Érythrocyte
L’érythrocyte (du grec erythros : rouge et kutos : cellule), aussi appelée hématie, ou plus communément globule rouge, fait partie des éléments figurés du sang. Chez les mammifères, c'est une cellule anucléée (dépourvue de noyau), tandis que chez les oiseaux c'est une cellule nucléée.
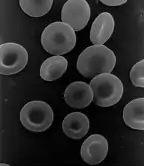
Son cytoplasme est riche en hémoglobine, qui assure le transport du dioxygène (O2), mais très pauvre en organites qui n'existent qu'à l'état de trace.
Le terme d'anémie s'applique parfois (dans le langage courant en particulier) à une diminution du nombre de globules rouges, mais en réalité elle est définie par une diminution du taux d'hémoglobine (les deux étant souvent simultanées). Le volume relatif des globules rouges ou hématocrite est le volume occupé par les hématies dans un volume donné du sang total.
L'hématie est un paramètre mesuré lors d'un hémogramme.
Aspect d'un globule rouge mammalien normal
Un érythrocyte normal se présente de profil comme un disque biconcave, de face comme un disque à centre plus clair : c'est une sorte de poche contenant l'hémoglobine.
Cette forme lui confère une élasticité importante, qui permet le transport de dioxygène à travers certains capillaires étroits. Le diamètre normal des globules rouges de face varie de 6,7 à 7,7 micromètres (moyenne 7,2 micromètres).
Ils sont en forme de disques biconcaves (région centrale : 0,8 micromètre, région périphérique : 2,6 micromètres)[1] la plus apte à une fixation maximale. Vus au microscope optique à l'état frais, ils sont de couleur rouge-orangé ; sur un frottis mince coloré au May-Grünwald Giemsa ou à la coloration de Wright, ils apparaissent plus rose. Leur forme est très bien visualisée au microscope électronique à balayage (MEB). Sur frottis épais, les globules rouges se disposent volontiers en rouleaux (particulièrement en présence d'un excès de fibrinogène ou d'une paraprotéine) et sont alors vus de profil.
Physiologie
La biosynthèse des globules rouges commence au stade embryonnaire, à partir de la 3e semaine de développement, au niveau du sac vitellin. La synthèse d'érythrocytes par le sac vitellin diminue vers la 5e semaine pour disparaître complètement lors de la 9e semaine de développement. Le relai est pris au cours du 3e mois par le foie, puis par la moelle osseuse hématopoïétique à partir du 5e mois de développement[2]. Cette dernière restera le seul site de synthèse des érythrocytes chez l'adulte.
Chez l'adulte, les globules rouges sont élaborés dans la moelle osseuse dite moelle hématopoïétique, que l’on retrouve dans les os plats (côtes, sternum, calvaria, os coxaux, clavicules) et aux extrémités (épiphyses) des os longs. La fabrication d'hématies par la moelle osseuse est appelée érythropoïèse. Tout commence avec des cellules souches hématopoïétiques, qui sont dites pluripotentes (elles pourront donner naissance à plusieurs types cellulaires). Certaines vont ensuite commencer à se différencier, et vont former les progéniteurs (BFU-E, CFU-E). Les premières cellules de la lignée érythrocytaire morphologiquement identifiables sont appelées proérythroblastes (pronormoblastes). Par division cellulaire (mitose) on passe à l'érythroblaste (normoblaste) basophile (en référence à la coloration du cytoplasme après coloration de May-Grünwald-Giemsa), puis à l'érythroblaste (normoblaste) polychromatophile de type I, puis de type II (également appelé orthochromatique, car la couleur de son cytoplasme est quasi identique à celle de l'hématie) ; toutes ces cellules sont nucléées et -sauf circonstances pathologiques- sont exclusivement médullaires. À chaque étape, on observe une diminution de taille de la cellule (et de son noyau). Au fur et à mesure, les cellules vont se charger en hémoglobine, responsable de la couleur rouge de leur cytoplasme, et la concentration cytoplasmique en hémoglobine augmente. À la fin (« à l'entrée dans la circulation », pourrait-on dire), l'érythroblaste perd son noyau. Quelques mitochondries et des fragments de REG (réticulum endoplasmique granuleux) ou appareil de Golgi persistent : c'est un réticulocyte qui a l'apparence d'un globule rouge en microscopie optique (une coloration spécifique est nécessaire pour le mettre en évidence, au bleu de crésyl brillant ou par fluorescence). Après quoi, l’expulsion de ces derniers résidus donnera naissance au globule rouge (érythrocyte ou hématie) mûr.
L'érythropoïèse est régulée par différents facteurs de croissance. L'érythropoïétine (EPO) va agir en stimulant les progéniteurs, surtout les CFU-E, et ainsi favoriser in fine la production de globule rouge. L'érythropoïétine est majoritairement produite par le cortex rénal (environ 90 % de la production) mais peut aussi être produite par le foie, le cerveau, l'utérus et peut même être produite artificiellement. Elle est actuellement utilisée à titre thérapeutique pour stimuler la production de globules rouges dans le traitement de certaines anémies, insuffisance rénale, au cours de certains traitements aplasiants… et comme agent dopant chez certains athlètes.
Chez l'humain, leur durée de vie atteint 120 jours, et près de 1 % des globules rouges d'un individu sont remplacés quotidiennement[3].
Valeurs plasmatiques normales
- Diamètre : 7 µm.
- Hématocrite : 47 % chez l'homme (±7 %) ; 42 % chez la femme (±5 %).
- Nombre moyen de globules rouges : 4,6 à 6,2 T/L chez l'homme et de 4,2 à 5,4 T/L chez la femme.
- Taux globulaire moyen d'hémoglobine (TGMH = Hémoglobine par Globule rouge) : de 28 à 34 pg.
La moelle osseuse
La moelle osseuse, située à l'intérieur des os, assure le renouvellement des cellules du sang. On distingue la moelle jaune, constituée de tissu adipeux, de la moelle rouge qui produit les cellules souches capables de se différencier en cellules sanguines : c'est le processus d'hématopoïèse. Les érythrocytes, ou globules rouges, sont des cellules anuclées constituant 99 % des cellules circulant dans le sang. Il y a également d’autres cellules avec cette fois un noyau, ce sont les leucocytes, ou globules blancs, qui constituent 1 % des cellules du sang et servent à la défense de l'organisme. Les thrombocytes, ou plaquettes, sont des fragments de cytoplasme de plus grandes cellules, les mégacaryocytes.
Composition
- On trouve environ 30 pg d'hémoglobine par hématie.
- Ces cellules se présentent sous formes de disques aplatis avec deux principales formes :
- chez les espèces à hématie nucléée, le disque épais de l'hématie est à peine renflé en son centre, là où est situé le noyau ;
- chez les espèces à hématie énucléée, le disque épais présente une forme biconcave, avec un centre plus fin que les bords.
- La membrane érythrocytaire est constituée d'une bicouche lipidique dont la partie centrale (entre les surfaces externe et interne), est hydrophobe. Elle est traversée par divers glycolipides et protéines plus ou moins glycosylées, glycophorines, aquaporines, AChe, Glut 1, Na+/K+ ATPase et protéine bande 3 (transférant HCO3- contre Cl-), entre autres. Ces protéines sont également présentes sur la membrane, lui offrant une protection mécanique et antiagrégante (glycocalix), ou contre l'action du complément (DAF et CD59), permettant les échanges entre l'érythrocyte et le milieu extérieur ou l'ancrage de cette membrane lipidique au cytosquelette interne. Leur polymorphisme est à l'origine des groupes sanguins (ABO, MNS, Rh, Kell, etc. présents à la surface des hématies).[style à revoir]
- Leur cytosquelette a été découvert au début des années 1980 et comprend : spectrine (dimère), protéine bande 4.1 (fixant l'actine), ankyrine, glycophorine (dimère), protéine bande 3, et des microfilaments intermédiaires à l'intérieur, leur permettant de conserver leur forme caractéristique, tout en leur conférant une très grande souplesse, indispensable pour passer dans les capillaires sanguins les plus fins.
- Chez tous les mammifères, les érythrocytes n'ont pas de noyau et par conséquent pas d'ADN nucléaire, ni de mitochondries. En revanche, les actinoptérygiens, les amphibiens et les sauropsides (reptiles + oiseaux) possèdent des érythrocites nucléés (Sporn and Dingman, 1963, Science).
Rôle des hématies
- Le transport de l'oxygène des poumons aux tissus et cellules du corps, grâce à l'hémoglobine contenue dans l'ergastoplasme (réticulum endoplasmique granuleux), à l'intérieur des globules rouges.
- La régulation du pH sanguin et le transport du CO2 grâce à l'anhydrase carbonique, une enzyme présente à la surface des hématies qui transforme les bicarbonates en CO2 ou l'inverse, selon les besoins du corps. Ainsi, les hématies transforment le CO2 fabriqué par les cellules en bicarbonates, puis elles vont jusqu'aux poumons, où elles retransforment le bicarbonate en CO2.
- Le transport de complexes immuns grâce au CD20, une molécule présente à la surface des hématies, qui fixe les complexes immuns et permet de les déplacer. Mais ceci est une arme à double tranchant, car en cas d'excès de complexes immuns dans le sang (par exemple au cours d'un lupus érythémateux systémique), les hématies déposent des complexes immuns dans le rein, ce qui aggrave les lésions rénales lors des lupus.
- Chez l'humain, elles portent à leur surface les antigènes des groupes sanguins érythrocytaires ABO, rhésus, Kell, Duffy[3], entre autres.
- Des études sur les érythrocytes nucléés d'espèces non mammifères révèlent leur rôle actif dans la réponse antivirale. De même les érythrocytes énucléés des mammifères sont également impliqués dans la réponse antivirale, notamment grâce à l'abondance considérable de glycophorines-A à leur surface, qui leur confèrent un potentiel rôle de "leurre" envers les protéines virales de surface, tel un piège à virions[4],[5].
Anomalie
Les hématies peuvent faire l'objet d'anomalies quantitatives : anémie dans un cas, polyglobulie dans l'autre.
Les hématies peuvent être malformées à la suite d'une déficience génétique ou beaucoup plus souvent, à une autre cause :
- Anomalies globales
- Acanthocytes
- Anisochromie
- Anisocytose
- Annulocytes
- Codocytes (Cellule-cible, en anglais : Target cells)
- Dacryocytose
- Drépanocytose (Drépanocytes ou « cellules en faucille » ou « cellules en feuille de houx »)
- Elliptocytes
- Fragmentocytes ou schizocytes (synonyme)
- Hypochromie
- Macrocytose
- Mégalocytose
- Microcytose
- Ovalocytose
- Poïkilocytose
- Polychromatophilie
- Rouleau formation
- Schistocytes
- Schizocytes, ou fragmentocytes (synonyme)
- Sidérocytes
- Sphérocytose
- Stomatocytes
- Anomalies internes (inclusions dans les globules rouges)
- Réticulocyte (caractérise des hématies jeunes dont la proportion dans la circulation est appelée réticulocytose)
- Corps de Howell-Jolly
- Anneaux de Cabot
- Corps de Heinz
- Ponctuations basophiles
- Sidérocytes
Bibliographie
- (en) Yoshikawa, Haruhisa & Rapoport, Samuel (edited by), Cellular and molecular biology of erythrocytes, Baltimore/Tokyo, University Park Press/ University of Tokyo Press, 1974.
- (en) Sporn MB, Dingman CW. Histone and DNA in isolated nuclei from chicken brain, liver, and erythrocytes. Science. 1963 Apr 19;140:316-8.
- (fr) Bader-Meunier B, Cynober T, Tchernia G (2001) Maladies de la membrane du globule rouge ; MTP, Volume 4, numéro 3, mai-
Notes et références
- (en) E. Evans et Y.-C. Fung, « Improved Measurements of the Erythrocyte Geometry », Microvascular Research, vol. 4, , p. 335-347 (DOI 10.1016/0026-2862(72)90069-6)
- Développement embryonnaire du système hématopoïétique.
- Harrisson, Principes de Médecine Interne (ISBN 2-257-17549-2).
- (en) « Is there a direct role for erythrocytes in the immune response? ».
- (en) « Evolutionary considerations in relating oligosaccharide diversity to biological function ».
 Portail de l’hématologie
Portail de l’hématologie