Faune abyssale
Au sens strict, la faune abyssale désigne l'ensemble des animaux vivant dans la zone abyssale des océans, c'est-à-dire vivant entre 4 000 et 6 000 m de profondeur. Toutefois, dans l'usage courant, l'expression faune abyssale désigne généralement l'ensemble des espèces vivant à partir de 200 mètres de profondeur et au-delà. À ces profondeurs, les caractéristiques de l'environnement sont très difficiles : la pression de l'eau est très élevée, la température est très basse et quasi constante (sauf à proximité des cheminées hydrothermales où elle est au contraire, très haute), l'oxygène n'est disponible qu'en quantité très limitée, la nourriture est très rare et, surtout, la lumière venant du soleil ne pénètre pratiquement plus : c'est la zone dite aphotique, elle-même divisée en zones mésale (200-1000 m), bathyale (1000-4000 m), abyssale (4000-6000 m) et hadale (plus de 6 000 m). La photosynthèse y étant difficile (voire impossible à partir d'environ 1 000 m), les végétaux ne peuvent s'y développer ; cet environnement est donc seulement occupé par des animaux et certains microorganismes non végétaux (protistes, bactéries, archées, virus, etc.).

La faune abyssale inclut des représentants de presque tous les embranchements d'animaux marins (dont quelques espèces amphibies, puisque l'éléphant de mer peut plonger à plus de 1 580 mètres de profondeur, la tortue luth à 1 200 mètres et le manchot à 350 mètres) mais de nombreuses espèces se sont adaptées aux conditions difficiles de la vie abyssale, soit directement en contact avec le fond marin (benthos) soit dans les eaux situées au-dessus (pélagos). Parmi les caractéristiques originales, les plus connues sont :
- la bioluminescence, la production de lumière « froide » dans des cellules spécialisées de l'épiderme ou dans des bactéries symbiotiques, comme Vibrio fischeri utilisée pour la chasse, la défense ou la recherche de partenaires sexuels dans des espaces gigantesques ;
- les modifications morphologiques et physiologiques : développement d'un illicium, adaptation aux grandes pressions (cages thoraciques plastiques, os et muscles plus légers, corps remplis de fluides et sans air, vessie natatoire réduites ou absentes chez certains poissons, etc.) ;
- l’adaptation à la vie à proximité des monts hydrothermaux, des suintements froids qui émettent des gaz (sulfure d'hydrogène pour les fumeurs noirs, silices et anhydrites pour les fumeurs blancs, méthane et hydrocarbures pour certains suintements froids, etc.) qui font l'objet d'une chimiosynthèse par les bactéries (parfois symbiotiques) ce qui permet à de nombreux animaux de se nourrir et de se protéger de la chaleur des cheminées hydrothermales[réf. nécessaire].
Historique

En 1839, le naturaliste Edward Forbes reçoit une subvention de la British Association for the Advancement of Science pour effectuer des dragages afin d'étudier les espèces marines animales et végétales. Au cours d’une campagne de dragages en mer Égée en 1840-1841, il effectue des prélèvements à des profondeurs croissantes et observe que le nombre d’animaux dans chaque drague diminue avec la profondeur. Sans dépasser les 130 brasses (238 m), il dresse alors une courbe théorique de diminution des peuplements en fonction de la profondeur, courbe qui extrapolée, lui montre que toute vie disparaîtrait au-delà de 550 m[1]. Il synthétise ses résultats en répartissant les espèces en cinq zones spécifiques : littorale, laminaire (jusqu'à 27 m de profondeur), coralline (jusqu'à 90 m), zone de coraux de mer profonde (de 90 à 550 m) et zone abyssale azoïque (absence de vie au-delà de 550 m)[2]. Forbes reprend ainsi l'hypothèse azoïque (« azoic hypothesis ») du géologue Henry de La Beche formulée en 1864 et fixe la zone à 550 m[3] (zone où la lumière est complètement arrêtée, stoppant la photosynthèse, et où les pressions seraient trop fortes pour toute forme de vie). Ardemment défendue par ses disciples et devenue un dogme qui inhibe toutes recherches marines en profondeur pendant plusieurs décennies[4], sa théorie est pourtant biaisée par ses expériences sur la mer Égée particulièrement pauvre et qui ne peut être considérée comme un modèle de l'ensemble des océans[5]. De plus, elle est contredite par des études et des rapports d'autres explorations qui situent cette zone beaucoup plus profondément (études d'Antoine Risso en 1810[6], sondages de John Ross en 1819, expédition Erebus et Terror de James Clark Ross, article de Michael Sars en 1846). La théorie abyssale azoïque résiste car ses partisans considèrent que les mesures de profondeur lors des sondages ne sont pas fiables. Ils ne peuvent cependant remettre en doute la découverte en 1860. Un câble télégraphique sous-marin cassé qui relie la Sardaigne et la Tunisie, posé à 1000 fathoms (1800 mètres) de profondeur, doit être remonté. Les ingénieurs constatant que le câble est encroûté d'organismes vivants, ils envoient des sections aux naturalistes Alphonse Milne-Edwards et George James Allman qui découvrent des coraux et des vers qui se sont développés à 1800 mètres[7]. L'expédition du Challenger lancée dans les années 1870 par Charles Wyville Thomson achève de rendre obsolète la théorie de Forbes. Au cours des 273 campagnes abyssales océanographiques du Prince Albert de Monaco à la fin du XXe siècle, les dragages ramènent des animaux, poissons et crustacés récoltés jusqu'à 6 035 m[8]. L'expédition danoise de la Galathea (en) au début des années 1950 rapporte 115 espèces d’animaux capturés au-delà de 6 000 m et, de la fosse des Mariannes, des bactéries et animaux invertébrés vivant à plus de 10 000 m de profondeur[9]. Ce n'est que dans les années 1960, grâce aux mailles plus fines des tamis, que l'on découvre la grande diversité de la faune profonde. En 1977, la découverte des monts hydrothermaux bouleverse encore plus les conceptions périmées d'uniformité de l'environnement et de la faune abyssale[10].
De nos jours, plusieurs institutions de recherche poursuivent d'importants programmes d'exploration des abysses, comme le Monterey Bay Aquarium Research Institute (en) et l'agence américaine NOAA, qui par son programme Okeanos Explorer diffuse en direct les images des abysses prises par son submersible (ROV) sur Youtube.
Répartition de la faune abyssale
Zone mésale
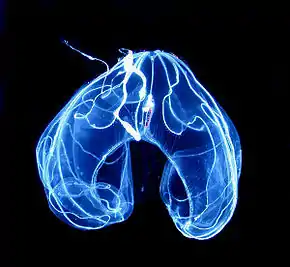
Appelée aussi zone crépusculaire ou zone de pénombre, elle commence à 200 m et termine à 1 000 m. Elle est caractérisée par une faible luminosité, empêchant le développement de la photosynthèse chez les plantes. Algues, alismatales aquatiques et diatomées sont donc absents.
Les sources de nourriture se font rares dans les profondeurs ; outre la neige marine, les seules sources de nourriture disponibles sont les animaux abyssaux eux-mêmes ; expliquant la grande population de prédateurs en ces lieux. Tout en bas de la chaîne alimentaire, se trouvent les plus petits organismes zooplanctoniques, nanoplanctoniques et picoplanctoniques, qui migrent vers la zone euphotique pour se nourrir de phytoplancton, avant de revenir à leur couche initiale.
Sur le pélagos, les prédateurs sont abondants : calmars, salpes, méduses, siphonophores, cténophores, crevettes, poissons cartilagineux (requins, chimères, etc.) ainsi que de nombreux poissons osseux, comme le poisson hachette, les poissons dragons (Malacosteus, Aristostomias et Pachystomias), le régalec, le barracudina ou encore le scopélide.
Sur le benthos, vers tubicoles, actinies, crinoïdes, bivalves, brachiopodes, gorgones, pennatules, bryozoaires, tuniciers, éponges, holothuries, hydroïdes, galathées, raies, poissons plats, et autres sont abondants.
Zone bathyale

La zone bathyale s’étend de 1 000 à 4 000 mètres de profondeur. C’est le plus grand de tous les étages océaniques. À partir de 1 000 mètres, la lumière ne pénètre plus, hormis une faible lumière bleue qui a pour origine la bioluminescence animale.
Sur le pélagos, les animaux bioluminescents sont nombreux. On trouve de nombreux céphalopodes (comme le calmar vampire, la pieuvre dumbo ou le calmar diaphane, par exemple) des nombreuses crevettes, des cténophores, requins, ainsi que de nombreux poissons osseux, dont beaucoup utilisent la bioluminescence pour chasser ou se défendre : linophryne (baudroies abyssales), anoplogaster, grangousiers, Lasiognathus, grenadiers, hoplosthètes, etc.
Sur le benthos, (qui comprend le talus continental) on trouve notamment des lis de mer, des éponges, des ophiures, des raies et des poissons plats. C'est souvent dans cette zone (ainsi que parfois sur la plaine abyssale) que se déposent de grandes carcasses de baleines et de gros poissons ; ces grandes sources de nourriture génèrent, sur les fonds boueux où ces cadavres se déposent, un grand attrait pour de nombreux animaux charognards, comme les myxines, les laimargues et divers arthropodes ainsi qu'un nombre incalculable de vers, bactéries et autres microorganismes se nourrissant des corps morts en décomposition de grands animaux déposés sur les grands fonds.
C'est aussi à cet étage que l'on trouve la grande majorité des communautés hydrothermales.
 Une étoile de mer (Benthopecten sp.) et une ophiure au mont sous-marin de Davidson (-2461 m)
Une étoile de mer (Benthopecten sp.) et une ophiure au mont sous-marin de Davidson (-2461 m) Un poulpe (Bentoctopus sp.) un crinoïde et une actinie au mont sous-marin de Davidson (-2422 m)
Un poulpe (Bentoctopus sp.) un crinoïde et une actinie au mont sous-marin de Davidson (-2422 m) Un poulpe (Benthoctopus sp.) et un bivalve (Acesta mori) au mont sous-marin de Davidson à 1461 mètres de profondeur.
Un poulpe (Benthoctopus sp.) et un bivalve (Acesta mori) au mont sous-marin de Davidson à 1461 mètres de profondeur. Bothrocara brunneum (-3228 m).
Bothrocara brunneum (-3228 m)..jpg.webp) Un jeune Careproctus ovigerum, et une éponge du genre Asbestopluma à 1324 mètres de profondeur (mont sous-marin de Davidson).
Un jeune Careproctus ovigerum, et une éponge du genre Asbestopluma à 1324 mètres de profondeur (mont sous-marin de Davidson). Bathychaunax coloratus à 2461 mètres de profondeur (mont sous-marin de Davidson).
Bathychaunax coloratus à 2461 mètres de profondeur (mont sous-marin de Davidson). Pachycara sp. (-2602 m).
Pachycara sp. (-2602 m). Un poulpe (Graneledone boreopacifica) et un type de corail noir (Trissopathes sp.) au mont sous-marin de Davidson (-1973 m)
Un poulpe (Graneledone boreopacifica) et un type de corail noir (Trissopathes sp.) au mont sous-marin de Davidson (-1973 m) Une actinie photographiée à 1874 mètres de profondeur (mont sous-marin de Davidson).
Une actinie photographiée à 1874 mètres de profondeur (mont sous-marin de Davidson). Un calamar (Gonatus onyx) Au mont sous-marin de Davidson (-1328 m).
Un calamar (Gonatus onyx) Au mont sous-marin de Davidson (-1328 m). Un poisson (Psychrolutes phrictus), et deux éponges (Staurocalyptus sp. et Ferrea sp.) Au mont sous-marin de Davidson (-1317 m)
Un poisson (Psychrolutes phrictus), et deux éponges (Staurocalyptus sp. et Ferrea sp.) Au mont sous-marin de Davidson (-1317 m).jpg.webp) Un halosaur (Aldrovandia sp.)
Un halosaur (Aldrovandia sp.) Une éponge cheminée.
Une éponge cheminée.
Zone abyssale

La zone abyssale, qui s’étend de 4 000 à 6 000 m de profondeur, n'a plus de luminosité du tout. Elle comprend la plaine abyssale, vaste étendue de sédiments boueux et vaseux. Ces sédiments et limons ont pour origine les minéraux (graviers, roches, sables, vases, etc.) et la décomposition de corps de milliards de créatures marines (plancton ou necton) accumulés pendant des millions d'années, formant dans certains endroits des couches de sédiments atteignant plusieurs kilomètres de haut. Sur le plancher océanique, les sédiments peuvent aller jusqu'à 1 000 mètres et jusqu'à 15 km sur le talus continental. Le maximum enregistré est de 20 km[11].
Sur le pelagos vivent de nombreuses espèces, comme de nombreuses crevettes et certains poissons, comme le rat tacheté, mais sont plus rares du fait du manque de nourriture. Par contre, sur le benthos, la vie est très abondante : les sédiments grouillent de foraminifères, de bactéries et de vers. Pour la plupart nécrophages, se nourrissant des matières organiques déposées sur la plaine abyssale. De nombreuses espèces vivent fixées sur la boue, mais doivent rester de manière permanente au-dessus. On compte notamment les lis de mer, les pennatules et les éponges (notamment les euplectelles). Des holothuries, des oursins et des euryalina se déplacent sur les sédiments, avec des pattes assez longues comme pour ne pas s'y enfoncer. Des poissons, comme le poisson trépied, se déplacent sur le benthos sédimenteux à l'aide de ses nageoires en forme d'échasses.
Il a été démontré que la biodiversité présente dans les sédiments de la plaine abyssale est aussi diverse que celle de la forêt tropicale[12].
Zone hadale
La zone hadale, qui va de 6 000 m à au-delà (la profondeur maximale enregistrée jusqu'à nos jours étant 10 916 mètres, dans la fosse des Mariannes) est la zone la plus méconnue de tous les étages océaniques. La biodiversité est très uniforme dans tous les océans, à cause du peu d'obstacles dans cette zone[13], excepté dans les fosses océaniques, où les obstacles et l'endémisme sont très élevés. Les espèces de cet étage sont peu connues et beaucoup d'endroits restent à explorer.
Sur le pelagos, peu d'espèces le fréquentent. On trouve par exemple des brotulides. Sur le benthos, on trouve par exemple des anémones de mer, des holothuries, des crevettes, des poissons plats.
La zone hadale étant relativement méconnue, il est certain que nombre d'espèces et d'écosystèmes restent à découvrir.
Alimentation de la faune abyssale
La faune abyssale adopte des comportements trophiques ambivalents : suspensivores, détritivores, carnivores, nécrophages. Cet opportunisme alimentaire serait une forme d'adaptation aux maigres apports nutritifs caractérisant le domaine abyssal[14].
En 1968, H. L. Sanders propose la théorie de la stabilité temporelle de l'équilibrium selon laquelle la biodiversité serait expliquée par la stabilité de conditions environnementales physiques et la faiblesse des ressources nutritives ayant permis la mise en place de stratégies adaptatives évolutives qui minimiseraient la compétition, notamment par ces comportements alimentaires variés[15]. Cette théorie est remise en cause par la découverte en 1977 des oasis hydrothermaux à faible biodiversité mais forte productivité primaire[16].
Bioluminescence dans la faune abyssale
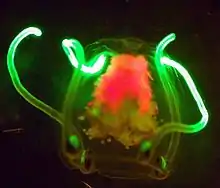
La bioluminescence, production de luminescence froide par des êtres vivants, est très répandue à partir de 200 mètres de profondeur, où la lumière est insuffisante. Cette lumière est créée de trois façons différentes :
- par symbiose avec une bactérie elle-même bioluminescente (Vibrio fischeri) ;
- par réaction entre une molécule, la luciferine, et une enzyme, la luciférase, dans une glande dans la peau ou directement sous la peau ;
- par des cellules spécialisées : les photocytes.
La lumière produite peut être amplifiée ou filtrée pour donner des couleurs caractéristiques grâce à des organes spéciaux : les photophores. Ils peuvent posséder une lentille, un conduit de lumière ou un filtre coloré. Les lumières émises sont souvent bleues, mais peuvent être jaunes, vertes ou rouges.
On connait plusieurs fonctions différentes à la bioluminescence :
- communication entre espèces ou au sein d'une espèce, par émissions soudaines ou clignotantes de lumières (Microorganismes et calamars) ;
- rôle de communication sexuelle, à fin reproductive (utilisée chez certaines baudroies abyssales dont le mâle, minuscule, accroche sa bouche dentée sur le corps de la femelle, fusionnant totalement avec elle après un certain temps, pour échanger du sperme contre des nutriments) ;
- repérage ou attraction des proies (leurres luminescents chez les baudroies abyssales et certains autres poissons et invertébrés, comme le poulpe Stauroteuthis syrtensis) qui attirent les proies avant d'être capturées et mangées, ou, dans le cas d'animaux comme la drague, production de lumières rouges par des photophores sous les yeux pour repérer les proies, la lumière rouge étant invisible pour la plupart des créatures abyssales) ;
- défense par éjection de sécrétions luminescentes sur le prédateur, l'aveuglant pendant quelques secondes (utilisée chez certains invertébrés, comme des calmars, des crevettes, des vers ou des méduses abyssales). D'autres animaux trompent leurs prédateurs : les portes-lanternes font clignoter les organes bioluminescents sur leurs joues. On a récemment découvert un ver Swima bombiviridis, aveugle, mesurant de 18 à 93 mm, et vivant de −1 800 à −3 800 mètres de fond, qui largue des bulles bioluminescentes comme leurre avant de fuir ses prédateurs[18]. Ces leurres sont de minuscules ballonnets emplis d'hémolymphe (diamètre de 0,7 à 1,1 mm) qui croissent près de sa tête. Quand elles sont libérées, ces petites bulles émettent durant quelques secondes une lumière intense qui semble pouvoir dérouter la plupart des prédateurs de cette espèce, laissant au ver quelques secondes de répit pour s'enfuir ;
- camouflage, comme pour le poisson hachette, qui possède en plus de son corps très fin une couleur argentée et des photophores dirigés vers le bas, dont il peut régler l'intensité ; lorsqu'il se trouve dans la lumière plongeante de la zone crépusculaire il devient quasiment invisible, empêchant les prédateurs de le voir quand il est au-dessus d'eux.
 Ce poisson des grands fonds, Photostomias guernei, possède des photophores lui servant à voir dans l'obscurité.
Ce poisson des grands fonds, Photostomias guernei, possède des photophores lui servant à voir dans l'obscurité. Ce hache d'argent (Argyropelecus aculeatus) possède des photophores spéciaux qui lui servent à se confondre avec la lumière tombante.
Ce hache d'argent (Argyropelecus aculeatus) possède des photophores spéciaux qui lui servent à se confondre avec la lumière tombante.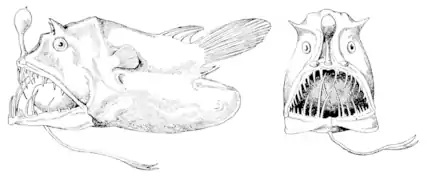 Un exemple de Lophiiformes abyssal utilisant un leurre bioluminescent pour chasser ses proies : Linophryne lucifer.
Un exemple de Lophiiformes abyssal utilisant un leurre bioluminescent pour chasser ses proies : Linophryne lucifer.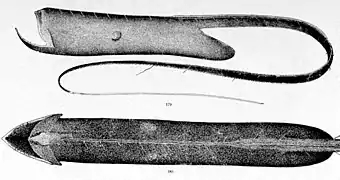 Cette espèce de grangousier, Saccopharynx flagellum, possède, comme toutes les espèces de son genre, un organe bioluminescent au bout de la queue; il sert peut-être de leurre pour attirer les proies.
Cette espèce de grangousier, Saccopharynx flagellum, possède, comme toutes les espèces de son genre, un organe bioluminescent au bout de la queue; il sert peut-être de leurre pour attirer les proies. Cette nouvelle espèce de poisson, découverte dans les eaux profondes antarctiques, possède un organe bioluminescent au bout d'un pédoncule fixé sous sa mâchoire inférieure.
Cette nouvelle espèce de poisson, découverte dans les eaux profondes antarctiques, possède un organe bioluminescent au bout d'un pédoncule fixé sous sa mâchoire inférieure.
Les écosystèmes d'eaux profondes

La zone aphotique compte plusieurs écosystèmes spécifiques riches en biodiversité et ne dépendant pas de la lumière pour vivre. C'est le cas notamment des communautés hydrothermales (fumeurs noirs et blancs) des communautés des suintements froids (qui lâchent du méthane, des hydrocarbures ou autres) mais aussi des récifs de coraux d'eaux froides (Composés d'espèces de cnidaires madréporiques comme lophelia pertusa); Il est certain que bon nombre d'écosystèmes d'eaux profondes restent à découvrir.
Voici une liste non exhaustive des écosystèmes de la zone aphotique connus à ce jour :
- Les communautés hydrothermales qui sont basées sur les monts hydrothermaux. Il en existe deux types connus : Les fumeurs noirs (cheminées lâchant des sulfures d'hydrogène à plus de 400 °C) et les fumeurs blancs (plus petits et aux émanations plus froides et plus lentes que les fumeurs noirs, et qui lâchent de l'anhydrite et de la silice). Sur ces fumeurs, des bactéries spécialisées peuvent fixer ces gaz pour en faire des nutriments : c'est la chimiosynthèse. Bon nombre d'animaux vivent à proximité de ces fumeurs, en symbiose avec ces bactéries. Les écosystèmes peuvent varier d'un endroit à l'autre. Ainsi, on trouvera, dans le Pacifique, à proximité des fumeurs noirs, le ver riftia, le ver de Pompéi et la Galathée yéti, alors que dans l'Atlantique, au niveau de la dorsale océanique, on trouvera la crevette Rimicaris exoculata.
- Les suintements froids (ou mofettes) beaucoup plus répandus puisque pouvant se trouver dans des eaux peu profondes ou dans des sources chaudes sur Terre. Ce sont des petits trous d'où sortent continuellement des gaz sous forme de bulles qui peuvent être de compositions variées : soufre, dioxyde de carbone, hydrocarbures, méthane, etc. Dans la zone aphotique il arrive que des suintements froids lâchent des hydrocarbures ou du méthane. Des bactéries spécialisées peuvent alors fixer ces gaz pour en faire des nutriments, comme chez les communautés hydrothermales. C'est donc à proximité de ces suintements que de nombreux animaux marins se développent, par exemple vestimentifères, bivalves, coraux mous ou crustacés. C’est dans le golfe du Mexique, à plus de 3 000 mètres de profondeur, que l'on a trouvé, autour d'un lac sous-marin, une grande communauté d'animaux s'étant développés grâce aux suintements de méthane présents autour du lac. Il arrive aussi que des communautés d'animaux vivant en symbiose avec des bactéries fixant le méthane se développent sur des fonds sans suintements de méthane. Dans ce cas, des terres riches en méthane sont apportées par les glissements de terrain et les courants venus plus d'en bas.
- Les récifs de coraux d'eau froide, qui sont composés de cnidaires madréporiques abritant de nombreuses espèces, comme des galathées. Ces coraux ne dépendent pas, à la différence des coraux d'eaux chaudes, de la zooxanthelle; ils peuvent donc aisément se développer en eaux profondes. On trouve notamment des récifs de plusieurs kilomètres de Lophelia pertusa, un peu partout dans le monde, mais beaucoup d'autres espèces d'eaux froides contribuent à l'élaboration de récifs profonds. Ces récifs ont été impactés par des pratiques de pêche quand la capture de l'hoplosthète orange (« empereur ») avait cours, et avant qu'ils ne fassent l'objet de mesures de protection spécifiques, particulièrement en Atlantique nord est (zones Natura 2000, zones fermées au titre de la Commission des pêches de l'Atlantique nord-est).
- Les récifs d'éponges : ces récifs sont constitués essentiellement d'éponges Heterochone calix, mais aussi d'Aphrocallistes vastus et de Farrea occa, qui abritent beaucoup d'espèces et peuvent couvrir plusieurs kilomètres entre 100 et 250 mètres de profondeur[19].
 Araignée de mer, bivalves et vestimentifères à proximité d'une source de méthane dans le golfe du Mexique, à plus de 3 000 mètres de profondeur.
Araignée de mer, bivalves et vestimentifères à proximité d'une source de méthane dans le golfe du Mexique, à plus de 3 000 mètres de profondeur. Un récif de coraux d'eaux froides du genre madrepora.
Un récif de coraux d'eaux froides du genre madrepora. Récif de Lophelia pertusa
Récif de Lophelia pertusa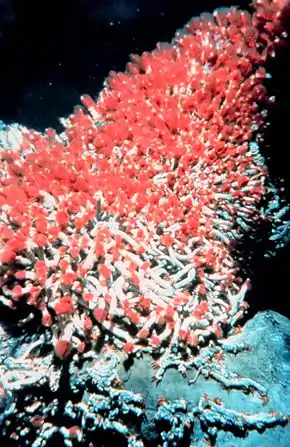


Bivalve géant des communautés hydrothermales le (Bathymodiolus sp.)
(moule abyssale)
Fossiles vivants dans la faune abyssale

Les eaux de la zone aphotique sont connues pour posséder de nombreuses espèces animales aux formes rappelant des espèces fossiles (d'où l'appellation un peu trompeuse de fossiles vivants). Elles ont subi peu de changements morphologiques car elles sont adaptées à la vie dans les abysses, milieux relativement stables à l'échelle évolutive.
C'est le cas des Cœlacanthes, poissons appartenant à une famille déjà présente au Devonien, au Trias et au Jurassique, que l'on pensait disparue jusqu'à ce que l'on pêche, en 1938, un individu au large de l'Afrique du Sud. Aujourd'hui, deux espèces du même genre ont été découvertes : Latimeria chalumnae, qui vit aux Comores et au sud de l'Afrique, et Latimeria menadoensis, trouvé en Indonésie, en 1999. Néanmoins, il est tout à fait probable que d'autres espèces du genre Latimeria n'aient pas encore été découvertes à ce jour.
D'autres espèces de poissons abyssaux sont connues pour leur forme ancienne notable. Le requin-lézard, une espèce de requin des grands fonds trouvé un peu partout dans le monde et notamment au Japon, en est un exemple. Il possède de nombreuses caractéristiques des requins anciens : bouche située à l'avant de la tête, narines situées au-dessus de la tête, corps évoquant une anguille, six paires d'ouïes, etc.
Chez les invertébrés, on trouve les nautiles, cousins des ammonites abondants à 400 mètres de profondeur dans les eaux tropicales de l'Indo Pacifique, mais aussi les lis de mer, les foraminifère ou les brachiopodes, abondants dans leur temps et aujourd'hui plus restreints.
Un exemple notable est celui de Vampyroteuthis infernalis, espèce dont les semblables vivaient en grand nombre dans des eaux moins profondes, il y a plus de deux cent millions d'années avant notre ère. Leur morphologie, rappelant autant le calmar que la pieuvre, fait penser que certains membres de l'ordre des vampyromorphes pourraient être les ancêtres communs de ces deux derniers ordres[20].
 Latimeria menadoensis, l'autre espèce de cœlacanthe vivant connu.
Latimeria menadoensis, l'autre espèce de cœlacanthe vivant connu. Un requin (Chlamydoselachus anguineus) au Davidson seamount (−1 736 m).
Un requin (Chlamydoselachus anguineus) au Davidson seamount (−1 736 m). Microfossiles de radiolaires, foraminifères et éponges trouvés dans des sédiments du talus continental Antarctique.
Microfossiles de radiolaires, foraminifères et éponges trouvés dans des sédiments du talus continental Antarctique. Un Nautile (Nautilus sp.)
Un Nautile (Nautilus sp.)
Galerie

 Une holothurie pélagique abyssale du genre Enypniastes.
Une holothurie pélagique abyssale du genre Enypniastes. Une méduse abyssale (peut-être Peryphilla peryphilla).
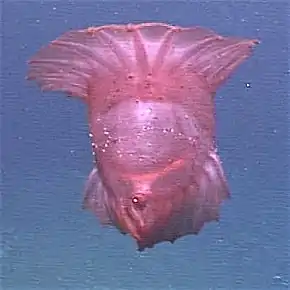
Une méduse abyssale (peut-être Peryphilla peryphilla). Une méduse d'eaux profondes (Tiburonia granrojo) cette espèce de Scyphoméduse a été découverte en 1993 en Californie par 1 500 mètres de fond. Elle se caractérise par une couleur rougeâtre et des bras buccaux d'un nombre variant de 4 à 7[21].
Une méduse d'eaux profondes (Tiburonia granrojo) cette espèce de Scyphoméduse a été découverte en 1993 en Californie par 1 500 mètres de fond. Elle se caractérise par une couleur rougeâtre et des bras buccaux d'un nombre variant de 4 à 7[21].







 Sebastes constellatus.
Sebastes constellatus.

 Gorgonocephalus' sp.
Gorgonocephalus' sp.

Voir aussi
Articles connexes
Liens externes
Notes et références
- (en) Forbes Edward, 1844a. On the light thrown on geology by submarine researches; being the substance of a communication made to the Royal Institution of Great Britain, 23 février 1844. Edinburgh New Philosophical Journal, vol. 36, p. 319–327.
(en) Forbes Edward, 1844b. Report on the Mollusca and Radiata of the Aegean Sea, and on their distribution, considered as bearing on geology. Report of the British Association for the Advancement of Science for 1843, p. 129–193 - Ritchie R. Ward, Into the ocean world : the biology of the sea, Knopf, , p. 66
- Forbes la fixe précisément à 300 fathoms (549 mètres).
- (en) Anderson Thomas R. & Tony Rice, « Deserts on the sea floor: Edward Forbes and his azoic hypothesis for a lifeless deep ocean », Endeavour, vol. 30, no 4, , p. 131-137
- Daniel Desbruyères, Les trésors des abysses, Quae, , p. 90
- Risso observe les poissons et crustacés capturés par les palangriers locaux et décrit des espèces provenant de profondeurs entre 600 et 1 000 m. Cf.Antoine Risso, Ichthyologie de Nice ou Histoire naturelle des poissons du département des Alpes-Maritimes, F. Schoell, , 388 p.
- (en) Susan Schlee, A History of Oceanography, Hale, , p. 89
- Keith Rodney Benson, Philip F. Rehbock, Oceanographic History : The Pacific and Beyond, University of Washington Press, , p. 241
- Jean Jarry, L'aventure des bathyscaphes : marins, ingénieurs et savants au plus profond des mers, Le gerfaut, , p. 31.
- Stéphane Blain, Jean-Luc Charlou et Chantal Compère, La chimie et la mer, EDP Sciences, , p. 124.
- Océan Geo, Encyclopédie universelle/Gallimard, , 181 sur 511 p.
- Océan Geo, Encyclopédie universelle/Gallimard, , 219 sur 511 p.
- Océan Geo, Encyclopédie universelle/Gallimard, , 171 sur 511 p.
- (en) M. N. Sokolova, Data on the feeding of deep-water bottom invertebrates, in Feeding of Marine lnvertebrates in Natural Habitat, Shirshov Institut of Oceanography, pp. 99-101, Academy of Sciences of the USSR, 1986.
- (en) H. L. Sanders, « Marine benthic diversity: A comparative study », The American Naturalist, no 102, , p. 243-282.
- (en) Erik Kristensen, Interactions Between Macro- and Microorganisms in Marine Sediments, American Geophysical Union, , p. 252.
- Claire Nouvian, Abysses, Fayard, , 75 sur 256 p.
- Découverte relatée par la revue Science, n° du 21 août 2009.
- >(fr) « Les récifs spongieux », sur cgc.rncan.gc.ca
- Claire Nouvian, Abysses, Fayard, , 134/135 sur 256 p..
- Claire Nouvian, Abysses, Fayard, , 202 sur 256 p.
 Portail des abysses
Portail des abysses  Portail de la biologie marine
Portail de la biologie marine  Portail de la zoologie
Portail de la zoologie