Liang Bua
La grotte de Liang Bua est un complexe cavernicole tributaire d'un réseau de chutes d'eau successives ayant creusé à flanc de paroi rocheuse une vaste cavité. Ce site naturel souterrain est situé sur l'île de Florès dans la province de Nusa Tenggara oriental en Indonésie. Par ailleurs la cavité se localise à proximité de la petite localité indonésienne éponyme Liang Bua. Son appartenance à la formation volcanique de l'île de Florès[N 2] soumet sa datation à l'Oligocène final / Miocène moyen sur l'échelle des temps géologiques et ce à l'instar du complexe géologique sub-océanique : l'arc insulaire de la Sonde[N 3]. Ce dernier observe un caractère de faille de subduction dite cisaillante, scindant et séparant les plaques tectoniques des Philippines d'une part, de la plaque australienne d'autre part et enfin de la plaque indo-océanique de tierce-part. Le site archéologique grotte de Liang Bua se caractérise notamment par les récentes mises au jour d'une nouvelle espèce du genre Homo : l'Homme de Florès ou Homo floresiensis, mais également d'un spécimen de Stegodon trigonocephalus florensis, un type d'éléphant nain préhistorique.
| |||||||||||||||||||||||||||||||||||||

.svg.png.webp)
Cadre géographique
Un contexte global de convergence lithosphérique
Le site archéologique de Liang Bua est localisé sur l'île de Florès appartenant à l'ensemble insulaire des Petites îles de la Sonde[N 4]. Cette zone géographique s'inscrit dans l'ensemble lithosphérique correspondant à la micro-plaque océanique de Timor[9],[10]. En outre, celle-ci est pourvue d'un angle solide[N 5] de 0,008 70 sr (le symbole sr pour stéradian)[N 6]. En termes purement géographiques, cette donnée correspond à une superficie d'environ 110 000 km2[11]. On détermine que la petite plaque océano-lithosphérique résulterait du morcellement de l'ancien sous-continent dit de Sunda[N 7]. À cet effet, l'estimation chronologique de la scission de cette terre pan-océanique est à imputer à l'ère géologique du Pliocène supérieur à Pléistocène inférieur (ou ancien)[12],[13],[10],[14].
Cependant on peut objecter que l'orogenèse succédant à la surrection des vestiges constituant les terres de Sunda fut effective au cours de l'Oligocène supérieur et se conclut du Miocène inférieur à moyen[15],[13],[9],[10],[14]. Sous cet angle, on détermine que la plaque eurasienne, la plaque philippine, la plaque de Sunda et enfin la plaque australienne ne formaient qu'une seule et même entité continentale[11],[12],[16],[13],[9]. Ce continent pan-australo-eurasien se développait en un vaste arc de cercle prenant appui de la partie occidentale de l'hémisphère Nord, jusqu'à la partie orientale de l'hémisphère Sud[12],[17],[13],[9],[18],[19]. Concrètement, cet élément signifie que la micro-plaque de Timor (et donc la terre de Florès) était géographiquement reliée à la plaque continentale eurasienne, il y a 2,5 Ma ; et ce, jusqu'au morcellement tectonique du sous-continent de Sunda, il y a approximativement 900 000 à 700 000 années[13],[9],[18],[19].
La micro-plaque de Timor s'insère au centre d'un nœud de contact lithosphérique : celle-ci est encadrée par la plaque continentale de l'Australie, au Sud ; la plaque de la mer de Banda, au Nord ; et la plaque océano-continentale de la Sonde, à l'Est[20],[13],[9],[18],[11],[14].
Géographiquement, les zones de contact lithosphérique de la plaque de Timor entre la plaque de la mer de Banda, d'une part, et la plaque de la Sonde, d'autre part, sont à l'origine géologique de la dorsale sub-océanique dit arc insulaire de la Sonde[14],[13],[9]. Ce dernier observe un vaste réseau de subductions longitudinales à caractère cisaillant et tangent à l'île de Florès[16],[19],[9],[13]. Par conséquent, la dorsale océanique de la Sonde impacte de façon directe l'îlot indonésien, celui-ci créant en son sein un volcanisme actif patent[16],[9],[19].
- Cartes de représentation de la zone lithosphérique indo-océanique.
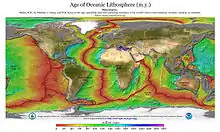 Carte chrono-lithosphérique des différentes plaques océaniques du globe terrestre. On peut remarquer que la petite plaque de Timor est relativement récente, en regard des autres plaques lithosphériques actuelles.
Carte chrono-lithosphérique des différentes plaques océaniques du globe terrestre. On peut remarquer que la petite plaque de Timor est relativement récente, en regard des autres plaques lithosphériques actuelles.
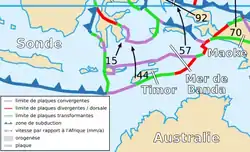 La plaque de Timor et ses différents contacts lithosphériques.
La plaque de Timor et ses différents contacts lithosphériques.
Un environnement régional d'isolement insulaire
Sous l'optique régionale, on peut observer que la grotte de id est sise dans la partie orientale de l'Indonésie : la province des Petites îles de la Sonde orientales[N 8], sur l'île de Florès. La petite terre volcanique de 302 kilomètres de long sur 10 à 12 kilomètres[3], évolue entre 8° et 9° de latitude Sud et 120° et 123° de longitude Est[21].
L'île à caractère volcanique est en outre baignée dans l'océan Indien et bordée dans sa partie méridionale par la mer de Timor[22],[23],[1],[21],[3].
- Situation géographique de l'île de Florès et de Liang Bua.

 Localisation de la province indonésienne des Petites îles de la Sonde orientales en Océanie. Ici représentées en couleur verte.
Localisation de la province indonésienne des Petites îles de la Sonde orientales en Océanie. Ici représentées en couleur verte. Localisation géographique de l'île de Florès. On peut la distinguer en haut, à gauche de l'ensemble régional des Petites îles de la Sonde orientales.
Localisation géographique de l'île de Florès. On peut la distinguer en haut, à gauche de l'ensemble régional des Petites îles de la Sonde orientales. Le village de Liang Bua se situe au centre du Kabupaten de Manggarai illustré en rouge sur la carte.
Le village de Liang Bua se situe au centre du Kabupaten de Manggarai illustré en rouge sur la carte.
Un cadre géographique local de volcanisme actif
D'un point de vue strictement local, la grotte de Liang Bua se situe au cœur Kabupaten de Manggarai[N 9], dans la partie occidentale de l'île de Florès. D'autre part, une structure rocheuse souterraine se déploie dans l'ouest du village éponyme Liang Bua, lequel est situé à environ 30 kilomètres au nord-ouest de Ruteng, chef-lieu du Kabupaten de Manggarai[24],[1].
Par ailleurs, la cavité de Liang Bua est assujettie à un petit ruisseau, lequel conflue à la rivière occidento-septentrionale de l'île de Florès, Wai Pesi[1],[22],[3].
Enfin, le site naturel est tributaire de deux volcans de type actif et d'un volcan dit éteint[N 10],[25]. se situant à son voisinage proche[N 11] et dont il est le barycentre : le Ranakah ; le Inierie ; et le Gunung Poco Leok, un stratovolcan à manifestations éruptives structurant sa composition rocheuse en andésites de type pyroxène[26],[27],[3],[25].
- Les trois volcans avoisinant la Grotte de Liang Bua.
 Le "Gunung Poko Leok"[N 12] est pourvu d'un point culminant à 1 625 mètres d'altitude. Il s'agit d'une montagne magmatique de type stratovolcan[N 13]. Ce genre de dôme volcanique est également appelé volcan composite.
Le "Gunung Poko Leok"[N 12] est pourvu d'un point culminant à 1 625 mètres d'altitude. Il s'agit d'une montagne magmatique de type stratovolcan[N 13]. Ce genre de dôme volcanique est également appelé volcan composite. Le "Gunung Ranakah" (altitude : 2 350 mètres) est un volcan de type volcan gris. Il se caractérise comme étant un dôme de lave[N 14].
Le "Gunung Ranakah" (altitude : 2 350 mètres) est un volcan de type volcan gris. Il se caractérise comme étant un dôme de lave[N 14]. Le "Gunung Inierie", présente également les caractéristiques d'un volcan gris[N 15]. Les analyses géologiques et sismologiques ont révélé que l'activité éruptive du volcan est demeurée nulle sur une longue période. Il s'agit d'un volcan dit éteint. Il culmine à 2 245 mètres.
Le "Gunung Inierie", présente également les caractéristiques d'un volcan gris[N 15]. Les analyses géologiques et sismologiques ont révélé que l'activité éruptive du volcan est demeurée nulle sur une longue période. Il s'agit d'un volcan dit éteint. Il culmine à 2 245 mètres.
.
Faune de la caverne
Environnement écologique
Le panorama global taxonomique de la grotte de Liang Bua bénéficie d'un assemblage faunique et végétal remarquable : la nature insulaire[N 16], volcanique et climatique de son environnement géographique proche de la ligne dite de Wallace[28], lui confère un caractère endémique significatif : on évalue que la terre insulaire de Florès possède une proportion effective de taxons spéciacisés d'environ 5,1 % (soit un nombre de 46 occurrences toutes espèces animales et végétales confondues)[29],[30],[31],[32]. Par ailleurs, la proximité équatoriale de l'île de Florès[N 17],[33],[34] et un régime climatique de moussons, induisent une biodiversité des espèces patente[35],[29],[30]. Dans les faits, on attribue la petite terre indonésienne de Florès, et par conséquent la grotte de Liang Bua, comme appartenant à l'ensemble biogéographique Wallacea. Cette dernière regroupe les trois aires géographiques de l'île de Sulawesi, des Moluques et des Petites îles de la Sonde[32],[35],[29].
Principaux taxons de la faune cavernicole
La cavité naturelle de Liang Bua affecte l'ensemble des critères propres à ceux d'une niche écologique[36],[34]. Toutefois, en raison de ses caractéristiques cavernicoles et donc pauvres en ressources alimentaires, le site souterrain oppose un contraste indéniable au sein de la petite insularité de Florès[37].
Pour autant, le site spéléologique de Liang Bua observe un cheptel de spécimens animaliers pourvu d'une remarquable biodiversité phylogénétique. De nombreuses araignées aranéomorphes sont connues de ce site, et notamment Tetrablemma manggarai[38],[39]. En outre, la grotte est également dotée d'un environnement écologique composé de plusieurs espèces de Murinae (de petits rongeurs proches des souris), ou de Psittacidae tels que le Loriquet de Weber (Trichoglossus weberi), proche parent du Loriquet à tête bleue (T. haematodus)[37]. La caverne observe également une notable occupation écologique de Petit-duc de Florès (Otus alfredi), hibou endémique de l'île de Florès[40],[41],[42],[43].
La grotte abrite une petite population de primates, essentiellement des orang-outans. Ces derniers occupent le site spéléologique à l'équivalence d'un habitat écologique et à la faveur d'une tierce faune relativement peu nombreuse et dont la dominance territoriale est significativement restreinte[44],[37].
- Espèces endémiques de la grotte et ses environs immédiats.
 Loriquet de Weber (Trichoglossus weberi).
Loriquet de Weber (Trichoglossus weberi).
 Oreophryne rookmaakeri, amphibien endémique observable dans la grotte de Liang Bua[46]
Oreophryne rookmaakeri, amphibien endémique observable dans la grotte de Liang Bua[46] }}
}}
Contexte chrono-géologique
La formation des roches structurantes du site est étalonnée à deux phases. D'une part, une première phase se développant à l'ère géologique de l'Oligocène supérieur (autrement dit, la subdivision stratigraphique du Chattien) et correspondant à la genèse géologique de l'île de Florès, entre −28,10 et −23,04 Ma[N 18],[47],[48],[49],[15],[50],[51]. Du point de vue paléogéographique, cette première étape de formation rocheuse est contemporaine de la dérive d'axe Sud/Sud-Est de la plaque australienne[50],[47],[51].
D'autre part, un second cycle de formation orogénique attribué au Miocène moyen à Miocène supérieur et correspondant aux subdivisions stratigraphiques du Serravallien et du Tortonien, soit en termes chronostratigraphiques : entre −13,82 et −7,246 Ma[51],[47],[15],[48],[52],[53]. Par ailleurs, cette période géologique se corrèle avec l'entrée en collision de la plaque indienne et de la plaque eurasienne, simultanément à l'interaction tectonique de la plaque australienne et de la plaque sud-pacifique. Ces deux dernières évolutions tectoniques engendrent ainsi l'arc insulaire des Petites îles de la Sonde orientales[47],[51],[53].
L'orogenèse des structures géologiques porteuses du site s'échelonne donc au sein d'une fourchette chronostratigrahique allant de −28,10 à −7,246 Ma[51],[15],[47],[53].
- Perspective de la formation de la grotte dans les temps géologiques.
 Oligocène final / Miocène moyen : La genèse de l'île de Florès se situe en haut à gauche de l'échelle des temps géologiques.
Oligocène final / Miocène moyen : La genèse de l'île de Florès se situe en haut à gauche de l'échelle des temps géologiques.
Dans un second temps, on atteste également de dépôts latéraux à caractère volcanique et/ou sédimentaires. Ces derniers possèdent un déterminant hétérochrone lié aux formes d'adaptations biologiques de la microfaune océanique indonésienne du Miocène moyen[16],[19],[54],[55]. Toutefois, les parois de la grotte, ainsi que ses couches sédimentaires pédologiques secondaires, observent également des formations de type turbidique ; mais aussi des microgranules de calcaire coquillier tel que la lumachelle, l'ensemble se prévalant de la périodechronostratigraphique du Miocène moyen à supérieur[19],[54],[56].
Enfin, on peut appréhender la présence de particules de granodiorite[N 19] essentiellement constituées de feldspaths et de quartz. Ces dépôts stratigraphiques affectent notamment les couches sédimentaires primaires du site spéléologique. Ces derniers sont contemporains d'un cycle géologique assigné au Pliocène moyen à supérieur[N 20] et correspondant aux reflux océanique sur la mosaïque de l'ancien sous-continent pan-océanique dit de Sunda, d'une part, et la résurgence tecto-volcanique des terres composant les Petites îles de la Sonde orientales, d'autre part[16],[19],[56],[57].
- Les différentes formations sédimentaires de la grotte de Liang Bua.
 Un feldspath.
Un feldspath. Une lumachelle, agglomérat de calcaire à dominante coquilleuse.
Une lumachelle, agglomérat de calcaire à dominante coquilleuse.
Description
Structure géologique de la grotte
La caverne de Liang Bua prédispose d'une infrastructure latérale et sub-pédologique composée de roches engendrées à l'Oligocène supérieur/Miocène moyen affichant des substrats essentiellement de type granodioritiques, tels que le quartz et le pyroxène ; et magmatique, tel que le basalte tholéiitique[58],[59],[60],[61],[62],[5].
- Les types de roches formant la caverne de Liang Bua.


 Exemple de basalte de type tholéiitique. Ici : une olivine provenant du piton de la Fournaise, sur l'île de La Réunion.
Exemple de basalte de type tholéiitique. Ici : une olivine provenant du piton de la Fournaise, sur l'île de La Réunion.
Les premières couches sédimentaires du sol présentent une nature essentiellement basaltique et dans une moindre mesure de type grès micro-granuliforme induit par l'érosion des roches structurantes de la cavité souterraine, mais également par l'incidence du facteur éolien sur ces dernières. La surface pédologique du complexe spéléologique se ponctue également par de nombreuses accrétions à caractère magmatique et dont la teneur géologique est comparable à celle des roches encadrant l'espace cavernicole[58],[59],[60],[61],[63].
La voûte du complexe souterrain, en prise directe des formations minérales porteuses, est quant à elle constituée de volumineux stalactites basaltiques, et de par leur abondance, dessinent un panorama chaotique. Ces derniers sont déterminés par deux facteurs concomitant. Premièrement on observe un processus de cristallisation des particules magmatiques contenues dans le ruissellement de l'humidité ambiante[N 21]. Deuxièmement, on atteste d'un phénomène d'épanchement des laves multimillénaires : sous cet angle, la grotte de Liang Bua peut être appréhendée comme un tunnel de lave[63],[61],[55],[64].
- Exemples types d'un tunnel volcanique et d'une à stalactites basaltique.
 Ici : le tunnel de lave de Cuevas, en Amérique centrale.
Ici : le tunnel de lave de Cuevas, en Amérique centrale.
 Lucarne de tunnel de lave.
Lucarne de tunnel de lave.
Architecture de la grotte
Le site cavernicole se présente sous la forme d'une creusée à flanc de roche tributaire de l'abondante pluviosité propre à l'île de Florès[6], mais également d'un glissement de terrain de nature sismique datant du Miocène moyen[65]. La cavité souterraine affiche de nombreuses concrétions[N 22] dont essentiellement des stalactites majoritairement pourvues d'une taille conséquente. Dans une moindre mesure, l'aspect de la grotte est affecté par un réseau hydrographique de chutes d'eau d'un petit ruisseau[N 23],[3],[65].
L'architecture du complexe cavernicole s'organise autour d'un vaste espace d'une longueur d'environ 50 mètres et d'une largeur comprise entre 6 et 11 mètres, couvrant ainsi une surface au sol d'approximativement 400 m2[63]. La structure spéléologique évolue selon une hauteur au plafond allant de 10 à 15 mètres[63]. Par ailleurs, la cavité souterraine offre un seul et unique moyen de pénétration piétonnier, mais qui néanmoins affecte de généreuses dimensions. Cet élément d'entrée confère à la grotte de Liang Bua une remarquable facilité d'accès[60],[66],[N 24].
Historique du site archéologique
L'inauguration du site
L'inauguration de la caverne et son indexation au statut de site archéologique national du patrimoine indonésien prirent effet au cours des années 1950[67],[68]. Dans un premier temps, les archéologues de l'époque réalisèrent des extractions taphonomiques relativement mineures en regard des découvertes effectuées dans les années 1990[69],[67],[68].
Première découverte : Stegodon orientalis
Au tournant des années 1990 / années 2000, dans le cadre d'un programme de recherches archéo-paléontologiques au sein des Petites îles de la Sonde orientales à l'ère du Pléistocène, deux équipes d'archéologues[N 25],[70],[1],[68] mettent au jour des vestiges fossilisés de type osseux appartenant à la famille des Stegodontidae, Stegodon florensis insularis[N 26],[71],[72],[68],[N 27]. L'individu paléontologique était incorporé aux premières couches stratigraphiques de la grotte de Liang Bua[1],[73],[74],[68]. Par l'obtention d'une analyse au (14C), les deux équipes de chercheurs indexent les restes osseux[N 28] du pachyderme nain[N 29],[75],[76] dans une fourchette chronologique comprise entre −75 000 et −30 000[1],[70],[77],[72],[68],[71]. Par conséquent, on détermine que ce dernier aurait vécu à la subdivision chrono-géologique du Pléistocène supérieur[71],[78],[1],[70],[77],[74],[79],[72],[68].
- Reconstitution informatisée et vitrines d'exposition d'un Stégodon.
 Reconstitution par ordinateur d'un Stegodontidae.
Reconstitution par ordinateur d'un Stegodontidae. Boîte crânienne pourvue de ses défenses d'un Stegodontidae exposée et conservée au Museum of natural history de Londres.
Boîte crânienne pourvue de ses défenses d'un Stegodontidae exposée et conservée au Museum of natural history de Londres.
 Molaire provenant d'un Stegodon orientalis.
Molaire provenant d'un Stegodon orientalis.
Seconde découverte : un crâne humain
En 2003, une équipe d'archéologues dirigée par Mike Morwood[N 30] et Radien P. Soejono[N 31] entreprend des recherches systématiques au sein des couches stratigraphiques primaires[N 32] sur le site naturel souterrain du village de Liang Bua. Ces dernières mettent au jour un crâne fossilisé. Une analyse, effectuée par technique de datation au (14C), permet d'attribuer son origine chronologique comprise entre -90 000 et -50 000 années : en terminologie paléontologique, il s'agit donc de la subdivision chrono-géologique du Pléistocène supérieur[15],[80],[24]. Le crâne extrait des premières couches sédimentaires affiche un volume variant entre 380 cm3 et 426 cm3[N 33],[66],[70],[80]. Pour comparaison, celui d'un homme moderne affiche un volume de structure crânienne d'environ 1 350 cm3, soit un rapport de 3 à 3,5 fois plus imposant[70],[80].
- Le fossile du crâne découvert dans la grotte.
_01.jpg.webp) La boîte crânienne mise au jour en 2003, par l'équipe d'archéologues sous la houlette de Mike Morwood et Radien P. Soejeno.
La boîte crânienne mise au jour en 2003, par l'équipe d'archéologues sous la houlette de Mike Morwood et Radien P. Soejeno.
2003 : Autres fouilles et troisième mise au jour
Quelques mois en aval de la seconde découverte, les mêmes intervenants effectuent des fouilles plus approfondies des couches stratigraphiques sédimentaires superficielles du sol spéléologique : ils mettent au jour la quasi totalité d'un squelette fossile, à l'exception de la boîte crânienne[81],[82]. L'obtention de deux études, la première par le biais d'une datation par le carbone 14, et la seconde par analyse au microscope électronique, confirment la corrélation entre le crâne et l'ensemble osseux : de facto, il s'agit d'un seul et même individu[1],[80],[83].
Par ailleurs, l'identification du bassin révèle de manière indubitable deux éléments essentiels :
- il s'agit d'un individu de sexe féminin[1],[24],[80] ;
- le bassin du spécimen LB1 témoigne d'une bipédie indubitable[1],[24],[80].
Cette dernière donnée impose aux chercheurs paléontologues que les vestiges osseux appartenaient à un Hominina[1],[73],[80].
En marge de ces découvertes majeures, l'équipe de paléontologues parvient à identifier des traces de combustible végétal[N 34] et daté entre −90 000 et −45 000 années[1],[73],[80],[84],[85]. En outre, des objets lithiques tels que des silex de type biface ou encore des nucléus, sont également identifiés et répertoriés. Il s'agit d'industries lithiques taillées dans une roche basaltique, laquelle a été induite par la formation géologique dite de Ola Bula[71]. Une estimation chronologique fournit un indice approximatif de leur réalisation : les éclats correspondraient à la période du Pléistocène moyen à supérieur[71].
Sous l'angle de ces derniers éléments, les spécialistes corroborent leur prime postulat : le spécimen extrait possédait très probablement toutes les caractéristiques évolutives propres aux espèces appartenant à l'embranchement phylogénétique des hominina[70],[73],[74],[86],[87].
L'ensemble des éléments identifiés sont baptisés Flora, en écho au nom de l'île sur lesquels ils ont été retrouvés : Florès. En outre, la dénomination binomiale dont ces derniers sont octroyés, se concrétise par Homo floresensis. Enfin, la terminologie du spécimen (ou holotype se matérialise par LB1[73],[88] ,[87],[80].
- Le squelette de Flora.
 Reconstitution de l'ensemble des éléments du squelette de Flora, Homo floresiensis.
Reconstitution de l'ensemble des éléments du squelette de Flora, Homo floresiensis.
Quatrième découverte : les autres fossiles humains
En 2005, une troisième session de fouilles s'organise à Liang Bua. Toujours sous la direction de Mike Morwood et Peter Brown, les archéologues réalisent des examens systématiques plus approfondis des couches stratigraphiques secondaires[N 35]. Les chercheurs mettent au jour un second individu fossile[89],[70].
Ultérieurement, huit autres individus sont découverts, portant le total à dix individus attribués à l'Homme de Florès[24],[70],[74],[89].
Cinquième découverte au sein de la grotte de Liang Bua : Leptoptilos robustus
Dans une optique d'appréhender l'ensemble des répercussions paléontologiques induites par ces nouvelles données, Mike Morwood et Peter Brown, pérennisent leurs fouilles archéologiques au sein de la caverne de Liang Bua au cours de l'année 2007[90]. Incidemment, ils mettent au jour une nouvelle occurrence de fossiles, un marabout fossile qui sera décrit sous le nom de Leptoptilos robustus[8],[90]. Cette espèce a un bec long et massif, de longues pattes et présente une corpulence morphologique et anatomique remarquable : 1,80 mètre de long et un poids estimé à environ 16 kilogrammes[8],[69]. Cet échassier aurait très probablement existé au Pléistocène moyen et supérieur, soit une période se déployant entre −258 000 et 50 000 années[8],[91],[15]. Néanmoins, une estimation effectuée par le biais d'une analyse au (14C), fournit une indication de datation plus précise : les fossiles de L. robustus découverts dans la grotte de Liang Bua s'échelonneraient entre −110 000 et −50 000[92].
Concrètement, cette période chevauche celle affichée par les fossiles d'Homo floresensis[N 36].
Par conséquent, ces dernières données obtenues témoigneraient d'une probable cohabitation entre les deux taxons préhistoriques au sein de la niche écologique souterraine incarnée par la caverne de Liang Bua[92],[93],[94],[73],[N 37],[95].
- Le Leptoptilos robustus.
 Reconstitution d'un Leptoptilos robustus.
Reconstitution d'un Leptoptilos robustus. Schéma comparatif de la taille d'un Leptopilos robustus avec celle d'un humain.
Schéma comparatif de la taille d'un Leptopilos robustus avec celle d'un humain.
La caverne de Liang Bua : une niche écologique au Pléistocène moyen et Pléistocène supérieur
L'ensemble taphonomique du site
Le cadre taphonomique des strates sub-pédologiques du site affecte d'abondantes ressources archéologiques. En parallèle des découvertes de fossiles de Stegodon florensis insularis, Homo floresiensis, et de Leptoptilos robustus, les paléontologues ont également mis au jour des ossements appartenant à quatre autres types d'espèces.
Les archéo-paléontologues en ont actuellement attesté et répertorié trois à quatre substrats purement fauniques.
On atteste ainsi de spécimens de bovidés, lesquels sont pourvus de liens phylogénétiques avec l'actuel Anoa de Malaisie de manière patente. La physionomie globale des vestiges fossilisés de ces ruminants suggère que l'élément pathologique d'insularisme aurait affecté ces derniers. Leur extraction a été réalisée au cours de l'année 2002.
Par ailleurs, on a extrait des restes fossilisés de mammifères à phénotype carnivore, dont notamment des tigres à dents de sabre[N 38],[74],[68],[73],[82],[70]. En outre, des restes de deux occurrences taxonomiques de murinés furent découverts au sein de la caverne en 2004. Leur identification taxonomique les subrogea au Papagomys armandvillei, d'une part, et au Papagomys theodorverhoeveni, d'autre part[N 39]. À contrario de l'Homme de Florès et des occurrences de bovidés, ceux-ci présentent une pathologie morphologique de gigantisme. Cette particularité anatomique procède du même déterminant hétérochrone, lequel est dû à un schéma géographique d'endémisme insulaire. En d'autres termes, ils auraient subi une forme d'adaptation évolutive en phase finale de leur développement : il s'agit de cas d'hypermorphose[96],[97],[98],[73],[1],[80],[99].
D'autre part, le sous-sol cavernicole recelait également des traces d'espèces de type lophozoaires, dont notamment des eumollusca. Ces extractions de dépôts fauniques d'origine océanique viennent corréler la scission orogénique du sous-continent pan-océano-eurasiatique dit Sunda aux alentours de −800 000 ans[12],[17],[16],[13],[9]. Cet élément témoignerait ainsi de l'émergence volcano-géologique de l'île de Florès. Dans ce même cadre, celui-ci pourrait également suggérer un impact significatif d'ordre orogénique en prise directe avec les couches sédimentaires de la caverne[55],[13],[18],[16],[19]. Les spécimens sous-marins lophozoaires ont été mis au jour en 1978 par l'archéologue indonésien Rokus Awe Due[100].
- Exemple de Lophozoa : la seiche.

- Exemples de tigres à dents de sabre.
 Homotherium crenatidiens crenatidens fabrini.
Homotherium crenatidiens crenatidens fabrini. Exposition au Hall Paléolithique du Musée d'histoire naturelle de Londres.
Exposition au Hall Paléolithique du Musée d'histoire naturelle de Londres.
- Mâchoires de Papagomys (ou rats géants) provenant de l'île de Florès.
 Papagomys d'Armandville et de Verœvehen.
Papagomys d'Armandville et de Verœvehen.
L'équipe de paléontologues menée par Mike Morwood et Peter Brown a extrait en 2014 des fossiles humains datés de 700 000 ans qui présentent certaines similitudes avec l'Homme de Florès, sur le site de Mata Menge, dans le kabupaten de Sikka, partie orientale de l'Florès, à 50 km de Liang Bua. Ces fossiles pourraient représenter les ancêtres de l'Homme de Florès[101].
Au plus fort des périodes glaciaires successives, Florès n'était séparée du Sunda que par un modeste détroit[24],[73],[102],[103],[104],[105],[83].
Globalement, l'ensemble de la documentation archéologique de la grotte de Liang Bua est assignée à l'époque géologique dite du Pléistocène supérieur[74],[68],[73],[82],[70],[8]. Concrètement, ces indices confirmeraient un probable processus d'enveloppement écologique au sein la caverne de Florès : différentes espèces taxonomiques préhistoriques auraient préempté le site naturel souterrain comme lieu d'habitat. De facto, celui-ci s’inscrit au cœur d'un milieu hostile induit par un vecteur de volcanisme notable, et ce depuis l'orogenèse de l'île de Florès à l'Oligocène supérieur[51],[47],[21],[7],[106]. Sous cet angle, la caverne de Liang Bua semblerait constituer un site à caractère écologique, dont l'indexation chronologique s’échelonnerait sur la période du Pléistocène final[74],[68],[73],[82],[70],[8].
Le cas de l'Homme de Florès
- Études et analyses de l'homme de Florès
 On peut observer la faiblesse de l'angle formé par le sinus ethmoïdal, ainsi que les légers bourrelets des os pariétaux (partie supérieure-arrière de la calotte crânienne).
On peut observer la faiblesse de l'angle formé par le sinus ethmoïdal, ainsi que les légers bourrelets des os pariétaux (partie supérieure-arrière de la calotte crânienne).

Notes et références
Notes
- Essentiellement des concrétions de type stalactite.
- Comme en atteste le co-voisinage de la grotte de Liang Bua avec le volcan actif de l'île de Florès.
- Également appelé arc de Kora.
- Autrement appelé Nusa Tenggara, en langue indonésienne.
- L'angle solide se définit comme étant la surface terrestre obtenue sous le biais de la projection de l'angle d'un point par rapport au rayon terrestre.
- À titre comparatif, la plaque continentale africaine est dotée d'un angle solide d'approximativement 1,440 7 sr.
- Le terme Sunda se traduit en français par le terme Sonde.
- Autrement dénommé Nusa Tenggarra Timur, en langue indonésienne.
- Le Kabupaten est une unité administrative indonésienne subordonnée à la province indonésienne.
- À ce titre, l'île de Florès manifeste la densité volcanique la plus importante sur l'ensemble des terres indonésiennes.
- Sur des distances respectives couvrant 20 à 25 kilomètres.
- En indonésien, le terme "Gunung" signifie volcan.
- C'est-à-dire un volcan affectant une forme conique et doté de plusieurs strates de coulées de lave.
- Il présente des coulées de lave basaltique à flanc de paroi et dont la viscosité est à l'état solide. Ce phénomène est en incidence de son altitude.
- Autrement dit, de type explosif, et dont les laves sont de nature andésiques.
- Et donc isolée.
- C'est-à-dire située entre 8° et 9° de latitude sud.
- La terminologie Ma signifie millions d'années.
- Les granodiorites sont des roches dites plutoniques à dominante magmatique.
- C'est-à-dire dans une fourchette chronologique comprise entre 4 Ma et 900 000 années.
- Ledit processus est lié au goutte-à-goutte
- C'est-à-dire une accumulation des sels de type minéral et/ou calcaire contenus dans les eaux de précipitations et d'humidité.
- Lequel se jette dans le fleuve "Wai Pesi", dont le delta est situé au Nord du Kabupaten de Manggarai témoignant d'un processus d'érosion de la roche.
- Confere la page du numéro d'Archéologia concerné.
- Celle de Mike Morwood d'une part, et celle de Gert van den Beert et Paul Sonaar, d'autre part.
- Nom scientifique de l'espèce en terminologie dite binominale.
- Il s'agit d'un genre d'éléphant d'une pathologie morphologique de nanisme due à l'insularité, et dont on attribue l'époque d'existence au Pliocène et Pléistocène et endémique aux Petites îles de la Sonde orientales.
- Dont notamment les défenses.
- Il s'agit d'un type d'adaptation morpho-pathologique dû à deux facteurs simultanés : une paupérisation en ressources alimentaires d'une part, et une faiblesse du brassage génétique inféodé à de très fréquents cas de consanguinité, d'autre part. En d'autres termes, il s'agit de spéciation allopatrique.
- Éminent spécialiste en archéologie rattaché à l'Université de Wollongong, dans l'État de Nouvelle-Galles du Sud, en Australie.
- Éminent archéologue rattaché au Centre Indonésien pour l'Archéologie de Jakarta.
- Couches stratigraphiques en surface.
- Selon les sources bibliographiques.
- Très probablement du bois.
- Les couches du sol au-dessous des couches de surfaces.
- Pour rappel : entre −90 000 et −50 000.
- Voir photo page 11 de l'article ici mentionné.
- Plus précisément des Hemimachairodus, espèce endémique à l'Indonésie au cours de l'ère géologique du Pléistocène moyen et supérieur.
- On atteste que cette espèce est éteinte.
Références
- Zeitoun 2005, p. 2.
- Pautreau et Zeitoun 2005, p. 7.
- Monition 1981, p. 14.
- Bucher et Frey 2002, p. 175-176.
- Bucher et Frey 2002, p. 29.
- Monition 1981, p. 13.
- Monition 1981, p. 15.
- Meijer et Awe Due 2010, p. 707.
- Forestier et Paloute-Edoumba 2000, p. 15.
- Bird 2003, p. 4.
- Bird 2003, p. 6.
- Bird 2003, p. 11.
- Forestier et Paloute-Edoumba 2000, p. 14.
- Bird 2003, p. 5.
- Tortosa 2015, p. 301.
- Bird 2003, p. 16.
- Bird 2003, p. 12.
- Forestier et Paloute-Edoumba 2000, p. 16.
- Bird 2003, p. 17.
- Bird 2003, p. 3.
- Monition 1981, p. 17.
- Wong 2014, p. 26.
- Wong 2014, p. 24.
- Balzeau 2016, p. 30
- Monition 1981, p. 18.
- Monition 1981, p. 19.
- Tortosa 2015, p. 206-207.
- Van Welsen, Parnell et Ferry Slik 2011, p. 534.
- Van Welsen, Parnell et Ferry Slik 2011, p. 535.
- Van Welsen, Parnell et Ferry Slik 2011, p. 531.
- Van Welsen, Parnell et Ferry Slik 2011, p. 532.
- Beyer 1981, p. 43.
- Beyer 1981, p. 45.
- Van Welsen, Parnell et Ferry Slik 2011, p. 533.
- Beyer 1981, p. 44.
- Beyer 1981, p. 46.
- (en) G. Lehtinen, « Spiders of the Oriental-Australian region III. Tetrablemmidae, with a world revision », Acta Zoologica Fennica, vol. 162, , p. 1-151.
- (en) « Tetrablemma manggarai Lehtinen, 1981 — Taxonomic Serial No.: 868229 », sur ITIS report (consulté le ).
- « Otus alfredi », sur Système canadien d’information sur la biodiversité (SCIB) (consulté le ).
- (en) « Otus alfredi », sur avibase.bsc-eoc.org (consulté le ).
- (en) « Otus alfredi », sur IOC World Bird List (consulté le ).
- (en) « Otus alfredi », sur Tree of Life (consulté le ).
- Beyer 1981, p. 47.
- (en) « Occidozyga floresiana Mertens, 1927 », sur The American Museum of Natural History (consulté le ).
- (en) « Oreophryne rookmaakeri », sur amphibiaweb.org (consulté le ).
- Erna Chambon, « Les séismes en Indonésie », sur Académie de Versailles section géologie, (consulté le ).
- Grimaud-Hervé, Serre et Bahain 2015, p. 10.
- Grimaud-Hervé, Serre et Bahain 2015, p. 11.
- Tortosa 2015, p. 37.
- Lorenzo Pareto, « Note sur les subdivisions que l'on pourrait établir dans les terrains tertiaires de l'Apennin septentrional », Bulletin de la Société Géologique de France, 2e série, vol. 22, , p. 210-277 (lire en ligne [PDF]).
- Grimaud-Hervé, Serre et Bahain 2015, p. 23.
- Tortosa 2015, p. 249.
- Bucher et Frey 2002, p. 182.
- Bucher et Frey 2002, p. 180.
- Bucher et Frey 2002, p. 183.
- Bucher et Frey 2002, p. 184.
- Bucher et Frey 2002, p. 177.
- Bucher et Frey 2002, p. 178.
- Brown et Morwood 2004, p. 1058.
- Brown et Morwood 2004, p. 1059.
- Bucher et Frey 2002, p. 179.
- Brown et Morwood 2004, p. 1060.
- Bucher et Frey 2002, p. 181.
- Monition 1981, p. 16.
- Balzeau 2016, p. 35.
- Meijer et Awe Due 2010, p. 719.
- (en) G.D. Van Den Bergh, Rokhus Due Awe et al., « Insularity and its Effects : The youngest stegodon remains in Southeast Asia from the Late Pleistocene archaeological site Liang Bua, Flores, Indonesia », Quaternary international, vol. 182, , p. 18-46 (DOI 10.1016/j.quaint.2007.02.001, lire en ligne, consulté le ).
- Meijer et Awe Due 2010, p. 709.
- Zeitoun 2005, p. 3.
- Forestier et Paloute-Edoumba 2000, p. 19.
- (en) « Stegodon florensis Hooijer 1957 (elephant) », sur fossilworks.org (consulté le ).
- Balzeau 2016, p. 31
- Grimaud-Hervé, Serre et Bahain 2015, p. 76.
- Tortosa 2015, p. 86.
- Tortosa 2015, p. 87.
- Zeitoun 2005, p. 4.
- Tortosa 2015, p. 301
- Tortosa 2015, p. 228.
- Grimaud-Hervé, Serre et Bahain 2015, p. 75.
- Détroit 2012, p. 65.
- Balzeau 2016, p. 32.
- Détroit 2012, p. 61.
- Grimaud-Hervé, Serre et Bahain 2015, p. 94.
- Grimaud-Hervé, Serre et Bahain 2015, p. 95.
- Wood 2014, p. 31.
- Wood 2014, p. 34.
- Brown et Morwood 2004, p. 1063.
- Bruno Maureille, « L'ADN en Europe et en Asie : Un nombre d'espèces en constante évolution », La Recherche, Belin, no 491, , p. 36-39 (ISSN 0029-5671).
- Meijer et Awe Due 2010, p. 708.
- Meijer et Awe Due 2010, p. 710.
- Meijer et Awe Due 2010, p. 711.
- Meijer et Awe Due 2010, p. 712.
- Meijer et Awe Due 2010, p. 713.
- Grimaud-Hervé, Serre et Bahain 2015, p. 77.
- (en) Jelle S. Ziijstra, Lars W. van den Hoek Ostende et Rokus Awe Due, « Verhoeven's giant rat of Flores (Pagagomys theodoverhoeveni) : extinct after all ? », Contributions to Zoology, vol. 77, no 1, , p. 25-31 (lire en ligne, consulté le ).
- (en) « Armandville », dans Bo Beolens, Michael Watkins et Michael Grayson, The Eponym Dictionary of Mammals, Boston, The Johns Hopkins University Press, (ISBN 978-0-8018-9304-9, lire en ligne), p. 18.
- Tortosa 2015, p. 108.
- Grimaud-Hervé, Serre et Bahain 2015, p. 29.
- Henri Chambert-Loir, « Le 3e Congrès archéologique indonésien (Pertemuan Ilmiah Arkeologi III), Ciloto, 23-28 mai 1983 », Archipel, vol. 28, , p. 19-24 (DOI 10.3406/arch.1984.1911, lire en ligne, consulté le ).
- Kate Wong, « L’ancêtre de l’Homme de Florès retrouvé », sur Pourlascience.fr (consulté le )
- Brown et Morwood 2004, p. 1064
- Brown et Morwood 2004, p. 1065
- Balzeau 2012, p. 6.
- Balzeau 2012, p. 7.
- Tortosa 2015, p. 26.
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- Antoine Balzeau, « Homo sapiens : Le dernier homme », Les Dossiers d'archéologie, Faton, no 351, , p. 2-7 (ISSN 1141-7137)

- Antoine Balzeau, « Homme de Florès : Son crâne nous renseigne sur ses origines », Archéologia, Faton, no 351, , p. 30-35 (ISSN 0570-6270)
- Antoine Balzeau et Philippe Charlier, « Les origines d'Homo floresiensis d'après l'intérieur de son crâne : La morphologie interne du crâne d'Homo floresensis », sur Hominidés.com, (consulté le )
- (en) Peter Bird, « An updated digital model of plate boundaries », Geochemistry Geophysics Geosystems, vol. 4, no 3, , p. 1-52 (ISSN 1525-2027, DOI 10.1019/2001GC000252, lire en ligne, consulté le )
- (en) Herald Broch Beyer, « Cultural Variation on the Islands in the Sea of Flores. », Achipel, vol. 22, , p. 43-53 (DOI 10.3406/arch.1981.1668, lire en ligne, consulté le )
- (en) Peter Brown, Mike Morwood et al., « A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia », Nature, vol. 431, , p. 1055-1065 (lire en ligne, consulté le )
- (en) Kurt Bucher et Martin Frey, Petrogenesis of Metamorphic Rocks, Springer Science & Business Media, , 7e éd., 341 p. (ISBN 3-540-43130-6, lire en ligne)
- Florent Détroit, « Homo sapiens en Asie : grandes migrations... et rencontres inattendues », Les Dossiers d'archéologie, Faton, no 351, , p. 30-35 (ISSN 1141-7137)
- Hubert Forestier et Elise Paloute-Edoumba, « Les industries lithiques du Paléolithique tardif et du début de l'Holocène en Insulinde », Aséanie, vol. 6, no 1, , p. 13-56 (DOI 10.3406/asean.2000.1683, lire en ligne, consulté le )
- Dominique Grimaud-Hervé, Frédéric Serre, Jean-Jacques Bahain et al., Histoire d'ancêtres : La grande aventure de la Préhistoire, Paris IVe, Errance, coll. « Guides de la préhistoire mondiale », , 5e éd., 144 p. (ISBN 978-2-87772-590-3)
- (en) Hanneke J. M. Meijer et Rokus Awe Due, « A new species of giant marabou stork (Aves : Ciconiiformes) from the Pleistocene of Liang Bua, Flores (Indonesia) », Zoogical Journal of Linean Society, The Linean Society of London, vol. 160, no 4, , p. 707-718 (DOI 10.1111, lire en ligne, consulté le )
- Michel Balard (dir.), Jean-Pierre Mohin et Yvette Taborin, Les sociétés préhistoriques, Hachette Supérieur, coll. « HU Histoire / Histoire de l'Humanité », , 320 p. (ISBN 978-2-01-145984-8)
- L. Monition, « Florès et les « Petites îles de la Sonde » (The little Sunda islands) », Bulletin de l'Association de géographes français (58e année), nos 475-476, , p. 13-23 (DOI 10.3406/bagf.1981.5220, lire en ligne, consulté le )
- Jean-Pierre Pautreau et Valéry Zeitoun, « Présentation », Les Dossiers d'archéologie, Faton, no 302, , p. 6-15 (ISSN 1141-7137)
- (en) Peter C. Van Welsen, John A. N. Parnell et J. W. Ferry Slik, « Wallace's Line of plant distributions : Two or three phytogeographical areas and where to group Java ? », Biological Journal of Linean Society, no 103, , p. 531-545 (lire en ligne, consulté le )
- Kate Wong, « La saga humaine réécrite », Pour la science, Belin, no 445, , p. 24-27 (ISSN 0153-4092)
- Bernard Wood, « Le buissonnant rameau humain », Pour la science, Belin, no 445, , p. 28-35 (ISSN 0153-4092)
- Valéry Zeitoun, « Présentation : Coup de bambou sur la paléoanthropologie en Asie du Sud-Est », Les Dossiers d'archéologie, Faton, no 302, , p. 2-5 (ISSN 1141-7137)
Voir aussi
Articles connexes
- Homme de Florès
- Mata Menge, site préhistorique à 50 km de la Grotte de Liang Bua
Liens externes
- « Homo floresiensis - The Hobbit », sur hominides.com (consulté le ).
 Portail de la Préhistoire
Portail de la Préhistoire 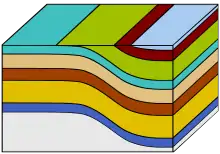 Portail de la géologie
Portail de la géologie  Portail de l’Indonésie
Portail de l’Indonésie