Mitose
La mitose, du grec mitos qui signifie « filament » (référence à l'aspect des chromosomes en microscopie), désigne les événements chromosomiques de la division cellulaire des eucaryotes. Il s'agit d'une duplication non sexuée/asexuée (contrairement à la méiose). C'est la division d'une cellule mère en deux cellules filles strictement identiques génétiquement.


Elle désigne aussi une étape bien particulière du cycle de vie des cellules eucaryotes, dit « cycle cellulaire », qui est l'étape de duplication de chaque chromosome de la cellule mère et de leur répartition égale dans chacune des deux cellules filles. Ainsi, chaque « noyau fils » reçoit une copie complète du génome de l'organisme « mère ». L’acide désoxyribonucléique (ADN) est répliqué grâce à l'ADN polymérase, lorsqu’il se trouve sous forme de chromatine (équivalent à un chromosome déroulé), lors de l’interphase du cycle cellulaire.
Le cycle cellulaire est divisé en plusieurs phases :
- La phase G1, première phase de croissance (la plus longue), appelé intervalle de temps ou Gap
- La phase S durant laquelle le matériel génétique est répliqué,
- La phase G2, qui est la seconde phase de croissance cellulaire
- La phase M, celle de la mitose proprement dite,
La phase G0 ou dite de quiescence correspond à la sortie du cycle , celle-ci survient généralement à la place G1 et ne fait pas partie du cycle cellulaire.
Les phases G1, S et G2 constituent l'interphase.
Les mécanismes de la mitose sont très semblables chez la plupart des eucaryotes, avec seulement quelques variations mineures. Les procaryotes sont dépourvus de noyau et ne possèdent qu'un seul chromosome sans centromère, ils ne se divisent donc pas à proprement parler par mitose, mais par scission binaire, tertiaire, multiple, ou par bourgeonnement.
Une cellule normale est « programmée » pour se diviser un nombre limité de fois, soit entre 50 et 70 fois[1], nombre appelé limite de Hayflick.
Les phases de la mitose
La mitose est un phénomène continu, mais pour faciliter la compréhension de son déroulement les biologistes ont décrit cinq étapes caractéristiques de la mitose qui sont la prophase, la prométaphase, la métaphase, l'anaphase et la télophase. La mitose dure entre 1 et 4 heures.
Certains auteurs considèrent la prométaphase comme une partie de la prophase plutôt qu'une phase distincte.
Prophase

L'ADN s'organise en filaments de plus en plus nets. Lors de cette phase, le matériel génétique (ADN), qui en temps normal est présent dans le noyau sous la forme de chromatine se condense en structures très ordonnées et individualisées appelées chromosomes. En effet, des protéines appelées histone H1 sont attachées de part en part sur l'ADN. Or, durant la prophase, ces histones H1 sont phosphorylées (par le MPF) ce qui provoque un enroulement accru de l'ADN qui semble se « condenser ». Le nucléole se désagrège. Comme le matériel génétique a été dupliqué avant le début de la mitose, il y aura deux copies identiques du génotype dans chaque cellule. Pendant cette phase, les chromosomes sont donc constitués de deux chromatides sœurs portant toutes les deux la même information génétique. Elles contiennent également chacune un élément d'ADN appelé centromère qui joue un rôle important dans la ségrégation des chromosomes. Les deux chromatides d'un même chromosome sont reliées au niveau de la région centromérique. Une protéine nommée cohésine joue le rôle de colle et unit les deux chromatides d'un même chromosome.
Le deuxième organite important de la prophase est le centrosome, composé initialement de deux centrioles pour les cellules animales (sauf celles ayant perdu leur capacité mitotique, comme la majeure partie des cellules nerveuses qui n'en contiennent aucun, et les cellules cancéreuses qui peuvent en contenir davantage ; les centrosomes des cellules végétales n'en contiennent qu'un seul). Comme pour les chromosomes, le centrosome s'est dupliqué avant le début de la prophase (en 4 centrioles). Les 4 centrioles se séparent durant la prophase, formant deux centrosomes qui migrent chacun vers un pôle de la cellule. Le cytosquelette de microtubules se réorganise pour former le fuseau mitotique, structure bipolaire qui s'étend entre les deux centrosomes mais reste à l'extérieur du noyau. Par la croissance des microtubules, le fuseau mitotique s'allonge, ce qui étire le noyau cellulaire.
On peut se représenter les microtubules comme des perches ou des rails, dans la cellule. Certaines cellules eucaryotes, notamment les cellules végétales, sont dépourvues de centriole.
Prométaphase
Certains auteurs considèrent la prométaphase comme une partie de la prophase, plutôt que comme une phase distincte. De plus, elle ne se produit pas chez toutes les cellules eucaryotes.
Durant la prométaphase, l'enveloppe nucléaire se désagrège sous forme de vésicules, initiant ainsi la mitose ouverte. L'enveloppe nucléaire se reformera en fin de mitose. (Chez certains protistes, l'enveloppe nucléaire reste intacte. On assiste alors à une mitose fermée).
Des complexes protéiques spécialisés : les kinétochores, se forment au niveau des centromères. Certains microtubules s'accrochent aux kinétochores. Ils seront alors appelés microtubules kinétochoriens. Les microtubules accrochés seulement aux centrosomes sont appelés microtubules polaires. Les microtubules qui ne font pas partie du fuseau mitotique forment l'aster et sont appelés microtubules astraux. Petit à petit chaque chromosome voit chacune de ses chromatides reliée à un pôle par l'intermédiaire des microtubules. Ceux-ci exerçant des tensions, les chromosomes ont alors des mouvements agités.
Métaphase

Deuxième phase de la mitose, après la prophase, c'est le rassemblement des chromosomes condensés à l'équateur de la cellule, pour former la plaque métaphasique (ou équatoriale) . Les tensions subies par chacun des kinétochores d'un chromosome s'équilibrent progressivement et ceux-ci s'alignent dans un plan situé à mi-chemin des deux pôles. On observe que les chromosomes sont alignés selon leur centromère. On pense que les kinétochores non accrochés aux microtubules génèrent un signal pour empêcher l'étape prématurée de l'anaphase sans les chromosomes tous alignés. Ce signal crée le point de contrôle du fuseau mitotique (en)[2].

Anaphase

L'anaphase est une phase très rapide de la méiose et de la mitose où les chromatides se séparent et migrent vers les pôles opposés de la cellule. Les fils chromosomiques sur lesquels étaient accrochés les centromères des cellules se détachent et les chromatides se déplacent chacune vers un pôle de la cellule.
Durant cette phase, à la suite d'un signal spécifique qui correspond à une augmentation d'un facteur 10 de la concentration en calcium intracellulaire et à l'inactivation du MPF (protéolyse de la cycline B du MPF), les chromatides sœurs se séparent brutalement. Elles sont alors « tirées » par les microtubules en direction du pôle auquel elles sont rattachées. Les chromatides migrent rapidement à une vitesse d'environ 1 µm/min. Il y a deux catégories de déplacements : l'anaphase A et l'anaphase B.

Pendant l'anaphase A, les chromatides, en réalité, se déplacent en direction du pôle sur les microtubules kinétochoriens qui raccourcissent, car ils se dépolymérisent par leur extrémité + au fur et à mesure de la progression du kinétochore. En effet, les kinétochores permettent non seulement d'« arrimer » une chromatide au microtubule, mais aussi de les faire transporter le long des microtubules. Au niveau des kinétochores on trouve des « moteurs » moléculaires (de type dynéine) utilisant de l'ATP qui permettent de tracter les chromatides le long des microtubules qui eux, restent fixes.
Pendant l'anaphase B, les microtubules polaires s'allongent, et les pôles du fuseau mitotique s'éloignent l'un de l'autre entraînant avec eux les chromatides.
Télophase
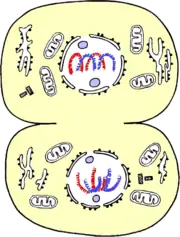
Le terme « télophase » dérive du grec « telos » signifiant « fin ». C'est la 4e phase de la mitose.
Durant cette période :
- les microtubules polaires vont persister au niveau de leur extrémité + pour former les microtubules interzonaux qui disparaîtront lors de la phase la plus terminale de la télophase, la cytodiérèse, qui correspond à la division terminale des deux cellules filles.
- Les microtubules kinétochoriens disparaissent.
- Les chromatides sœurs commencent à se décondenser.
- L'enveloppe nucléaire ainsi que les nucléoles commencent à se reformer dans la métabiose.
Cytodiérèse
Appelée encore cytokinèse ou cytocinèse, la cytodiérèse fait partie de la télophase (même si certains auteurs la considèrent extérieure à la mitose), et son mécanisme n'est pas le même dans les cellules animales et végétales.
Dans les cellules animales, le sillon de division se forme dans un plan perpendiculaire à l'axe du fuseau mitotique et sépare la cellule en deux. Il peut en effet commencer à se former dès l'anaphase. Le clivage est dû à un anneau contractile qui est composé principalement d'actine et de myosine (myosine II). L'anneau contractile comporte des filaments d'actine de 5 à 7 nm de diamètre arrangés concentriquement. La myosine et l'α-actinine sont mises en évidence par immunofluorescence.
Ce sont des interactions entre actine et myosine de l'anneau contractile qui sont à l'origine des forces provoquant la contraction de l'anneau et le partage du cytoplasme parental par étranglement. Cette constriction se fait de manière centripète. Le sillon de division se resserre jusqu'à former un corps intermédiaire, formant un passage étroit entre les deux cellules filles et qui contient le reste du fuseau mitotique. Celui-ci finira par disparaître entièrement et les deux cellules filles se sépareront complètement. Par ailleurs, l'enveloppe nucléaire et les nucléoles finissent de se reconstituer et l'arrangement radial interphasique des microtubules nucléés par le centrosome se reforme.
À la fin de la cytocinèse, les deux cellules filles ne sont plus reliées que par un pont au milieu duquel une excroissance sphérique, le corps central, détermine le futur des cellules filles. La cellule qui en hérite va continuer à se diviser (elle demeure une cellule souche), tandis que l'autre va se différencier en un type précis de cellule[3].
Chez la cellule végétale, la cytodiérèse est très différente de par la présence d'une paroi rigide (divisée en une paroi primaire, cellulosique, et une paroi primitive, pectinique, l'ensemble formant une paroi pecto-cellulosique). Elle se réalise par la construction d'une nouvelle paroi, phragmoplaste appelé plus simplement corps intermédiaire entre les deux cellules filles. Cette nouvelle paroi se développe de manière centrifuge : des vésicules golgiennes contenant de la propectine s'accumulent du centre de la cellule vers la périphérie, puis ces vésicules fusionnent pour former le phragmoplaste qui se raccorde à la paroi primaire de la cellule mère, provoquant sa division en deux cellules filles. La paroi primaire et la membrane des deux cellules filles se reforment alors au niveau de cette séparation et le phragmoplaste se différencie en lamelle moyenne, ou paroi primitive.
Conséquences des erreurs
Des erreurs peuvent survenir lors de la formation des nouvelles cellules : le processus peut en effet mal se dérouler (il est à noter que l'organisme humain adulte est constitué de quelques dizaines de milliers de milliards de cellules et que chaque jour un milliard de cellules doivent être renouvelées par division cellulaire[4]). Et lorsque ces erreurs mitotiques surviennent pendant les premières divisions cellulaires d'un zygote, elles peuvent avoir des conséquences particulièrement néfastes.
Exemples d'erreurs mitotiques :
- Phénomène de non-disjonction : les deux chromatides d'un chromosome ne se séparent pas pendant l'anaphase. Une cellule fille recevra un chromosome à deux chromatides et l'autre n'en recevra aucun. Une des cellules filles aura alors une trisomie et l'autre une monosomie, qui sont des cas d'aneuploïdie.
- Délétion, translocation, inversion, duplication chromosomiale :
La mitose est un processus traumatique. La cellule subit des changements importants dans son ultrastructure, ses organites se désintègrent et se reforment plusieurs heures après, et les chromosomes sont constamment déplacés par les microtubules. Occasionnellement, les chromosomes peuvent être endommagés. Un bras du chromosome peut être cassé et le fragment est alors perdu, causant une délétion. Le fragment peut être incorrectement rattaché à un autre chromosome non homologue, ce qui cause une translocation. Il peut être réattaché au chromosome initial, mais en sens inverse, causant une inversion. Ou encore, il peut être considéré à tort comme un chromosome séparé, causant alors une duplication chromosomale. L'effet de ces anomalies dépend de la nature spécifique de l'erreur. Parfois il n'y aura aucune conséquence, d'autre fois, cela peut induire un cancer, ou même causer la mort de l'organisme.
Pour éviter ces erreurs, de nombreux points de contrôle (en), notamment à la fin de la phase G1 ou G2, bloquent le déroulement du cycle lorsqu’une anomalie est détectée (endommagement de l’ADN, ADN non complètement répliqué, chromosomes non attachés au fuseau mitotique) : une fois la division arrêtée, un mécanisme réparant les erreurs se met en place et si ce mécanisme ne se produit pas, une mort cellulaire programmée est enclenchée[5].
Un exemple de maladie entraînée par une délétion, touchant la partie terminale du bras court du chromosome 5, est le syndrome du cri du chat. Dans cette maladie l’enfant pousse des cris semblables à des miaulements, présente une microcéphalie entraînant notamment un retard mental prononcé et possède un visage en forme lunaire caractéristique.
Méiose et mitose
La mitose et la méiose diffèrent sur un certain nombre de points, mais présentent également des similitudes (mécanismes de séparation des chromosomes, etc.). La mitose correspond à une reproduction asexuée des cellules, alors que la méiose est un prélude à la reproduction sexuée. Par la méiose, chaque parent produit des gamètes différents et destinés à se rencontrer. De nombreux types de cellules sont capables de mitose mais seules celles des organes reproducteurs, les gonades (ovaires et testicules) réalisent la méiose. À partir d'une cellule, à la fin de la mitose il y a deux cellules génétiquement identiques alors qu'à la fin de la méiose il y a quatre cellules le plus souvent génétiquement différentes et donc uniques.
Mitose végétale
Les principales différences entre la mitose végétale et la mitose animale sont l'absence de centrioles chez les plantes (à part chez les algues et certains gamètes), la présence d'une paroi qui conduit à une cytodiérèse particulière, son rôle dans le développement postembryonnaire et sa régulation hormonale. La mitose végétale est encore mal comprise, notamment la manière dont le fuseau mitotique peut se former en l'absence de centrioles et de centrosomes (mais au niveau de chaque pôle en début de prophase on a une condensation cytoplasmique appelée calotte polaire qui émet des rayonnements qui vont former en fin de prophase le fuseau mitotique). Donc la différence est que pour la cellule animale pendant la mitose au niveau des pôles on a les asters provenant des centrioles et pour la cellule végétale on a les calottes polaires provenant de la condensation du cytoplasme. Néanmoins, les événements de mitose sont fortement liés aux réarrangements du cytosquelette.
Cytodiérèse : La séparation des cellules filles se produit par formation d'une nouvelle paroi pectocellulosique sur le plan équatorial de la cellule. Ce plan est déterminé par la localisation de certaines protéines dès le début de la mitose. À la fin de la télophase des microtubules forment une plaque au niveau équatorial, c'est le phragmoplaste. Des vésicules de membranes provenant de l'appareil de Golgi et des précurseurs des composants de la paroi viennent s'y associer.
Rôle dans le développement : Chez les organismes unicellulaires la disponibilité des nutriments dans le milieu est le facteur régulateur principal de la mitose qui dépend en fait de la taille de la cellule. Chez les organismes pluricellulaires les divisions se produisent uniquement dans les méristèmes, et les cellules méristématiques dépendent pour la régulation de leur cycle cellulaire (comme pour leur approvisionnement en nutriments) des signaux générés par les cellules somatiques (en phase G0, c'est-à-dire quiescentes, qui ne se divisent pas) : il s’agit d’un contrôle social. La formation des tissus et des organes ne se produit qu’au niveau de méristèmes par accumulation de cellules (mérèse).
La mérèse n’ayant lieu que dans les méristèmes, si une cellule somatique est endommagée ou détruite elle n’est pas remplacée, contrairement à ce qui se passe dans le règne animal. Ce qui fait que les plantes n’ont pas un plan d’organisation aussi strict que celui des animaux, il y a formation de nouveaux organes et sénescence des anciens. Autre différence, chez les plantes l’apoptose est peu importante dans la formation des organes.
Régulation hormonale : Le signal de différenciation est donné aux cellules immatures par les cellules matures. Les signaux peuvent être des hormones non-peptidiques (auxine, cytokinines, éthylène, acide abscissique, brassinostéroïdes), des lipo-oligosaccharides (facteur nod), des peptides (systémine). La réponse aux hormones est variable selon les tissus. Elle intervient via les gènes MAPK (cascades kinases MAPK), déclenchant l’accumulation de cyclines nécessaire à l’entrée en phase S.
L’auxine et les cytokinines jouent de concert un rôle majeur dans la mitose. L’apport exogène d’auxine est nécessaire aux méristèmes qui peuvent être autosuffisants en cytokinines. Si une des deux hormones est absente aux niveaux suffisants la mitose n’a pas lieu. L’auxine active l’expression des gènes SAUR (réponse 2-5 min) et AUX/IAA (réponse 5-60 min). Elle agit surtout sur les méristèmes secondaires (principalement le cambium). Les cytokinines stimulent la séparation des chromosomes et la cytokinèse, provoquent l’accumulation de cyclines et activent la phosphatase cdc25 qui active la cycline kinase cdc2 par déphosphorylation de la tyrosine 15. Elles sont nécessaires à l’initiation du cycle cellulaire comme à sa progression.
L’ABA inhibe la mitose en réponse au stress hydrique en induisant la synthèse d’ICK, inhibiteur de cdk-cycline, dans les tissus méristématiques. Les brassinostéroïdes et les gibbérellines favorisent la mitose. Les gibbérellines stimulent la prolifération des méristèmes intercalaires (monocotylédones) et des tissus corticaux et épidermiques, insensibles à l’auxine en augmentant l’expression de l’histone H3 et de la cycline 1.
Le facteur Nod déclenche la nodulation racinaire en présence de bactérie Rhizobium.
En réponse à un stress une plante diminue la croissance de ses organes en ralentissant le cycle cellulaire ce qui réduit le taux de mitose et la taille finale des nouveaux organes (ils contiennent moins de cellules). Cet effet est plus important dans les racines que dans les feuilles. La réponse aux stress hydriques et salins a lieu par l’intermédiaire de l’ABA qui augmente l’expression de ICK1 qui interagit avec CDKA et inhibe l’activité histone H1 kinase. De plus, la cycline kinase cdc2 est désactivée par phosphorylation (la phosphorylation de cdc2 est considérée comme un élément majeur de la réduction de la division cellulaire en réponse au stress). Un autre messager de stress est le jasmonate, impliqué dans la réponse aux blessures, aux pathogènes et la synthèse des parois végétales qui neutralise l’activité des cytokinines et inhibe la division cellulaire. La sensibilité des cellules au jasmonate dépend de la phase du cycle (plus importante en G1).
Les signaux environnementaux affectent la croissance et la division cellulaire. C'est une des formes d’adaptation de la plante aux changements environnementaux. Les cellules quiescentes (G0) peuvent occasionnellement sous l’influence de facteurs hormonaux (auxine), nutritionnels ou environnementaux (lumière) repasser en phase G1 pour entreprendre un cycle de division. Ce maintien d’une capacité mitotique des cellules quiescentes permet d’atteindre les ressources environnementales (lumière et minéraux).
Maintien de trois génomes : En plus du génome du noyau et de celui des mitochondries, les plantes doivent répliquer le génome des chloroplastes. La réplication des génomes des organites n’intervient que dans les méristèmes et les organes primordiaux. Lorsque la cellule est en division rapide le nombre de génomes par organite augmente grandement. Lorsque la vitesse de division ralentit la réplication des génomes cesse et le nombre d’organites par cellule augmente par division jusqu’à ce qu’il n’y ait plus qu’un ou deux génomes par organite.
Médias
- La mitose en animation.
- [vidéo]La mitose filmée de cellule animale (puis accélérée)
- [vidéo]Mitose de cellule végétale, filmée (puis accélérée), sur YouTube.
Notes et références
- Elaine N. Marieb (trad. de l'anglais), Anatomie et physiologie humaines, Canada, Édition du Renouveau Pédagogique inc (ERPI), , 1293 p. (ISBN 978-2-7613-3071-8), p. 164.
- (en) Chan G, Yen T., « The mitotic checkpoint : a signaling pathway that allows a single unattached kinetochore to inhibit mitotic exit », Progress in Cell Cycle Research, no 5, , p. 431–439
- Kheira Bettayeb, « Division cellulaire. Le moment clé révélé », Science & vie, , p. 71 (lire en ligne)
- L. Meijer, « Le cycle de division cellulaire et sa régulation », Oncologie, no 5, , p. 311
- L. Meijer, op. cité, p. 312
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire