Biologie évolutive du développement
La biologie évolutive du développement souvent nommée évo-dévo (en anglais evo-devo pour Evolutionary Developmental Biology) est un champ disciplinaire en biologie de l'évolution qui a pour objectif de comprendre l'origine de la complexité morphologique des organismes (plantes ou animaux) à travers l'étude comparée des gènes qui régulent leur développement. À l'intersection de la génomique, de la phylogénie moléculaire, de l'embryologie et de la génétique évolutive du développement, l'évo-dévo est l'un des courants les plus dynamiques en biologie de l'évolution au début du XXIe siècle.
Cette discipline consiste à observer l’évolution du développement des organismes. Elle permet de comparer les processus développementaux des espèces et d’observer les liens phylogénétiques mis en place entre elles. En étudiant l’origine et l’évolution du développement embryonnaire, elle permet de comprendre comment les changements dans les processus développementaux rendent possibles l'apparition de nouveaux caractères ou de nouvelles structures à partir de gènes déjà existants. L'évo-dévo montre donc comment l’évolution peut altérer certains mécanismes biologiques et ainsi créer des nouveautés chez les espèces. De plus, l'homoplasie et l’homologie sont deux concepts importants qui permettent de retracer les liens entre les espèces en comparant l’apparition et la disparition de certains caractères chez leurs ancêtres communs.
La biologie évolutive du développement permet aussi de comprendre le rôle de la plasticité, c’est-à-dire l’influence qu'à l'environnement sur ces changements.
Grâce à l'étude de l'évo-dévo, il est possible d'avoir une meilleure compréhension des gènes chez différentes espèces et des mutations qui les affectent et d'observer l'impact de leur développement et de l'évolution en général. En observant notamment comment se développe l'Humain, de l'embryon à l'adulte, il est par exemple plus facile de comprendre comment les mutations et donc des maladies congénitales se développent et affectent notre santé.
Histoire
Cette discipline est née dans les dernières décennies du XXe siècle mais elle s'inscrit dans la continuité de l'embryologie comparative fondée au XIXe siècle.
Les théories développées au XIXe siècle
- Le principe de divergence selon Karl Ernst Von Baer (1792-1876) : Les caractéristiques générales communes à plusieurs taxons apparaissent prématurément chez l’embryon par rapport aux caractéristiques spécifique de chaque taxon[1]. Les patrons de développement montrent des similitudes initialement et divergent plus tard, durant le développement, pour donner des embryons très différents.
- L'embryon comme source d'homologies par Milne-Edwards (1800-1885) : Milne-Edwards orienta les morphologistes vers l’embryon en tant que source abondante d'homologies[2]. Si une homologie permet de retrouver les caractéristiques générales ou essentielles d’un organe ou d’un tissu alors en observant un embryon, on peut retrouver le caractère donné dans sa forme la plus générale, la moins différenciée. L’idée selon laquelle les caractères développementaux sont d’importants indicateurs des relations phylogénétiques prédomine encore de nos jours.
- Théorie de la récapitulation par Ernst Heinrich Haeckel (1834-1919) : Théorie selon laquelle l'ontogenèse récapitule la phylogenèse. Elle n’est pas vraie au sens strict. Le développement embryonnaire d’un organisme ne présente pas de stade équivalent aux différentes formes adultes de ses ancêtres. Mais un vertébré présente, lors de son développement embryonnaire, des stades ressemblant aux stades embryonnaires de ses ancêtres[3]. En effet, durant le développement d’un poulet, l’embryon ne passe pas à travers les stades cellule unique, ancêtre multicellulaire invertébré, poisson, lézard ancestral, oiseau ancestral puis finalement poussin ; mais il ressemble à l’embryon de poisson et reptile à certains points de son développement.
- La théorie de l'évolution par Charles Darwin (1809-1882) : Les espèces dérivent toutes d' ancêtres communs et c'est pourquoi toutes les espèces d'un même genre se ressemblent[4]. Ainsi un caractère est homologue dans deux taxons ou plus si on retrouve cette caractéristique chez l’ancêtre commun le plus proche de ces taxons. Depuis Darwin, l'homologue est l’expression de la continuité généalogique de l’information.
- La première théorie de l'évolution, ou Transformisme, de Jean-Baptiste de Lamarck (1744 - 1829), avec le principe de la transmission des caractères acquis depuis un ancêtre commun[5].
De 1900 à 1980
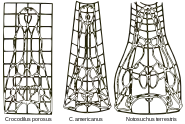
- La Théorie des transformations par D'Arcy Wentworth Thompson (1860-1948) :
Des transformations peuvent être appliquées à la forme d’une espèce (Figure 1 : des crocodiles) pour donner la forme d’autres espèces à l’intérieur d’un même groupe[6]. Les individus pris ici sont sous forme adulte. Les changements que l’on observe, sont dus à des changements tardifs dans l’ontogénie, lorsque la croissance allométrique est prépondérante. Thompson mit en avant le fait que les changements dans le développement au cours de l’évolution ne sont pas effectués par morceaux, dans le sens où chaque caractéristique de l’animal aurait évolué séparément, mais plutôt qu'ils sont coordonnés au sein de l’organisme.
- Gavin de Beer et l’hétérochronie (1899-1972) :
Il étudie l’embryologie expérimentale, comparative, descriptive et enfin évolutionnaire[7]. Notion centrale dans son œuvre Embryologie et Évolution (1930), l’hétérochronie tient d’après lui, un rôle prédominant dans l’évolution morphologique[7]. Définie comme un changement dans le timing des évènements développementaux, la notion est attribuée à Ernst Haeckel.

- Richard Goldschmidt et la modèle saltatoire (1878-1958) :
Selon Goldschmidt, l'évolution des espèces pourrait se produire par « saut », du passage d'une forme à une autre très différente et ce grâce à de petites mutations des gènes. Son travail sur les mutants homéotiques chez les Drosophila melanogaster influença certainement sa théorie. Chez les mutants homéotiques, une structure correcte apparaît à la mauvaise place: sur la figue 2, on peut observer le cas de la mutation antennapedia : présence d'une paire de patte à la place des antennes. Les études de Goldschmidt portaient surtout sur les mutant bithorax : le troisième segment thoracique devient une répétition du second, et les haltères sont remplacés par des ailes. Cependant, les mutants homéotiques connus de Drosophila melanogaster ne sont pas viables en comparaison du phénotype sauvage et le modèle de Goldschmidt n'est pas accepté aujourd'hui.
.jpg.webp)
- Conrad Hal Waddington et le paysage épigénétique (1905-1975) :
Il illustra son principe de canalisation[8] par le schéma d’un paysage épigénétique : dans la figure 3, les différents parcours de développement possibles d’une cellule (balle) sont représentés sous la forme d’un paysage dont les reliefs sont commandés par des gènes et leurs systèmes épigénétiques. C’est le même principe qui est soutenu par Waddington quant au développement d’un organisme d’une espèce données : (au départ) un embryon va évoluer en étant soumis à l’influence de l’environnement et des gènes jusqu’à un stade adulte (l’arrivée). Le degré de stabilité de la trajectoire est donné par la profondeur de la gouttière, par le degré de canalisation. La canalisation est une appréciation de la capacité de produire un phénotype constant en étant soumis à des effets environnementaux et/ou génétique et met en évidence la résistance des systèmes développementaux. Lorsque la perturbation est trop importante, une autre vallée est empruntée, on observe un changement de patron (repatterning). Waddington est aussi à l’origine de la découverte du processus d’assimilation génétique : un stimulus environnemental qui induit un phénotype particulier peut être remplacé par un facteur génétique[9].
L'avènement : les années 1980
- L'étude des mutations de gènes affectant le processus de développement chez Drosophila melanogaster au début des années 1980, fit passer la biologie évolutive du développement du stade expérimental et descriptif à un stade plus avancé traitant de génétique.
- La découverte de l'homéoboîte en 1984 par Walter Gehring, fut presque aussi importante en biologie que celle de la structure de l'ADN en 1953 par Watson et Crick. L'homéoboîte est une séquence de 180 paires de bases d'ADN, commune à certains gènes homéotiques (responsable du plan d'organisation) chez les animaux. Après transcription et traduction, elle donne une séquence de 60 acides aminés appelé homéodomaine, très conservé au cours de l'évolution (le reste de la protéine l'est généralement moins). Ce domaine donne des propriétés de liaison à la protéine auquel il appartient, lui permettant de remplir son rôle de régulation d'activité des gènes, de les activer ou de les désactiver. Ces protéines vont donc contrôler le développement de l’œuf à l'adulte. Le caractère ubiquiste de l'homéoboîte à travers le règne animal en fait une base commune du développement. Les gène HOX, responsables de la détermination de l'axe antero-postérieur du corps des animaux bilatériens contiennent cet homéoboîte. Une particularité de ces gènes est que leurs positions linéaires dans le complexe de gène d'un chromosome est identique à l'ordre antéro-postérieur de leur zone d'expression sur le corps de l'animal (phénomène de colinéarité).
- Chez les végétaux, c'est la famille des gènes MADS-box qui fait figure de base commune de développement.
Voir aussi
Articles connexes
- Accommodation génétique
- Accommodation phénotypique
- Acide rétinoïque
- Amphimedon queenslandica
- Assimilation génétique
- Atavisme
- Biais de développement
- Bicoïd (gène)
- Biominéralisation
- Biosignature
- BMP4
- Bois (cervidé)
- Boîte homéotique
- Canalisation
- Coadaptation
- Construction de niche
- Contrainte
- Cooptation
- Développement mosaïque
- Développement à régulation
- Développement des mycètes
- Écologie du développement
- Écotype
- Edwin Grant Conklin
- Effet Baldwin
- Embryologie expérimentale
- Épigenèse (biologie)
- Épigénétique chez les plantes
- Étoile de mer
- Évolvabilité
- Évolution chez les larves
- Évolution de la multicellularité
- Exaptation
- Explosion cambrienne
- Expression des gènes
- Gène engrailed
- Gènes de segmentation
- Gènes homéotiques chez les animaux
- Gènes Hox
- Gène HOXD13
- Gènes homéotiques chez les plantes à fleurs
- Hunchback (gène)
- Gènes KNOX
- Génétique
- Génétique évolutive du développement des plantes
- Génétique quantitative
- Guêpe parasitoïde
- Hedgehog
- Hétérochronie
- Hétérométrie
- Hétérotopie
- Hétérotypie
- Homoplasie
- Homologie (évolution)
- Induction embryonnaire
- Lignée cellulaire (développement)
- Loi de Dollo
- Loi de von Baer
- Modes de sélection naturelle
- Modularité biologique
- Morphogenèse
- Nouveauté évolutive
- Paysage adaptatif
- Phénocopie
- Phylogénétique
- Plan d'organisation
- Plasticité phénotypique des plantes
- Plasticité chez les plantes
- Règle de Gloger
- Remodelage du patron de développement
- Schistes de Burgess
- Séquence codante
- Séquence régulatrice
- Stade phylotypique
- Syndrome de domestication
- Système de polarité segmentaire
- Tétra aveugle
- Théorie de la récapitulation
- Théorie des histoires de vie
- Théorie du gène égoïste
- Trompe (Spandrel)
- Saccoglossus
- Variations du développement
Bibliographie
- (en) Wallace Arthur, Mechanisms of Morphological Evolution : a combined genetic, developmental and ecological approach, Chichester/New York/Brisbane etc., A Wiley Interscience Publication Series, , 275 p. (ISBN 0-471-90347-7), p. 291.
- Leo W.Buss, The Evolution of Individuality, Princeton Legacy Library, (ISBN 0-691-08468-8), p. 189.
- (en) Wallace Arthur, A Theory of the Evolution of Development, Chichester/New York/Brisbane etc., John Wiley & Sons Canada, , 94 p. (ISBN 0-471-91974-8), p. 94.
- (en) Keith Stewart Thompson, Morphogenesis and evolution, New York/Oxford, Oxford University Press, , 154 p. (ISBN 0-19-504912-8, présentation en ligne), p. 156.
- K.J MacNamara, Mickael McKinney, Heterochrony : The evolution of Ontogeny, Springer US, (ISBN 0-306-43638-8), p. 437.
- Brian K.Hall, Evolutionnary Developmental Biology, London/Weinheim/New York etc., Spinger, , 491 p. (ISBN 0-412-78580-3, présentation en ligne), p. 67.
- (en) Stanley N.Salthe, Devolpment and evolution : Complexity and change in Biology, Cambridge (Mass.)/London, The MIT Press, , 357 p. (ISBN 0-262-19335-3), p. 363.
- C.David Rollo, Phenotypes : Their Epigenetics, Ecology and Evolution, Nature, (ISSN 0028-0836), p. 398.
- Wallace Arthur, The origin of animal body plans : A Study in Evolutionary Developmental Biology, Cambridge University Press, , 357 p. (ISBN 0-521-55014-9), p. 343.
- Rudolf A. Raff, The Shape of Life : Genes, Development, and the Evolution of Animal Form, The University of Chicago Press, , 520 p. (ISBN 978-0-226-70266-7 et 0226702669).
- (en) Marc Kirschner et Jon Gerhart, Cells, embryo, and evolution, Malden (Mass.)/London/Paris, Wiley, , 642 p. (ISBN 0-86542-574-4), p. 642.
- (en) Carl Schlichting et Massimo Pigliucci, Morphogenesis and evolution, New York/Oxford, Sinauer, , 154 p. (ISBN 0-19-504912-8), p. 387.
- Scott D.Weatherbee Sean B.Carroll, Jennifer K.Grenier, From DNA to diversity : Molecular genetics and the evolution of animal design second edition, Blackwell Publishing, (ISBN 1-4051-1950-0), p. 258.
- Giuseppe Fusco Alessandro Minelli, Evolving Pathway : Key Themes in Evolutionary Thought, Cambridge University Press, , p. 126.
- Adam S.Wilkins, The evolution of developmental pathways, Sinauer, , p. 603.
- Marie-Jane West-Eberhard, Developmental Placticity and Evolution, Oxford University Press, (ISBN 0-19-512235-6), p. 797.
- (en) Alessandro Minelli, The development of animal form : Ontogeny, Morpholgy and Evolution, Cambridge etc., Cambridge University Press, , 323 p. (ISBN 0-521-80851-0), p. 323.
- (en) Wendy M. Olson et Brian K. Hall, Keywords and concepts in evolutionary developmental biology, Cambridge (Mass.), Harvard University Press, , 476 p. (ISBN 0-674-00904-5), p. 476.
- (en) Alessandro Minelli (trad. de l'italien), Forms of Becoming : The Evolutionary Biology of Development, Princeton (N.J.), Princeton University Press, , 228 p. (ISBN 978-0-691-13568-7, présentation en ligne).
- Jason Scott Robert, Embryology, epigenesis and evolution : Taking development serisouly, Cambridge University Press, (ISBN 978-0-511-18923-4), p. 158.
- Wallace Arthur, Biased embryo and evolution, Cambridge University Press, , 233 p. (ISBN 0-521-54161-1, présentation en ligne), p. 130.
- Sean B. Carroll, Endless Forms Most Beautiful : The New Science of Evo Devo and the Making of the Animal Kingdom, Norton, , 350 p. (ISBN 978-0-393-06016-4).
- Marc Kirschner et John Gerhart, The Plausibility of Life : Resolving Darwin's Dilemma, Yale University Press, , 314 p. (ISBN 978-0-300-10865-1).
- Ron Amundsun, The changing role in evolutionary thought, Cambridge University Press, (ISSN 0018-1196), p. 280.
- Jane Maienschein et Manfred Dietrich Laubichler, From embryology to Evo-Devo : a history of developmental biology, MIT Press, (ISBN 978-0-262-12283-2), p. 569.
- (en) Alessandro Minelli (trad. de l'italien), Forms of becoming : The evolutionary of development, Princeton (N.J.), Princeton University Press, , 228 p. (ISBN 978-0-691-13568-7, présentation en ligne), p. 233.
- (en) David Epel et Scott F. Gilbert, Ecological developmental biology : integrating epigenetics, medicine, and evolution, Sunderland, Sinauer, , 480 p. (ISBN 978-0-87893-299-3), p. 480.
- Wallace Arthur, Evolution : a developmental approach, Wiley-Blackwell, , 416 p. (ISBN 978-1-4051-8658-2), p. 480.
- Journal Evo-Devo
- Journal Developments, genes and evolution
- Journal Molecular and developmental Evolution
- Journal Evolution & Development
Notes et références
- Pour une discussion générale du cadre conceptuel dans lequel s'inscrit la biologie évolutive du développement, on peut se reporter à l'ouvrage de Stephen Jay Gould, La Structure de la théorie de l'évolution (Belknap/Harvard 2002) et en particulier au chapitre 10.
- Jacques Berthet et Alain Amar-Costesec, Dictionnaire de Biologie, de Boeck, 2006, 1034 p.
- Toby A Appel, The Cuvier-Geoffroy debate : French biology in the decades before Darwin, New York : Oxford University Press, 1987, 219 p.
- Peter H.Raven and George B.Johnson, Biology, Raven, 6th edition 2002, 1229 p.
- Charles Darwin, L'origine des espèces,Librairie François Maspero, 1980, 169p.
- Lamarck J-B., Recherches sur l’organisation des corps vivants et particulièrement sur son origine, sur la cause de ses developpemens et des progrès de sa composition., Paris, "Chez l’Auteur au Muséum d’Histoire Naturelle",
- D'Arcy Wentworth Thompson,On Growth and form, Cambridge University Press, 1917, p 724 725 727 744 753
- Ingo Brigandt, Homology and Heterochrony : The Evolutionary Embryologist Gavin Rylands de Beer (1899-1972), Journal of Experimental Zoology (Part B : Molecular and Developmental Evolution) 306B : 317-328, 2006, p 4 5
- C.H.Waddington, « Canalization of development and the inheritance of acquired characters », Nature, n°3811, (1942), pp 563-565
- Paul-Antoine Miquel, "Biologie du XXIe siècle : évolution des concepts fondateurs", De Boeck, (2008), pp 145-153
Liens externes
- (en) Lancelet and Ancestral Body (article : évolution des chordés, patrimoine génétique, découvertes récentes) - PENICHEFOSSIL
 Portail origine et évolution du vivant
Portail origine et évolution du vivant