurer d'autres fonctions telles que dans l'endocytose, l'invasion par des parasites, les réarrangements du cytosquelette, la motilité cellulaire, la circulation et le recyclage des protéines dans les membranes, la transduction du signal et la compartimentation des tissus[8],[9],[10].
Notes et références
- (en) Andrew Dalby, Zbigniev Dauter et Jennifer A. Littlechild, « Crystal structure of human muscle aldolase complexed with fructose 1,6-bisphosphate: Mechanistic implications », Protein Science, vol. 8, no 2, , p. 291-297 (PMID 10048322, PMCID 2144250, DOI 10.1110/ps.8.2.291, lire en ligne)
- (en) Shaza M. Zgiby, Graeme J. Thomson, Seema Qamar et Alan Berry, « Exploring substrate binding and discrimination in fructose1,6-bisphosphate and tagatose 1,6-bisphosphate aldolases », The FEBS Journal, vol. 267, no 6, , p. 1858-1868 (PMID 10712619, DOI 10.1046/j.1432-1327.2000.01191.x, lire en ligne)
- (en) Nicola J. Patron, Matthew B. Rogers et Patrick J. Keeling, « Gene Replacement of Fructose-1,6-Bisphosphate Aldolase Supports the Hypothesis of a Single Photosynthetic Ancestor of Chromalveolates », Eukaryotic Cell, vol. 3, no 5, , p. 1169-1175 (PMID 15470245, PMCID 522617, DOI 10.1128/EC.3.5.1169-1175.2004, lire en ligne)
- (en) Bettina Siebers, Henner Brinkmann, Christine Dörr, Britta Tjaden, Hauke Lilie, John van der Oost et Corné H. Verhees, « Archaeal Fructose-1,6-bisphosphate Aldolases Constitute a New Family of Archaeal Type Class I Aldolase », Journal of Biological Chemistry, vol. 276, no 31, , p. 28710-28718 (PMID 11387336, DOI 10.1074/jbc.M103447200, lire en ligne)
- (en) A. Gopher, N. Vaisman, H. Mandel et A. Lapidot, « Determination of fructose metabolic pathways in normal and fructose-intolerant children: a 13C NMR study using [U-1313C]fructose », Proceedings of the National Academy of Sciences of the United States of America, vol. 87, no 14, , p. 5449-5453 (PMID 2371280, PMCID 54342, DOI 10.1073/pnas.87.14.5449, lire en ligne)
- (en) Ernst U. Walther, Martin Dichgans, Stephen M. Maricich, Rita R. Romito, Fei Yang, Suzan Dziennis, Saul Zackson, Richard Hawkes et Karl Herrup, « Genomic sequences of aldolase C (Zebrin II) direct lacZ expression exclusively in non-neuronal cells of transgenic mice », Proceedings of the National Academy of Sciences of the United States of America, vol. 95, no 5, , p. 2615-2620 (PMID 9482935, PMCID 19434, DOI 10.1073/pnas.95.5.2615, lire en ligne)
- (en) Sebastian Estelmann, Michael Hügler, Wolfgang Eisenreich, Katharina Werner, Ivan A. Berg, W. Hugo Ramos-Vera, Rafael F. Say, Daniel Kockelkorn, Nasser Gad′on et Georg Fuchs, « Labeling and Enzyme Studies of the Central Carbon Metabolism in Metallosphaera sedula », Journal of bacteriology, vol. 193, no 5, , p. 1191-1200 (PMID 21169486, PMCID 3067578, DOI 10.1128/JB.01155-10, lire en ligne)
- (en) Erumbi S. Rangarajan, HaJeung Park, Emanuelle Fortin, Jurgen Sygusch et Tina Izard, « Mechanism of Aldolase Control of Sorting Nexin 9 Function in Endocytosis », Journal of Biological Chemistry, vol. 285, no 16, , p. 11983-11990 (PMID 20129922, PMCID 2852936, DOI 10.1074/jbc.M109.092049, lire en ligne)
- (en) A. H. Ahn, S. Dziennis, R. Hawkes et K. Herrup, « The cloning of zebrin II reveals its identity with aldolase C », Development, vol. 120, no 8, , p. 2081-2090 (PMID 7925012, lire en ligne)
- (en) Maria Merkulova, Andrés Hurtado-Lorenzo, Hiroyuki Hosokawa, Zhenjie Zhuang, Dennis Brown, Dennis A. Ausiello et Vladimir Marshansky, « Aldolase directly interacts with ARNO and modulates cell morphology and acidic vesicle distribution », American Journal of Physiology. Cell Physiology, vol. 300, no 6, , C1442-C1455 (PMID 21307348, PMCID 3118619, DOI 10.1152/ajpcell.00076.2010, lire en ligne)
 Portail de la biochimie
Portail de la biochimie  Portail de la médecine
Portail de la médecine