Choanoflagellata
Les Choanoflagellés, Choanoflagellata ou Choanomonada, sont un petit groupe d’eucaryotes opisthocontes unicellulaires proches des métazoaires. Ces protistes flagellés ont un mode de vie colonial et non-fixé (vivant en pleine eau) ou pour certaines espèces sont associés au périphyton.

| Domaine | Eukaryota |
|---|---|
| Sous-domaine | Unikonta |
| Super-règne | Opisthokonta |
Familles de rang inférieur
- Monosigidae
- Salpingoecidae
- Acanthoecidae
- Choanomonada[1]
On en connait plus de 125 espèces, largement réparties et souvent abondantes. L'inventaire n'en est peut-être pas exhaustif.
Parmi les microorganismes, ils sont considérés comme étant les plus proches parents vivants des premiers animaux et à ce titre sont utilisés par les biologistes évolutionnistes et la biologie évolutive du développement comme modèle pour les reconstructions du dernier ancêtre des animaux unicellulaires. Certains processus biochimiques observés dans notre cerveau pourraient directement dériver de ceux qui sont observés chez les Choanoflagellés[2].
Bien que discrets, ils sont très présents dans les océans où ils jouent un rôle important dans les cycles biogéochimiques et en particulier pour le cycle du carbone et le cycle de la silice.
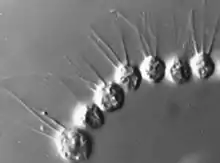
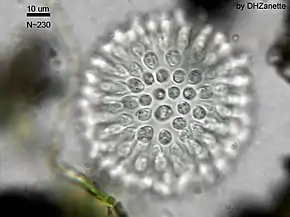
Les traits de vie des Choanoflagellés sont encore mal compris. Beaucoup d'espèces sont supposées solitaires, mais des comportements coloniaux semblent avoir surgi indépendamment et à plusieurs reprises au sein du groupe (mais les espèces coloniales conservent un stade initial solitaire)[3].
Description
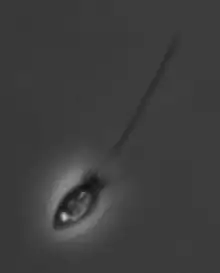
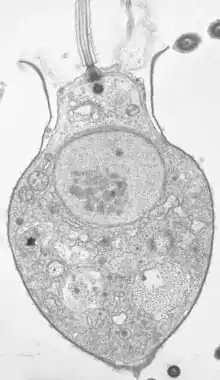
La cellule mesure de 3 à 10 µm et se caractérise par :
- la présence d'une collerette de microvillosités (microvilli, notamment constituée d'actine) au centre de laquelle bat un unique flagelle apical (un appareil de Golgi se trouve à sa base) de structure similaire à celle des choanocytes des éponges. Ce flagelle sert à la fois au déplacement de la cellules (à la manière de la queue d'un spermatozoïde) et à sa nutrition ;
- un type constant d'organisation des organites dans le cytoplasme[4].
- le corps cellulaire d'un grand nombre de Choanoflagellés est entouré d'une matrice (ou périplasme extracellulaire, dont la forme ou la structure est souvent spécifique à l'espèce, ce qui l'a fait utiliser pour l'identification taxonomique ).
Beaucoup de Choanoflagellés construisent un périplasme de structures complexes, constituées de plusieurs bandes de silice cimentées ensemble[5] ; Leur signification biologique est inconnue, mais chez les espèces fixées, on estime que le périplasme joue un rôle dans la fixation à un subtrat ; Chez les microorganismes planctoniques, une hypothèse est que le périplasme pourrait augmenter la traînée de la cellule en mouvement, contrecarrant ainsi la force générée par le flagelle et via des microturbulences contribuant à mieux apporter la nourriture aux vacuoles alimentaires[6] ; - la sécrétion d'une « loge » ovoïde ou sphérique dite « thèque ».
Les Choanoflagellés nagent librement dans la colonne d'eau ou sont sessiles (ils adhèrent alors directement au substrat ou par l'intermédiaire du périplasme ou d'un mince pédoncule[7].
Interactions intraspécifique ou interspécifiques
Selon les connaissances actuellement disponibles, les Choanoflagellés semblent mener une vie toujours hétérotrophe sans directement dépendre d'une autre espèces, mais un certain nombre de parents des Choanoflagellés tels que certains Ichthyosporea ou Mesomycetozoa ont au cours de l'évolution adopté un mode de vie parasitaire ou sont des pathogènes d'autres espèces[8].
Contrairement aux plantes et aux champignons ce groupe se montre capable de former des colonies motile et pouvant se déformer via l'activation collective d'une contractilité (chez les animaux pluricelullaire, la contractilité collective de groupes de cellules est à la base de nombreux processus (ex gastrulation, ingestion, excrétion, réflexion de fuite, motilité musculaire, etc.
Brunet et al. décrivent en 2019 un choanoflagellé récemment découvert (Choanoeca flexa sp. Nov.), qui forme des colonies en forme de coupe (formée d'une monocouche de cellules polarisées) ; ces colonies sont capables de contractilité et de comporement alimentaire collective, mais aussi de changement rapide de morphologie en réponse à une soudaine privation de lumière (la colonie inverse alors son sens de courbure, grâce à une protéine photosensible). Les auteurs notent que les mécanismes cellulaires qui président à ce processus sont les mêmes chez C. flexa et chez les animaux, ce qui éoque un dernier ancêtre commun déjà capable de contracter des cellules polarisées[9].
Reproduction, cycle de vie
les Choanoflagellés ont une croissance végétative, avec pour de nombreuses espèces une reproduction par fission longitudinale[4].
Le cycle de la vie reproductive des Choanoflagellés reste cependant encore incompris. On ignore encore s'il existe aussi une phase sexuelle dans le cycle de vie de ce groupe. Son niveau de ploïdie est également inconnu[4],[10]
La reproduction sexuée n'a pas encore été observée, mais deux rétrotransposons et des gènes clés impliqués dans la méiose ont été découverts dans leur génome[11] suggérant une possible sexualité.
Certains choanoflagellés peuvent s'enkyster (avec perte du flagelle et du collier)[12].
Habitat, répartition
Les Choanoflagellés sont planctoniques ou fixés sur des algues (périphyton).
On les rencontre principalement en milieu marin, de répartition mondiale puisque dans tous les océans. Certaines espèces sont cependant dulçaquicoles (d'eau douce).
Alimentation
Leur nourriture semblaient essentiellement constituée de bactéries, mais des chercheurs ont récemment montré que - au moins dans certaines zones du monde (par exemple dans le Golfe de Gabès sur la côte est de la Tunisie) - ils pouvaient aussi « brouter » de grandes quantités de nanophytoplancton[13].
Le système digestif est rudimentaire, constitué de simples vacuoles alimentaires positionnées dans la partie basale du cytoplasme[5],[4].
Les particules alimentaires (bactéries, microalgues) sont apportées aux vacuoles digestives grâce à un mouvement du flagelle entraînant le déplacement de l'eau qui crée une dépression sur les bords de la collerette, qui conduit à une aspiration de l'eau apportant des bactéries au niveau de la collerette, alimentant ainsi la cellule qui peut les ingérer.
Biosynthèse de la silice
Les Choanoflagellés Acanthoecidae produisent une structure extracellulaire dite lorica (du nom d'une armure romaine faite de bandes). Cette structure est composée de bandes individuelles formées d'un biocomposite (une sorte de polymère composé de silice et de protéines). le matériau de chacune de ces bandes est synthétisé à l'intérieur de la cellule puis excrêtée à l'extérieur, sur la surface de la cellule.
Chez les Choanoflagellés « nudiformes », l'assemblage de ces bandes se fait grâce à un certain nombre de petits tentacules. Elles sont produites par la cellule jusqu'à ce que la lorica soit entièrement réalisée.
Chez les Choanoflagellés dits tectiformes, des bandes costales sont produites en excès et s'accumulent dans une zone situé sous le col. Quand la cellule-mère se divise pour produire une cellule fille, la nouvelle cellule capte une partie de ces bandes dans le cadre de la cytokinèse et assemble sa propre lorica (en n'utilisant que les bandes siliceuses produites précédemment)[14]
La biosilicification n'est possible que si de la silice (en l’occurrence sous forme d'acide silicique) a été préalablement bioconcentrée dans la cellule, ce qui est possible grâce à des protéines transporteuses dédiées dites « SIT » (pour « Silicon Transporter »).
L'étude de ces protéines SIT a montré qu'elles étaient similaires à celles trouvées chez les diatomées et d'autres straménopiles biosynthétisant également des squelettes siliceux, cependant la famille des gènes codant la protéine SIT ne montre que peu ou pas d'homologie avec les autres gènes des Choanoflagellés, y compris de Choanoflagellés non-siliceux ou des straménopiles. Ceci suggère que la famille des gènes de SIT a évolué ailleurs et qu'il y a eu un événement de transfert de gène entre Acanthoecidae et straménopiles. C'est un cas remarquable de transfert horizontal de gènes entre deux groupes eucaryotes lointainement apparentés[15]
Espèces de Choanoflagellés
- Diaphanoeca pedicellata
- Parvicorbicula socialis
- Pleurasiga minima
Articles connexes
Bibliographie
- Boucaud-Camou, E. (1967). Les choanoflagellés des côtes de la Manche: I. Systématique. Bull. Soc. linn. Normandie, 10(7), 191-209.
- Lecointre L & Le Guyader H (année ?) Classification phylogénétique du vivant par aux éditions Belin
- Hollande, A. (1952). Ordre des Choanoflagellés ou Craspédomonadines. Grasse, PP: Traité de Zoologie, 1.
Notes et références
- NCBI, consulté le 8 décembre 2019
- Your brain chemistry existed before animals did newscientist
- Leadbeater, B.S.C. (1983). "Life-History and Ultrastructure of a New Marine Species of Proterospongia (Choanoflagellida)". J. Mar. Biol. Ass. U.K. 63 (63): 135–160. doi:10.1017/S0025315400049857
- Karpov S. & Leadbeater, B.S.C.(1998) Cytoskeleton structure and composition in choanoflagellates, Journal of Eukaryotic Microbiology ; volume 45, no 3 ; p. 361–367 doi:10.1111/j.1550-7408.1998.tb04550.x
- Leadbeater, B.S.C.; Thomsen, H. (2000). "Order Choanoflagellida". An Illustrated Guide to the Protozoa, Second Edition. Lawrence : Society of Protozoologists 451: 14–38
- Leadbeater, B.S.C.; Kelly, M. (2001). "Evolution of animals choanoflagellates and sponges". Water and Atmosphere Online 9 (2): 9–11.
- Leadbeater B.S.C (1983) "Life-History and Ultrastructure of a New Marine Species of Proterospongia (Choanoflagellida)". J. Mar. Biol. Ass. U.K. 63 (63): 135–160. doi:10.1017/S0025315400049857
- Mendoza L.; Taylor, J. and Ajello, L. (2002). "The class Mesomycetozoea : a heterogeneous group of microorganisms at the animal-fungal boundary". Ann. Rev. Microbiol. 56: 315–44. doi:10.1146/annurev.micro.56.012302.160950.
- Thibaut Brunet, Ben T. Larson, Tess A. Linden, Mark J. A. Vermeij, Kent McDonald & Nicole King (2019) Light-regulated collective contractility in a multicellular choanoflagellate|Science 18 Oct 2019: Vol. 366, Issue 6463, p. 326-334 |DOI: 10.1126/science.aay2346
- Claus Nielsen. Animal Evolution: Interrelationships of the Living Phyla. 3rd ed. Claus Nielsen. Oxford, Royaume-Uni: Oxford University Press, 2012, p. 14.
- Carr M.; Leadbeater B., and Baldauf, S. (2002). "Conserved Meiotic Genes Point to Sex in the Choanoflagellates". J. Eukaryot. Microbiol. 57 (1): 56–62. doi:10.1111/j.1550-7408.2009.00450.x .
- Leadbeater, B.S.C.; Karpov, S. (2000). "Cyst Formation in a Freshwater Strain of the Choanoflagellate Desmarella moniliformis Kent". J. Eukaryot. Microbiol. 47 (5): 433–439. Doi:10.1111/j.1550-7408.2000.tb00071.x .
- Hamdi, I., Denis, M., Bellaaj-Zouari, A., Khemakhem, H., Hassen, M. B., Hamza, A.... & Maalej, S. (2014), The characterisation and summer distribution of ultraphytoplankton in the Gulf of Gabès (Eastern Mediterranean Sea, Tunisia) by using flow cytometry revue Continental Shelf Research (résumé) ; doi:10.1016/j.csr.2014.10.002
- Leadbeater, Barry SC; QiBin Yu; Joyce Kent; Dov J Stekel (January 7, 2009). "Three-dimensional images of choanoflagellate loricae". Proceedings of the Royal Society B 276 (1654): 3–11. Doi:10.1098/rspb.2008.0844. PMC 2581655. .
- Marron, Alan O; Mark J. Alston; Darren Heavens; Michael Akam; Mario Caccamo; Peter W. H. Holland; Giselle Walker (2013). "A family of diatom-like silicon transporters in the siliceous loricate choanoflagellates". Proceedings of the Royal Society B 280 (1756): 1471–2954. 7 avril 2013 ; doi:10.1098/rspb.2012.2543
Liens externes
- (en) Référence NCBI : Choanoflagellata Kent, 1880 (syn. de Choanomonada) (taxons inclus) (consulté le )
- ChoanoWiki Ressources collaborativement améliorées et partagées par une communauté de chercheurs étudiant les Choanoflagelés.
- Portail des Choanoflagellés dans l'arbre de la vie (Tolweb
- Monosiga brevicollis (genome browser)
- Choanobase, the Choanoflagellate genetic library, developed and maintained by the Nicole King laboratory at the University of California, Berkeley
 Portail de la zoologie
Portail de la zoologie  Portail de la microbiologie
Portail de la microbiologie