Bryophyte
Les bryophytes sont des plantes terrestres thalloïdes ou feuillées non vascularisées. Parmi les plantes actuelles, les bryophytes terrestres et les bryophytes aquatiques sont celles qui ont conservé le plus de caractères des premières plantes ayant colonisé la terre ferme. Les ancêtres de toutes les plantes terrestres, donc des bryophytes, sont des algues vertes charophycées.
Ne doit pas être confondu avec Bryophyta.



Même si certaines structures anatomiques leur permettent de résister à la dessiccation, les Bryophytes sont encore extrêmement dépendantes de l'eau ou des milieux humides, ou en tout cas d'une hygrométrie minimale au moment de leur reproduction. Cette exigence n'empêche une grande plasticité écologique qui leur permet de vivre dans toutes les régions du globe, depuis l'équateur aux pôles[3].
Pris au sens large, c'est-à-dire celui des classifications traditionnelles, le terme bryophyte s'applique aux trois phylums de plantes terrestres qui ne possèdent pas de vrai système vasculaire, les Marchantiophytes (hépatiques), les Bryophytes s.s. (mousse, sphaigne) et les Anthocérotophytes (anthocérotes). Au sens strict botanique, l'embranchement actuel des Bryophyta ne concerne que les mousses et les sphaignes (à l'exception donc des Hepaticophyta et des Anthocerotophyta).
Avec près de 25 000 espèces de mousses et sphaignes, 9 000 espèces d'Hépatiques (Marchantiophytes) et 300 espèces d'anthocérotes, les bryophytes constituent le second groupe de végétaux terrestres, après les Dicotylédones.
Composants importants des biotopes forestiers et montagnards, les bryophytes sont une source majeure de la biodiversité spécifique des environnements frais ou humides et contribuent de façon non négligeable à la structure et au fonctionnement des écosystèmes que ce soit en termes de dynamique végétale, de formation des sols et plus largement dans les cycles biogéochimiques et la régulation des climats[3].
Historique
Connues et utilisées depuis la nuit des temps, les bryophytes ne font l'objet d'une attention particulière que depuis la fin du XVIIIe siècle, le seul ouvrage consacré exclusivement aux mousses avant étant l'Historia muscorum de Dillenius publié en 1741[4]. Au cours des siècles précédents, les bryophytes, comme les autres cryptogames (taxon désormais non valide comprenant les algues, les lichens, les fougères), sont en effet peu étudiées compte tenu de leur petitesse, de leur organe reproducteur non visible et du peu d'avantages que l'Homme arrive à en tirer[5]. En 1784, Johannes Hedwig, « père de la bryologie », en fait un ensemble véritablement naturel, subdivisé en Musci frondosi (Mousses) et Musci hepatici (Hépatiques)[6]. En 1789, Jussieu propose une classification naturelle dans son Genera plantarum, et est le premier à utiliser le terme de mousse pour représenter cet ensemble qu'il classe dans les « plantes sans fleurs » avec les Fungi (champignons), Algae (algues) et Filices (fougères)[5]. Le terme Bryophyte est inventé en 1864 par le botaniste allemand Alexander Braun qui accole deux mots grecs, bryo signifiant mousse et phytos qui signifie plante[7].
Les classifications phylogénétiques récentes intègrent les bryophytes dans le taxon monophylétique des Embryophytes, plantes terrestres comprenant les « cryptogames » (Bryophytes et Ptéridophytes)[5]. La systématique phylogénétique suggère également qu'elles sont un groupe paraphylétique (la monophylie ou la polyphylie de ce taxon a longtemps été discutée[8]), abandonné par les cladistes mais conservé comme grade évolutif par les systématiciens évolutionnistes. Il comporte trois phylums, les Marchantiophytes (hépatiques), les Bryophytes s.s. (mousse, sphaigne) et les Anthocérotophytes (anthocérotes)[9].
Cycle de vie des bryophytes
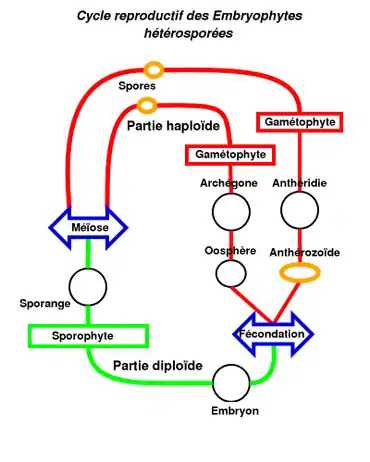





Comme pour les algues vertes et les plantes vertes, le cycle de vie présente une alternance de sporophytes (ou Sporogone) et de gamétophytes, mais, comme pour toutes les Embryophytes, les anthérozoïdes (ou spermatozoïdes) sont protégés (et formés) par des anthéridies et les oosphères par des archégones (caractères partagés avec les plantes vasculaires). Ces structures fertiles en forme de corbeilles renferment ainsi les gamétanges qui sont généralement séparés par des poils stériles (appelés paraphyses) qui jouent un rôle protecteur contre la dessiccation en favorisant la rétention d'eau par capillarité[11].
La fécondation est une zoïdogamie aquatique (oogamie), les anthérozoïdes (ou spermatozoïdes) ciliés, libérés de l'anthéridie par gélification et rupture de la paroi, sont entraînés par les gouttes d'eau de pluie ou de rosée, retenue par les feuilles ou les thalles (ou par les éclaboussures de gouttes de pluie lorsque les mousses sont dioïques) dans lesquelles ils se déplacent pour atteindre l'oosphère (la distance maximale de dispersion des spermatozoïdes étant d'une dizaine de centimètres)[12]. L'embryon qui en résulte est nourri et protégé par le gamétophyte (matrotrophie). Le sporophyte non ramifié fixé sur le gamétophyte ne devient jamais indépendant de celui-ci. Le gamétophyte haploïde est donc le stade dominant sous lequel se rencontre la plante. Il peut être monoïque (plante bisexuée) ou dioïque (plante monosexuée). Lorsqu'il est monoïque, une protandrie (maturation de l'anthéridie avancée par rapport à celle de l'archégone) permet généralement d'éviter l'autofécondation[13].
Les spores sont entourées d'une paroi contenant de la sporopollénine qui les protège de la dessication au cours de la dissémination[14]. Les botanistes parlent de digénétique haplodiplophasique (présence de 2 générations séparées, le gamétophyte haploïde et le sporophyte diploïde) qui devient de plus en plus hétéromorphe, avec prédominance du gamétophyte sur le sporophyte (sporogone)[15].
Généralement, seule une oosphère remplie de réserves et fixée dans son réceptacle est fécondée et conduit à la production d'un sporophyte (monosétie). Il arrive que plusieurs oosphères d'un même réceptacle soient fécondées aboutissant à la production de plusieurs sporophytes. Ce phénomène est appelé polysétie[16].
Dans certains cas, en particulier chez les anthocérotes, les gamétophytes sont bisexués et nourrissent alors de nombreux sporophytes.
- Après la fécondation, l'embryon matotrophe est porté par le gamétophyte femelle et donne un sporophyte souvent formé d'une soie et d'un sporange (en vert). Le sporange libère des spores mâles et femelles (sauf dans le cas des gamétophytes bisexués) qui sont dispersés principalement par le vent (parfois par les mouches chez les Splachnaceae (en)[1]). La plupart des spores tombent au sein de la colonie parentale avant de germiner sous forme de protonema, après une phase de croissance et de développement, donne les gamétophytes[17].
- Les stades mobiles (en jaune dans le schéma) sont principalement les spores pour lesquelles les mousses ont développé des mécanismes assurant leur dispersion (capsule explosive des sphaignes, fentes de déhiscence des andréales, péristome des vraies mousses) et les anthérozoïdes (ou spermatozoïdes) avant la fécondation.
Les bryophytes peuvent se multiplier de façon végétative par fragmentation (bouturage naturel). Certaines hépatiques ont développé des structures de dispersion, les propagules. Des structures particulières (gemmules, bulbilles) existent aussi chez certaines mousses[18]. Tous ces éléments sont principalement dispersés par le vent, mais il existe des vecteurs animaux, tels que les chauves-souris[19], les fourmis[20], les limaces[21]. L'allocation des ressources (correspondant à l'allocation de nutriments) reflète l'existence de compromis évolutifs entre deux traits biologiques, la reproduction sexuée favorisée pendant les stades de colonisation et la reproduction asexuée favorisée au sein des colonies matures[22].
Organisation fonctionnelle
Pas de système vasculaire ou un système vasculaire rudimentaire
Les Bryophytes ne disposent pas de xylème ni de phloème (faisceaux conducteurs de l'eau et des nutriments). Ces deux tissus ne sont apparus que plus tard au cours de l'évolution, chez les plantes vasculaires. L'irrigation de l'axe feuillé (appelé caulidie) se réalise par un processus de capillarité, possible grâce à l’étroit agencement des « feuilles » sur cet axe. Certains bryophytes disposent cependant de tissus conducteurs spécialisés permettant le transport de l'eau, des sels minéraux et des produits de la photosynthèse, mais ces derniers ne sont pas lignifiés : les « leptoïdes » (cellules vivantes allongées assurant la distribution des produits de la photosynthèse, l'équivalent de la sève élaborée) et les « hydroïdes » (cellules mortes allongées assurant la distribution de l'eau et des sels minéraux, l'équivalent de la sève brute)[23]. Les mousses endohydriques ont un tissu conducteur développé à la périphérie de l'axe feuillé, les mousses ectohydriques ont seulement des jonctions protoplasmiques entre les cellules, ce qui permet une circulation très localisée des nutriments[24]. Les mousses possèdent cependant des tannins complexes proches de la lignine, les lignanes (biopolymères présents dans les vacuoles et les parois cellulaires). Leur appartenance aux métabolites secondaires ainsi que les caractéristiques toxiques, antifongiques ou antibactériennes de certains lignanes font émettre l'hypothèse de leur implication dans les défenses chimiques des plantes contre les herbivores et la photoprotection[25].
Les pseudophylles ou phyllidies n'ont pas l'histologie des vraies feuilles.
Il n'existe pas d'organes comparables aux racines chez les bryophytes dotés de « rhizoïdes » (mono ou pluricellulaires) qui les fixent au substrat mais n'absorbent pas la solution du sol. Les bryophytes ont un cormus (« tige » et « feuille », ces deux organes donnant un axe feuillé). En l'absence de photosynthèse, ce cormus se nécrose, se décolore et forme ainsi un humus primitif.
Les bryophytes ne pouvant absorber l'eau par leur rhizoïdes ont la faculté de permettre à l'eau de rentrer par toutes les parties de la plante. De plus en cas de forte sécheresse la plante peut entrer en état de vie ralentie (reviviscence) en attendant des conditions de développement mieux adaptées.
 Coupe transversale de l'axe feuillé de Dawsonia[26].
Coupe transversale de l'axe feuillé de Dawsonia[26]. Coupe transversale de la « feuille » du Polytric commun[27].
Coupe transversale de la « feuille » du Polytric commun[27].
Symbioses avec des champignons
Comme les plantes vascularisées, les bryophytes peuvent profiter d'une symbiose avec des champignons. On a encore peu de connaissances sur la diversité fongique associée aux bryophytes. Mais quelques études basées sur la biologie moléculaire ont montré qu'elle était importante.
Par exemple, en forêt boréale les bryophytes se montrent très résistantes au froid et capables de se nourrir sur des sols gelés tout ou partie de l'année. Les champignons les y aident. Il existe au nord du Canada « un ensemble écologiquement et phylogénétiquement diversifié de champignons associés aux parties vivantes de 3 bryophytes abondantes et largement distribuées (...),les Hylocomium splendens (Hedw.) Schimp. in B.S.G., Pleurozium schreberi (Brid.) Mitt. et Polytrichum commune Hedw »[28] ; Au moins 158 espèces de champignons leur sont associées dans la région étudiée. Ces champignons appartiennent surtout à l'embranchement des Ascomycota (62,8 % des cas), et à la subdivision des Basidiomycota (32 % des cas), parfois des Chytridiomycota (considérés comme les ancêtres de tous les autres champignons (3,9 % des cas) et rarement aux Glomeromycota (1,3 % seulement des cas, bien que ces champignons soient très fréquemment associés aux plantes supérieures)[28]. Les principaux ordres concernés étaient dans cette région les Hélotiales (18,6 %), les Agaricales (11,5 %), les Chaetothyriales (9,6 %) et les Tremellales[28]. L'association la plus souvent repérée était l'agaric Entoloma conferendum avec Lophodermium piceae (Fckl.) Hoehn. comme endophyte associé[28]. D'autres taxons avaient des affinités avec des Hélotiales du genre Hyphodiscus ou avec plusieurs basidiomycètes mycorhiziens[28]. La plupart (72,2 %) des taxons de champignons endophytes n'ont été trouvés qu'une seule fois dans cette étude[28]. De plus, très peu de champignons étaient associés avec l'ensemble des 3 bryophytes étudiés, ce qui indique une diversité d'associations très large et peut-être certaines spécificités[28], qui restent à explorer.
Autre exemple, la Pleurozie dorée est une mousse qui peut fournir du phosphate et du carbone aux racines du Pin tordu colonisées par le bolet des bouviers qui forme des ectomycorhizes avec eux[29].
Habitats et répartition
Capables de tirer leurs nutriments de l'air et des aérosols, comme les lichens qu'ils accompagnent souvent, les bryophytes sont en tant que groupe taxonomique très ubiquistes. Grâce à de nombreuses adaptations, ils ont colonisé presque toute la planète, jusqu'aux zones subpolaires[30]. Ils sont toutefois absents (plus que les lichens) du milieu marin, et des milieux extrêmement arides. Grâce à des spores aéroportées, ils comptent parmi les premières espèces à pouvoir coloniser les nouveaux habitats (ex : terrils, îles volcaniques nouvelles...)
De nombreuses espèces vivent sur le sol (terricoles), dans l'humus (humicole) ou les milieux tourbeux (turficole), d'autres sont saxicoles (ou devenues muricoles) ou épiphytes, ou encore se développent sur les bois-morts tombés au sol (corticoles sur l'écorce). Dans les réserves naturelles et en forêt dans les îlots de sénescence, après une vingtaine d'années de non-gestion (« restauration passive »[31]), le taux de bois mort au sol ré-augmente au profit des bryophytes notamment[31] mais aussi de nombreux cortèges saproxylophages, chacun caractéristique de l'un des stades de décomposition du bois, et du type d'essences (bois dur, bois tendres, etc) ou de situation (à l'ombre, au soleil, dans l'eau...)[32].
 Hypne cyprès, espèce ubiquiste.
Hypne cyprès, espèce ubiquiste. Riccia fluitans, hépatique flottante utilisée en aquariophilie pour l'oxygénation de l'eau.
Riccia fluitans, hépatique flottante utilisée en aquariophilie pour l'oxygénation de l'eau.

 Mousse des jardiniers, espèce humicole.
Mousse des jardiniers, espèce humicole.
Écologie

Turgescentes, les mousses se gorgent d'eau, un kilogramme de bryophyte sèche pouvant retenir 15 litres d'eau (les capacités d'absorption étant maximisées chez les sphaignes dont un kg peut retenir 70 à 75 litres d'eau)[35].
Les Bryophytes sont généralement autotrophes par photosynthèse grâce aux cellules pourvues de chlorophylle. Certaines en sont dépourvues, telle l'hépatique Cryptothallus mirabilis qui vit sous d'autres mousses en milieu bois. Parasite sur un champignon du genre Tulasnella (lequel forme une ectomycorhize avec des arbres du genre Betula ou Pinus), elle est l'unique hépatique non chlorophyllienne d'Europe et reste associée à des milieux naturels de très grande qualité[36].
Dans une certaine mesure, en empêchant physiquement des graines d'accéder au sol ou en limitant l'accès de la lumière aux graines déjà présentes dans les sols (spectre du rouge notamment, qui déclenche la germination de certaines graines) et/ou par allélopathie, les tapis de mousses peuvent éliminer ou sélectionner certaines espèces[37] ; par exemple dans une pelouse calcaire néerlandaise (et de même en conditions contrôlées en serre), un tapis dense de mousses sur le sol réduit jusqu'à 30 % le nombre de plantules de certaines espèces phanérogames[37].
Les mousses ont un pouvoir absorbant et isolant mis à profit par les rongeurs qui utilisent ce matériau pour tapisser leurs terriers ou par les oiseaux pour leurs nids. Lors de relevés bryophytiques, les bryologues ont distingué 65 espèces de mousses qui garnissaient le nid de passereaux comme les mésanges. Plus de la moitié de la masse de leurs nids est composé de mousses[38].
Malgré le pouvoir nutritif des mousses identique aux plantes vasculaires et le fait qu'elles offrent un habitat à de nombreux invertébrés, elles sont très peu consommées par les herbivores, notamment en raison de la synthèse de composés peu digestes voire inappétents (tanins, terpénoïdes) qui interviennent probablement dans des réactions de défense contre les herbivores[39]. Elles servent de nourriture à quelques espèces de coléoptères, orthoptères, collemboles, chenilles ou pucerons[40] Elles sont plus consommées dans les régions de haute latitude où elles dominent la végétation (zones arctiques et toundras) où elles font partie du régime alimentaire hivernal des ruminants comme le renne et le bœuf musqué[41].
Services écosystémiques

Les mousses fournissent de nombreux services écosystémiques.
Espèces ingénieurs, elles ont la capacité, en s'installant, de créer des micro-habitats qui hébergent une diversité faunistique (un mètre carré de mousses abrite des milliers d'espèces de vers, acariens, rotifères, collemboles, tardigrades, pseudoscorpions, et une entomofaune riche de coléoptères, fourmis) et floristique (lichen, épiphytes telle que les Hyménophyllacées et les orchidées) importantes[43]. Une seule mousse peut abriter des dizaines d'espèces d'invertébrés[44]. Les mousses servent également de site de reproduction à des insectes et acariens[45].
Elles ont pris une place dans le quotidien des hommes (serviettes hygiéniques en sphaignes, couches, papier toilette, carburant en Europe, costumes traditionnels en Nouvelle Guinée), en médecine (utilisées en Chine pour leurs vertus médicinales), en agriculture (engrais, phytosanitaire)[46], dans leur foyer (remplissage de matelas ou d'oreiller, calfeutrage des habitations en bois, toits végétalisés pour l'isolation, plantes ornementales en aquariophilie) ou dans la construction navale (calfatage des embarcations)[47],[48]. Les chercheurs ont trouvé chez les bryophytes des antibiotiques, des antifongiques, des hormones végétales, des anti-tumoraux[49].
Bryophyte, bioindication, environnement et pollution...
Les bryophytes, et les mousses en particulier tirent l'essentiel de leur nourriture de l'air et des nutriments apportés par l'eau[50], par la pluie ou les dépôts de particules et aérosols. En raison notamment de leurs grandes capacités d'absorption d'eau et de minéraux passivement, et de leur tissu épidermique constitué le plus souvent d'une seule couche de cellules et exempt de cuticule (ce qui permet d'accroître la capacité d'échange), de nombreux bryophytes sont vulnérables à l'acidification de l'air, à sa pollution[51] et à l'eutrophisation des milieux[52].
Ils jouent à ce titre un rôle important dans l'épuration de l'air, mais aussi en matière d'accumulation de certains polluants (métaux lourds et radionucléides notamment). Ce sont aussi des organismes pionniers de succession (capables de croître sur des sols squelettiques et de fournir de l'humus pour les communautés suivantes) importants dans les processus de résilience écologique.
Ils sont moins sensibles à la continuité temporelle de l'état forestier que les lichens[53]
Les espèces les plus sensibles aux polluants acides et aux pesticides véhiculés par l'air et les pesticides ont parfois disparu de tout ou partie de leur habitat naturel. Certaines espèces pourraient être considérées comme des bioindicateurs, en matière de qualité thermohygrométrique de l'air notamment. De par leur résistance naturelle à de nombreux polluants elles ne font pas de bons bioindicateurs de pollution, mais comme bioaccumulateurs, elles peuvent permettre de cartographier la pollution présente et d'un passé récent. On les a utilisées par exemple pour cartographier des pollutions par le plomb ou le cadmium. Orthotrichum diaphanum (de) est une mousse épiphyte qui vit sur les murs et les trottoirs humides des villes et qui résiste à la pollution urbaine[54].
Ils ont aussi été utilisés comme indicateurs de changement climatique récent[55].
Histoire évolutive
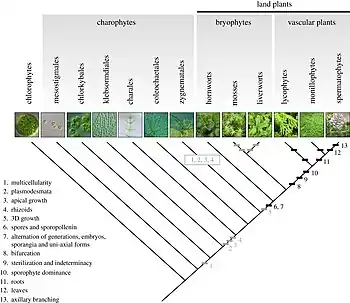
_4293.jpg.webp)
Le fossile de mousse Pallavicinites devonicus, daté du dévonien supérieur (environ 350 millions d'années)[57], rappelle que les embryophytes (i.e. les plantes terrestres) sorties des eaux il y a environ 450 millions d’années sont probablement issues d'algues vertes du groupe des charophytes, qui ont évolué en eau douce et sont capables de supporter des émersions temporaires grâce à ces associations symbiotiques (bactéries, micro-algues et micro-champignons)[58].
Les premiers phylums à émerger au sein des bryophytes sont respectivement et successivement les hépatiques à thalle, les anthocérotes et les mousses vraies[59].
La conquête du milieu terrestre exige un certain nombre d'adaptations qui sont toujours présentes chez les mousses actuelles. Faisant partie des premières plantes terrestres pour qui l'eau et le gaz étaient les facteurs limitants, elles avaient certainement peu de possibilités de contrôler leur teneur en eau (absence de cuticule et de stomates). Ne possédant ni de vrais vaisseaux conducteurs, ni de véritables racines, ni d'organes de stockage, elles ont développé une grande capacité d'absorption de l'eau (elles fonctionnant pour la plupart comme une éponge), une tolérance à la dessiccation et une capacité de reviviscence[60], ce qui explique que les bryophytes se sont autant affranchies du milieu aquatique que les végétaux vasculaires, beaucoup de mousses survivant dans des milieux secs (par exemple les espèces pionnières capables de coloniser les roches nues, les déserts chauds et froids[61]les toits ensoleillés des maisons)[62]. Elles ont toujours besoin d'eau (par exemple de pluie) pour leur reproduction sexuée aquatique. L'absence de tissus conducteurs pose des problèmes de soutien en raison de la poussée d'Archimède réduite en milieu aérien, d'où leur petite taille qui ne dépasse généralement pas 7 cm[63],[64]. Elles disposent de rhizoïdes assurant l'ancrage, tandis que l'absorption de l'eau et des minéraux est réalisée par une symbiose fongique avec l'équivalent de mycorhizes[65].
Peu de changements morphologiques sont observés depuis les fossiles jusqu'aux formes actuelles, ce qui fait des bryophytes le groupe le plus stable morphologiquement des plantes terrestres[66].
Les anthocérotes forment un lien important pour l'étude de l'évolution des plantes car il représente un groupe frère des végétaux vasculaires[67]. Ils montrent le passage de la phase gamétophytique dominante au profit du sporophyte dominant des plantes vasculaires (réduction de taille mais aussi du temps de vie du gamétophyte), conduisant au succès évolutif des plantes à fleurs. Une hypothèse est que la réduction de cette phase gamétophytique du cycle biologique au cours de la conquête des terres (milieu difficile) pourrait être que la diploïdie du sporophyte autorise le masquage de l'expression de mutations délétères (provenant d'une exposition aux UV) par complémentation génétique[68],[69].
Classification des bryophytes
- Hepaticophyta (hépatiques), l'embranchement aux caractères les plus primitifs
- Marchantiopsida (hépatiques à thalle complexe)
- Jungermanniopsida (hépatiques à thalle simple ou à feuille;
- Anthocerotophyta (anthocérotes) petit embranchement voisin des hépatiques
- Bryophyta (mousses), en 3 classes
- Sphagnopsida (sphaignes)
- Andreaeopsida (andréales)
- Bryopsida (vraies mousses)
Avec près de 25 000 espèces de mousses et sphaignes, 9 000 espèces d'Hépatiques (Marchantiophytes) et 300 espèces d'anthocérotes, les bryophytes constituent le second groupe de végétaux terrestres, après les Dicotylédones[70].
En France, près de 1 300 espèces sont connues, le pays comptant de nombreux taxons très rares[71].
La phylogénétique moléculaire actuelle remet en cause les caractères morphologiques utilisés pour la classification classique, en montrant qu'il y a souvent une homoplasie substantielle dans plusieurs de ces caractères employés pour définir des genres, des familles et des sous-ordres[72].
État, pression, menaces sur les populations de bryophytes
Certaines espèces ont disparu de régions entières, ou doivent être considérées comme rares, menacées ou en danger (une liste rouge de bryophytes a par exemple été publiée pour le Luxembourg[73]).
D'autres sont devenues envahissantes.

 Buxbaumia viridis, espèce protégée en France.
Buxbaumia viridis, espèce protégée en France.
Notes et références
- (en) A. Smith, Bryophyte Ecology, Springer Science & Business Media, , p. 314.
- Jean-Claude Rameau, Dominique Mansion, Gérard Dumé, Flore forestière française : guide écologique illustré, Forêt privée française, , p. 173.
- (en) T. Hallingback & N. Hodgetts , Status Survey and Conservation Action Plan for Bryophytes. Mosses, Liverworts and Hornworts, IUCN, World conservation union, Cambridge, 2000, p. 106 p.
- Jean-Louis De Sloover, Anne-Marie Bogaert-Damin, Les Muscinées du XVIe au XIXe siècle dans les collections de la Bibliothèque universitaire Moretus Plantin, Presses universitaires de Namur, , p. 103.
- Jean Vallade, L'œil de lynx des microscopistes. La sexualité végétale : l'apport des micrographes depuis le XVIIe siècle, Éditions universitaires de Dijon, , p. 111.
- J. Hedwig, Theoria generationis et fructificationis plantarum cryptogamarum Linnæi, Pétersbourg, 1784, p. 52 sqq
- (en) Anupama Krishna, Botany for Degree Students. Bryophyta, S. Chand Publishing, , p. 12.
- (en) A. Jonathan Shaw, Bernard Goffinet, Bryophyte Biology, Cambridge University Press, , p. 135.
- (en) K.S. Renzaglia, J.C Villareal & R.J. Duff. 2008. New insights into morphology, anatomy and systematics of hornworts, in : Goffinet, B., & Shaw, A.J. (eds), Bryophyte Biology, Cambridge University Press, p. 139–171
- L'étymologie de ce nom scientifique fait référence aux corbeilles en forme de lunule, de croissant, cruciata aux pieds du sporophyte en croix. Ces propagules se forment au fond de la corbeille, puis s'élèvent sur un court pédicule et enfin se détachent.
- Daniel Richard, Martine Hausberger, Patrick Chevalet, Sylvie Fournel, Nathalie Giraud, Frédéric Gros, Patrick Laurenti, Fabienne Pradere, Thierry Soubaya, Frédéric Ysnel, Biologie, Dunod, , p. 115.
- (en) R. E. Longton, « Reproductive biology and evolutionary potential in bryophytes », Journal of the Hattori Botanical Laboratory, vol. 41, , p. 205-223.
- (en) Kishan Gopal Ramawat, Jean-Michel Mérillon, K. R. Shivanna, Reproductive Biology of Plants, CRC Press, , p. 64.
- Brown RC, Lemmon BE. 2011 Spores before sporophytes: hypothesizing the origin of sporogenesis at the algal-plant transition. New Phytol. 190, 875–881
- Yves Tourte, Michel Bordonneau, Max Henry et Catherine Tourte, Le monde des végétaux. Organisation, physiologie et génomique, Dunod, , p. 48.
- Amélie Pichonet, « Variabilité et dispersion au sein du genre Dicranum Hedw. (Dicranaceae, Bryophyta) : approches moléculaire et morphologique », Thèse Museum National d'Histoire Naturelle, 2011, p. 6
- Amélie Pichonet, op. cit., p. 7
- (en) A.E. Newton, B.D. Mishler, « The evolutionary significance of asexual reproduction in mosses », Journal of the Hattori Botanical Laboratory, vol. 76, , p. 127-145.
- (en) J.G. Parsons, A. Cairns, C.N. Johnson, S.K.A. Robson, L.A. Shilton, D.A. Westcott, « Bryophyte dispersal by flying foxes: a novel discovery », Oecologia, vol. 152, no 1, , p. 112–114.
- (en) Jörgen Rudolphi, « Ant-mediated dispersal of asexual moss propagules », The Bryologist, vol. 112, no 1, , p. 73-79 (DOI 10.1639/0007-2745-112.1.73).
- (en) R. W. Kimmerer & C.C. Young, « The role of slugs in dispersal of the asexual propagules of dicranum flagellare », The bryologist, vol. 98, no 1, , p. 149-153 (DOI 10.2307/3243652).
- (en) Christopher G. Eckert, « The loss of sex in clonal plants », Evolutionary Ecology, vol. 15, nos 4–6, , p. 501–520 (DOI 10.1023/A:101600551).
- Vincent Chassany, Marie Potage et Maud Ricou, Biologie végétale, Dunod, , p. 81.
- (en) David H. S. Richardson, The biology of mosses, John Wiley & Sons Canada, , p. 25-26.
- Marc-André Selosse, Les Goûts et les couleurs du monde. Une histoire naturelle des tannins, de l'écologie à la santé, Actes Sud Nature, , p. 91
- Cette coupe permet d'observer, de l'extérieur à l'intérieur, une assise superficielle, à valeur d'épiderme ; une zone corticale constituée de cellules parenchymateuse ; un cordon central formé de leptoïdes et d'hydroïdes. Cf Valérie Boutin, Jean-François Fogelgesang, Jean-François Beaux et Françoise Ribola, Atlas de biologie végétale, Dunod, , p. 17.
- Finement dentée, la feuille ou pseudophylle porte à sa surface supérieure des lamelles chlorophylliennes uniassisiales dressées de 5 à 10 cellules de haut. Le cordon conducteur est normalement composé d'hydroïdes et de leptoïdes. Ces lamelles constituées d'une seule assise de cellules ont pour effet d'accroître la surface d'échange et de favoriser la diffusion de CO2, facteur limitant chez ces mousses caractérisées par l'absence de stomates au niveau du gamétophyte. Cf (en) Mariann Marschall, Michael CF Proctor, « Are Bryophytes Shade Plants? Photosynthetic Light Responses and Proportions of Chlorophyll a, Chlorophyll b and Total Carotenoids », Annals of Botany, vol. 94, no 4, , p. 603 (DOI 10.1093/aob/mch178).
- Håvard Kauserud, Cecilie Mathiesen et Mikael Ohlson, « High diversity of fungi associated with living parts of boreal forest bryophytes », Botanique, vol. 86, no 11, , p. 1326-1333 (DOI 10.1139/B08-102, résumé)
- (en) T. J. Carleton & D. J. Read, « Ectomycorrhizas and nutrient transfer in conifer – feather moss ecosystems », Revue canadienne de botanique, vol. 69, no 4, , p. 778-785 (DOI 10.1139/b91-101).
- R.E. Longton (1988), Biology of polar bryophytes and lichens. British Bryological Society Cambridge University Press..
- P. Meyer et M. Schmidt, « Accumulation of dead wood in abandoned beech (Fagus sylvatica L.) forests in northwestern Germany », Forest Ecology and Management, vol. 261, , p. 343–352 (résumé)
- J. Bardat. 2006. Écologie des bryophytes. In Lamy, D., Decocq, G., Boudier, P., Chavoutier, J., Bardat, J., Hugonnot, V., & Rausch de Trautenberg, C. (eds), Bulletin Mycologique et Botanique Dauphiné-Savoie, p. 47–61
- Du grec rugchos bec d'oiseau, et eu, bien ; du latin praelongus très long, allusion au rostre très long de l'opercule au sommet de la capsule portée par le sporophyte.
- Jean-Claude Rameau, Dominique Mansion, Gérard Dumé, Flore forestière française. Plaines et collines, Forêt privée française, , p. 89.
- (en) David H. S. Richardson, The biology of mosses, John Wiley & Sons Canada, , p. 37.
- (en) Martin I Bidartondo, Thomas D Bruns, Michael Weiss, Cecília Sérgio & and David J Read, « Specialized cheating of the ectomycorrhizal symbiosis by an epiparasitic liverwort », Proc Biol Sci., vol. 270, no 1517, , p. 835–842 (DOI 10.1098/rspb.2002.2299).
- B.V. Tooren, « Effects of a bryophyte layer on the emergence of seedlings of chalk grassland species », Acta Oecologica, vol. 11, no 2, , p. 155-163 (lire en ligne)
- (en) Jennifer Britt, Charles Deeming, « First-egg date and air temperature affect nest construction in Blue Tits Cyanistes caeruleus, but not in Great Tits Parus major », Bird Study, vol. 58, no 1, , p. 78-89 (DOI 10.1080/00063657.2010.524916).
- (en) O. O. Oyesiku & O. R. Ogunkolade, « The relationship between the Nigerian garden snail Limicolaria aurora and the moss Hyophila crenulata », Journal of Bryology, vol. 28, no 2, (DOI 10.1179/174328206X105380).
- (en) U. Gerson, « Moss-arthropod association », The Bryologist, vol. 72, , p. 495-500.
- (en) Claudia Ihl & Perry S. Barboza, « Nutritional Value of Moss for Arctic Ruminants: A Test with Muskoxen », The Journal of Wildlife Management, vol. 71, no 3, , p. 752-758.
- (en) Longton, R. E. (1992) The role of bryophytes and lichens in terrestrial ecosystems. In, J. W. Bates & A. M. Farmer (eds). Bryophytes in a Changing Environment, 32-76. Clarendon Press, Oxford
- (en) Alain Vanderpoorten et Bernard Goffinet, Introduction to Bryophytes, Cambridge University Press, , p. 35.
- (en) Bojana Božanić et al, « Factors Affecting Invertebrate Assemblages in Bryophytes of the Litovelské Luhy National Nature Reserve, Czech Republic », Acat Zoologica Bulgarica, vol. 65, no 2, , p. 197-206.
- (en) Gerson U. 1982. Bryophytes and Invertebrates. – In: Smith A. J. E. (Ed.): Bryophyte Ecology. London, New York (Chapman & Hall Ltd), p. 291-332.
- (en) Jan-Peter Frahm, « Recent Developments of Commercial Products from Bryophytes », The Bryologist, vol. 107, no 3, , p. 277-283.
- Sébastien Leblond & Anabelle Boucher, « Initiation à la bryologie - Voyage au cœur de la vie secrète des mousses », 2011, p.24
- C. Ah-Peng, P. Staménoff, J. Bardat, T. Hedderson, L. Marline, N. Wilding, D. Strasberg, « Initiation à l'etude des bryophytes des îles du Sud Ouest de l’océan Indien », 2014, Faculté des sciences Université de la Réunion, p. 6
- (en) Ando, H. & A. Matsuo (1984). Applied bryology, In W. Schultze-Motel (ed.), Advances in Bryology, vol 2 Cramer, p. 133- 224.
- A.M. Kooijman et C. Bakker, « Species replacement in the bryophyte layer in mires: the role of water type, nutrient supply and interspecific interactions », Journal of Ecology, vol. 83, , p. 1–8 (résumé)
- A.M. Kooijman et C. Bakker, « The acidification capacity of wetland bryophytes as influenced by clean and polluted rain », Aquatic Botany, vol. 48, , p. 133–144 (DOI 10.1016/0304-3770(94)90080-9, résumé)
- A.M. Kooijman (1993), Changes in the bryophyte layer of rich fens as controlled by acidification and eutrophication. PhD Thesis, Utrecht University, the Netherlands
- Ö. Fritz, L. Gustafsson et K. Larsson, « Does forest continuity matter in conservation? - A study of epiphytic lichens and bryophytes in beech forests of Southern Sweden », Biological Conservation, vol. 141, , p. 655–668 (résumé)
- (en) Theresa Greenaway, Mosses and Liverworts, Steck-Vaughn, , p. 20.
- J.P. Frahm et D. Klaus, « Bryophytes as indicators of recent climate fluctuations in Central Europe », Lindbergia, vol. 26, no 2, , p. 97–104
- R. Wattenhofer, Contribution à l'étude des macrophytes du Léman, Conservatoire Botanique de Genève, , p. 115.
- (en) F. M. Hueber, « Hepaticites devonicus, a New Fossil Liverwort from the Devonian of New York », Annals of the Missouri Botanical Garden, vol. 48, no 2, , p. 125–131 (DOI 10.2307/2394879).
- (en) Douglas E. Soltis, Pamela Soltis, J.J. Doyle, Molecular Systematics of Plants II : DNA Sequencing, Springer, , p. 530
- « Du Silurien au Dévonien : les sorties des eaux », sur cnrs.fr (consulté le ).
- Propriété de certains êtres vivants d'effectuer un retour aux manifestations de la vie, après être entrés en anhydrobiose sous l'effet de la dessiccation, lorsque les conditions extrêmes (sécheresse, climat…) qu’ils ont dû endurer sont terminées. Les bryophytes elles ont développé la capacité de supporter une période de sécheresse en cessant toute activité métabolique. « Pour résister à la dessiccation, elles appliquent leurs feuilles contre la tige ou retournent leur thalle et prennent alors une apparence brune et rabougrie. Cette phase de « dormance » peut perdurer jusqu'à une trentaine d'années pour certaines espèces ». Cf Laurent Burgisser & Ariane Cailliau, « Les mousses » : Liste Rouge, inventaire et initiation aux bryophytes du canton de Genève, Conservatoire et jardin botaniques de la ville de Genève, 2012 , p. 32
- Cette colonisation de milieux oligotrophes pauvres en azote est permise grâce à l'association symbiotique avec des cyanobactéries du type Nostoc réalisant la photosynthèse et fixant l'azote. Cf. (en) Adams, D. G. & Duggan. P. S. (2008) Cyanobacteria-bryophyte symbioses. Conference Information: Annual Meeting of the Society-for-Experimental-Biology, MAR 31-APR 04, 2007 Glasgow, Scotland. Journal of Experimental Botany 59: 1947-1058
- (en) P. Alpert & M. J. Oliver (2002) Drying without dyin, In: Black M, Prichard HW (eds) Desiccation and survival in plants: drying without dying, CABI, Wallingford, p. 3–43
- (en) Hugues Renault, Annette Alber, Nelly A. Horst, Alexandra Basilio Lopes, Eric A. Fich, Lucie Kriegshauser, Gertrud Wiedemann, Pascaline Ullmann, Laurence Herrgott, Mathieu Erhardt, Emmanuelle Pineau, Jürgen Ehlting, Martine Schmitt, Jocelyn K. C. Rose, Ralf Reski & Danièle Werck-Reichhart, « A phenol-enriched cuticle is ancestral to lignin evolution in land plants », Nature Communications, vol. 8, no 14713, (DOI 10.1038/ncomms14713).
- Peter H Raven, Susan R Singer, Georges B Johnson, Kenneth A Mason, Jonathan B Losos, Biologie, De Boeck Superieur, (lire en ligne), p. 591.
- Jean Garbaye, La symbiose mycorhizienne, Quae, , p. 121.
- (en) R. Reski, « Development, Genetics and Molecular Biology of Mosses », Plant biology, vol. 111, no 1, , p. 10 (DOI 10.1111/j.1438-8677.1998.tb00670.).
- Peter H Raven, op. cit., p. 593
- (en) H. Bernstein, G.S. Byers et R.E. Michod, « Evolution of sexual reproduction: Importance of DNA repair, complementation, and variation », The American Naturalist, vol. 117, no 4, , p. 537–549 (DOI 10.1086/283734).
- (en) R.E. Michod et T.W. Gayley, « Masking of mutations and the evolution of sex », The American Naturalist, vol. 139, no 4, , p. 706–734 (DOI 10.1086/285354).
- (en)Buck, W.R. & Goffinet, B. 2000. 3. Morphology and classification of mosses. In A.J. Shaw & B. Goffinet (eds.), Bryophyte Biology. Cambridge University Press, p. 71
- Vincent Hugonnot, Un haut-lieu pour la conservation des bryophytes : les gorges de la Rhue (Cantal), Journée Forêt & Biodiversité du 17 octobre 2013
- B. Crandall-Stotler, R. E. Stotler & D. G. Long, Morphology and classification of the marchantiophyta, in Goffinet, B., & Shaw, A.J. (eds), Bryophyte Biology, Cambridge University Press, 2008, p. 1–54
- Werner J (1987) Liste rouge des bryophytes du Grand-Duché de Luxembourg. Musée d'histoire naturelle (PDF, 46 p).
Voir aussi
Bibliographie
- Frey, W., Frahm, J.P., Fischer, E., & Lobin, W. (2006), The Liverworts, Mosses and Ferns of Europe. Harley Books, Colchester
- Goffinet, B., Buck, W.R., & Shaw, A.J. (2008), Morphology, anatomy and classification of the Bryophyta. Pages 55–138 of: B., Goffinet, & A.J., Shaw (eds), Bryophyte Biology second edition. Cambridge University Press.
- Prune Pellet - Bryoflore des pierriers gréseux du massif du Suquet (Aigoual, Cévennes), Répartition, inventaire, caractérisation bryocénotiques et écologiques et évaluation de l'intérêt patrimonial, Rapport de stage Parc National des Cévennes, MNHN. Mis en ligne par Tela-Botanica.
- Vincent Hugonnot, Mousses et hépatiques de Païolive (Ardèche et Gard, France), Jarnac, Société Botanique du Centre-Ouest, , 293 p. (ISSN 0759-934X)
- Vincent Hugonnot, Jaoua Celle, Florine Pépin, Mousses et hépatiques de France, Biotope Editions, 2015, 320 p. (ISBN 9782366621952)
- (en) Alain Vanderpoorten et Bernard Goffinet, Introduction to Bryophytes, Cambridge University Press, , 293 p. (lire en ligne)
Articles connexes
Liens externes
- Bryophytes, article en ligne de l'Encyclopædia Universalis
- Les bryophytes, présentation du Centre de ressources pédagogiques en Biologie Végétale de l'Université de Bourgogne
- (fr+en) Référence ITIS : — Hepaticophyta
- (fr+en) Référence ITIS : — Anthocerotophyta
- (fr+en) Référence ITIS : — Bryophyta
- Bryophytes de France
- (en) Bryophyta. Tree of life
- Bryologia Gallica
- Clés de détermination (chapitre 3)
 Portail origine et évolution du vivant
Portail origine et évolution du vivant  Portail de la botanique
Portail de la botanique