Fragmentation (écologie)
La notion de fragmentation ou de morcellement des écosystèmes / des habitats / écopaysagère(er) / écologique englobe tout phénomène artificiel de morcellement de l'espace, qui peut ou pourrait empêcher une ou plusieurs espèces de se déplacer comme elles le devraient et le pourraient en l'absence de facteur de fragmentation.
Avec le concept d'hétérogénéité, celui de fragmentation est une des bases théoriques de l'écologie du paysage.

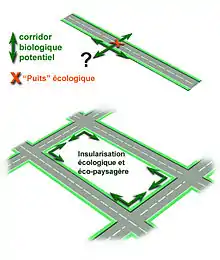
Remarque : les bords de routes semblent être de bons corridors, mais ils sont pollués par les gaz d’échappements et les matériaux catalytiques, et le risque d'écrasement et de prédation y est accru (manque d'abris)

La notion de fragmentation concerne uniquement le phénomène de morcellement, sans perte de surface. Toutefois, l’usage commun du terme contraint à en élargir la définition en y incluant la perte d’habitat. On parle alors de fragmentation per se quand on fait référence uniquement au morcellement, et de fragmentation quand on y inclut la notion de perte d’habitat[2]. En ce sens, un obstacle placé dans une matrice ne modifie ni le morcellement ni les habitats. Il modifie entre autres la connectivité écopaysagère mais n’est pas un élément strictement fragmentant.
Les individus, les populations et les espèces sont différemment affectés par la fragmentation de leur habitat. Ils y sont plus ou moins vulnérables selon leurs capacités adaptatives, leur degré de spécialisation, leur dépendance à certaines structures écopaysagères. D'autres facteurs sont leur capacité à voler ou à franchir les obstacles (parois, grillages, routes, zones traitées par des pesticides, etc.), et selon la biologie de leurs populations. Par exemple, les oiseaux forestiers, qui volent et qui peuvent donc exploiter des « taches » différentes, semblent beaucoup moins affectés par la diminution de la couverture forestière que par la fragmentation de la forêt elle-même [3] (hormis quelques exceptions notamment dues aux « effets-lisière » [4]).
Dans l'Évaluation des écosystèmes pour le millénaire, la fragmentation écologique est considérée comme une des premières causes d'atteinte à la biodiversité, avant la pollution[5]. L'Observatoire wallon de la biodiversité estime par exemple qu'en 2000, 5 à 15 % des espèces ont déjà disparu, que 30 à 50 % sont en régression et que « la disparition et la fragmentation des espaces naturels en est la première cause»[6]. Toutefois, la fragmentation per se, c’est-à-dire sans considérer la perte d’habitats, est un phénomène majoritairement positif[7], dont les avantages sont liés à la théorie du SLOSS.
État des lieux
Tous les pays riches en infrastructures routières, en canaux et, moindrement, ferroviaires sont concernés.
Les réseaux régionaux de transport ferroviaire et routier ne sont pas l'apanage des régions riches ou des régions abritant une capitale, dont en Europe[8], mais plutôt des régions densément peuplées et industrielles. En Europe, les réseaux autoroutiers les plus denses et parmi les plus fragmentants ont été construits aux Pays-Bas (plus de 100 km d'autoroute par 1 000 km2 dans l'ouest du pays), au Luxembourg et paradoxalement à Chypre. Des régions industrielles à PIB peu élevé (Nord-Pas-de-Calais) ou à PIB élevé, telles que celles de Düsseldorf et de Cologne (Allemagne) ou la Comunidad de Madrid (Espagne), ont un réseau autoroutier très dense. Certaines capitales (Berlin, Prague, Bucarest) ont encouragé le train plutôt que les autoroutes. Les grandes infrastructures portuaires ont encouragé le maillage autoroutier et ferroviaire.
En 2011, pour l'Agence européenne pour l'environnement (AEE)[9], la fragmentation est l'une des premières causes de régression de la biodiversité en Europe ; elle s'est fortement aggravée depuis le début des années 1990, avec de graves conséquences pour la faune et flore européennes. De plus, le nombre de projets d'infrastructures continue à augmenter ou est stable. Les pays les plus fragmentés sont la Belgique, le Luxembourg, les Pays-Bas, suivis de près par Malte, l'Allemagne et la France. Londres, le Nord de la France et la région parisienne le sont cependant à des niveaux comparables à ceux des pays les plus touchés. La densité d'autoroutes est plus élevée en République tchèque, en Belgique et en Allemagne. Le pays le mieux préservé de la fragmentation est la Roumanie, grâce notamment à 13 grands parcs nationaux et 500 zones protégées. Ce pays a conservé un habitat peu fragmenté qui abrite 60 % des ours, 40 % des loups et 35 % des lynx européens[9]. La Pologne, après une période de prise en compte des réseaux écologiques, s'est lancée dans un vaste programme routier et autoroutier qui devrait y augmenter fortement la fragmentation[9].
Dans plusieurs pays comme l'Espagne, la Grèce, l'Italie et la France, les zones centrales sont relativement préservées, mais les zones littorales sont de plus en plus fortement fragmentées. Les différences sont parfois contrastées dans un même pays, par exemple au Royaume-Uni où la région londonienne a un niveau record de fragmentation (malgré sa ceinture verte), mais où les Highlands écossais comptent parmi les zones où l'intégrité paysagère est la mieux préservée en Europe, grâce notamment à une faible densité de population, comme dans certaines régions scandinaves moins peuplées[9]. Les Pays-Bas ont depuis 1984 une politique ambitieuse de défragmentation du paysage qui a par exemple, comme en Belgique, permis un début de restauration des populations de blaireau grâce à de petits écoducs en tunnels qui évitent qu'ils se fassent écraser sur les routes[9].
L'AEE appelle à multiplier le nombre d'écoducs, y compris sur les routes anciennes pour permettre aux animaux de se déplacer. L'AEE recommande aussi de détruire des routes anciennes ou dont le trafic est en baisse plutôt que d'en construire de nouvelles, ce au profit du train et d'alternatives. Elle recommande également de planifier des contournements près des zones faunistiquement importantes au lieu de continuer à construire des routes et voies ferrées les unes à côté des autres[9].
Origine du concept

Les ouvrages de Forman et Godron et 1986[13] et Forman en 1995[14] ont formalisé les bases de l'analyse quantitative de la structure et des composantes du paysage (ici compris comme une surface variable composée d’une mosaïque d'écosystèmes[15]), suivi notamment de Opdam en 1991[16] Ces auteurs ont montré que la fragmentation d’un habitat naturel est une forme de destruction de l'habitat qui, à l'exception de quelques espèces ubiquistes, affecte d’abord la taille des populations en augmentant les risques d'extinction[17].
Wiens en 1997 fait valoir que la fragmentation modifie la taille, les formes et l’isolement des « taches » du paysage, générant un impact sur les fonctions écopaysagères des éléments ainsi isolés et - par effet second - sur des éléments situés aux niveaux supérieurs ou inférieurs du paysage. La fragmentation ;
- induit le remplacement d’éléments écopaysagers par d’autres,
- modifie le contexte spatial, notamment en altérant la connectivité écologique et donc le degré d'intégrité éco-paysagère.
- est un puissant facteur d'isolement écologique de milieux naturels (ou semi-naturels) au sein d’une « matrice » plus « hostile » (car aux conditions environnementales altérées par les activités humaines ou leurs conséquences secondaires).
- par l’insularisation qu’elle provoque, en diminuant les taux de dispersion et d’immigration augmente les risques d'extinction[18].
- modifie les effets-bordure (écotone/effet lisière), plus ou moins selon les caractéristiques de la matrice environnante[19].
En France le concept a notamment été diffusé par le réseau Man and Biosphère de l'UNESCO, et le travail de Françoise Burel (Thèse à l'Université de Rennes) a été le début d'une suite de thèses et de travaux à Rennes, Montpellier et ailleurs.
Enjeux


L'enjeu principal est celui de l'intégrité écologique du milieu, car la fragmentation des habitats s'oppose aux besoins vitaux qu'ont la faune, la fonge et la flore de pouvoir se déplacer dans le paysage. Elle contribue à altérer la qualité des habitats et à l’isolement des populations. Or, la probabilité de recolonisation dépend pour chaque espèce activement ou passivement mobile de 3 facteurs[21] :
- relations spatiales entre éléments paysagers utilisés par la population, dont les lieux de reproduction et les éléments de la mosaïque de tache à travers laquelle les disperseurs de gènes ou propagules doivent se déplacer (par exemple d'une mare à l'autre pour des espèces aquatiques[20]) ;
- caractéristiques de dispersion de l'organisme (qui selon l'espèce peut ou non nager, ramper, voler, courir ou se déplacer dans le sol, ou se faire transporter, etc. plus ou moins rapidement, pondre des œufs plus ou moins durablement résistants à la dessication[22], etc.) ;
- les changements temporels dans la structure du paysage.
Les données de terrain, comme les analyses comparatives issues de la cartographie des corridors biologiques, d’images satellitaires, de photographies aériennes, ou de cartes anciennes montrent que les écosystèmes sont de plus en plus artificialisés et fragmentés, par des infrastructures de transport de véhicules, biens ou personnes ou d'énergie (lignes électriques, lignes téléphoniques). Les barrages hydroélectriques en sont un équivalent visible et facile à comprendre pour les réseaux fluviaux, mais d'autres barrières écologiques non physiques existent (pollutions par des pesticides, des eutrophisants ou d'autres biocides sont également des facteurs majeurs de fragmentation, plus discrets, mais touchant de vastes espaces.
Les individus, mais aussi les populations sont confrontés à un nombre croissant de « verrous écologiques » (barrières physiques ou immatérielles) ou « goulets d'étranglement » ou pièges écologiques, qui — en fragmentant anormalement les écosystèmes — limitent ou interdisent la circulation des individus et populations et donc le mélange des gènes au sein de leur aire normale de répartition, au point de provoquer des régressions ou disparitions d'espèces, ou de les menacer à moyen ou long terme.
Dans deux cas, la fragmentation anthropique semble avoir un aspect localement et provisoirement positifs ; C'est le cas pour certaines espèces écotoniales, souvent ubiquistes, qui voient ainsi la longueur de leur habitat augmenter (mais souvent de manière linéaire, ce qui les rend plus vulnérables à leurs prédateurs ; c'est l'effet-lisière). Un autre bénéfice, très relatif car généralement provisoire, est que la fragmentation d'un milieu peut limiter la diffusion de certaines espèces invasives (par ex Bothriochloa ischaemum, une herbacée invasive, s'est répandu moins rapidement au Texas sur les parcelles fragmentées que non fragmentées, sachant qu'inversement, les bords d'infrastructures fragmentantes de transport, sont de bons corridors de dispersion pour de nombreuses invasives[23].
Même des animaux volants sont affectés : On a longtemps cru que les oiseaux et chauves-souris échappaient aux impacts de la fragmentation par les routes forestières ou des cœurs de forêt[24], mais outre que certaines espèces sont très affectées par le Roadkill, diverses expériences (dont en Guyane dans le cadre de l'étude d'impact et des mesures compensatoires de la RN2) ou au Brésil[25] notamment sur un site expérimental situé à 80 km au nord de Manaus ont montré que même sur des routes peu fréquentées, les oiseaux et les chauve-souris sont affectés. La fragmentation forestière et des zones humides ou antérieurement sauvages est également responsable du déclin des chiroptères[26] — en dépit de leur capacité à voler — sont affectés par la présence de routes dans les forêts. Sur les voies fréquentées et bruyantes, le seul bruit éloigne les oiseaux chanteurs (jusqu'à plus d'un km sous le vent dominant d'autoroutes en Grande-Bretagne) et les oiseaux urbains doivent adapter leurs chants pour s'entendre, notamment dans les basses fréquences[27]
L'écologie du paysage tente de répondre à des questions cruciales pour la Conservation de la nature ; En particulier, quelle taille minimale d'habitat et quelle qualité d'interconnectivité faut-il conserver pour assurer la survie des espèces qui y coévoluent ? Protège-t-on mieux un plus grand nombre d'espèces et d'écosystèmes dans une grande réserve naturelle ou dans plusieurs petites ?
Définition


Des barrières naturelles existent dans la Nature. Les plus importantes sont les longues et hautes chaines montagneuses (p. ex. : Alpes, Pyrénées, Andes, Himalaya). Les grands déserts, les bras de mer en constituent d’autres (pour les espèces continentales). Les océans et la plupart des mers sont physiquement interconnectés, mais les zones dépourvues de nutriment, pauvres en plancton ou à température, salinité ou profondeur particulières présentent des caractéristiques qui leur confèrent une fonction de barrière pour certaines espèces (coralliennes par exemple).
Ces barrières peuvent être traversées en les contournant ou, dans le cas des montagnes, par les cols et/ou par certaines espèces (oiseaux notamment). Les oiseaux migrateurs étant eux-mêmes porteurs de divers parasites ou espèces involontairement transportées, parfois sous forme d’œuf ou de spore, ils contribuent deux fois par an à transporter divers organismes, qui ne constituent cependant qu’une infime part de la biodiversité.
Pour les espèces continentales, dans un bassin versant normal (illustration), ce sont les fleuves qui, s’ils sont des corridors biologiques pour certaines espèces, sont les principales barrières naturelles. Les rivières sont des obstacles moins importants et les ruisseaux encore moindres. La plupart des espèces, après un certain temps peuvent traverser ou contourner les fleuves. Dans la nature, le bassin versant est entièrement potentiellement accessible à de nombreuses espèces. Pour les espèces qui traversent difficilement un canal ou une route, ou sont incapables de traverser un champ traité par des pesticides, le paysage anthropisé, tel qu’il se développe depuis quelques décennies devient une succession d’obstacles plus ou moins infranchissables[29].
L’Homme prend peu à peu conscience qu’il est à l’origine de nombreuses et nouvelles barrières écologiques qui sont soit des zones impropres à la vie de nombreuses espèces, soit des zones simplement défavorables à la circulation et/ou à la reproduction des espèces. Elles peuvent être matérielles et visibles (un mur, une ligne de TGV ou une autoroute en France munis d’une double-clôture), mais elles sont souvent invisibles à nos yeux (on les dit alors parfois « immatérielles »). Il est possible que tous les types de barrière écologique ne soient pas encore identifiés.
- Exemples de barrières matérielles :
- infrastructures construites, telles que éoliennes, routes, autoroutes, voies ferrées, TGV, canaux (le canal de Panama ou de Suez coupent et isolent des continents), fleuves canalisés, barrages-réservoir ou hydroélectriques, lignes à haute tension, antennes ou câbles sur les corridors de migration, etc.
- zones urbanisées et très dévégétalisées
- zones artificialisées
- zones d’agriculture et de sylviculture intensive
- zones d’expérimentation ou culture OGM; Outre que certaines plantes produisent leurs propres pesticides, ces champs doivent théoriquement être gérés pour ne pas faciliter la dispersion de pollens, graines ou autres propagules.
- Exemple de barrières immatérielles :
- odeurs (l’odeur laissée par un chien ou par un humain peut durant quelques heures ou jours perturber des animaux craintifs qui y sont très sensibles)
- microclimats (température, hygrométrie, …)
- dérangement, perturbation, surfréquentation
- pollution sonore
- pollution lumineuse
- pollution thermique de l’eau, voire de l'air ou du sol (réseau de chaleur mal isolé)
- pollution atmosphérique (biocide, ozone, métaux lourds, …)
- pollution électromagnétique (lignes haute-tension, antennes-relais…)
Effets de seuils
La relation entre la fragmentation et la régression de la biodiversité est de type « non linéaire »[30].
À titre d'exemple, des scientifiques ont recherché d'éventuels « seuils écologiques » dans les boisements fragmentés d'une zone périurbaine de Lombardie (Italie du Nord)(étude publiée en 2010).
Ils ont pour cela choisi une batterie d'indicateurs au sein de la strate herbacée. Ces indicateurs étaient des plantes vivaces, typiques de trois syntaxons phytosociologiques (Fagetalia sylvaticae, Carpinion betuli et Erythronio-Carpinion), et de trois caractéristiques de fragmentation des boisements (localisation, degré d'insularité, forme du fragment). Les « facteurs écologiques » ont été évalués au moyen des indices de Landolt L et H ; calculés à partir de relevés antérieurs faits dans des forêts de référence et jugées de haute qualité écologique.
Certaines plantes (vivaces herbacées) se montrent en effet indicatrices de forêts anciennes, et d’autres de forêts hautement fragmentées[30]. Sur des bases statistiques, des « modèles linéaires généralisés » ont été construits à partir des relevés faits en forêt ; Ils montrent une corrélation entre le nombre d'indicateurs forestiers et la taille des fragments, ainsi que leur degré de connectivité avec d’autres milieux forestiers[30]. Les modèles additifs généralisés confirment une relation non linéaire entre l’indicateur de richesse et la taille des fragments ; avec présence de deux seuils[30] :
- ) Le nombre d'indicateurs forestiers a augmenté jusqu'à 35-40 ha (au-dessus de ce seuil de superficie, la richesse herbacée est restée constante).
Dans cette région d’Italie, les politiques de conservation pourraient ou devraient donc viser à protéger les fragments de forêt atteignant au moins 35 à 40 ha d’un seul tenant ; - ) L’autre seuil concerne la relation entre l'aire et le nombre d'indicateurs ; cette relation est moins prononcée dans les fragments de moins de 1 à 1,5 ha
Donc, si les fragments de taille comprise entre ces 2 seuils sont agrandis, leur valeur pour la biodiversité végétale grandira fortement.
L’existence de ces 2 seuils suggère une réponse brutale à de la flore à certains changements écopaysagers[30]. Elle fournit aussi des objectifs quantitatifs en matière de conservation[30].Si l’on veut y conserver (ou restaurer) leur biodiversité, Il apparait utile - avant d’atteindre des seuils critiques - de défragmenter les forêts périurbaines. Les herbacées vivaces de forêts anciennes ou typiquement associées aux forêts hyperfragmentées semblent être des indicateurs fiables pour l’évaluation environnementale, mais ils varient selon chaque zone biogéographique.
Effets de bordure

Saunders et ses collègues en 1991, puis Collinge en 1996 montrent que les bordures artificiellement créées par la fragmentation subissent des altérations immédiates de facteurs importants tels que température, hygrométrie, vent, luminosité/albédo, odeurs qui perturbent ou bloquent les déplacements de certaines espèces. D'autres facteurs sont souvent altérés à moyen et plus long terme (qualité de sol et de l'eau, niveau de la nappe, qualité de l'air, bruit/dérangement, etc.). Ces effets-lisière perturbent d'abord les communautés animales, fongiques et végétales qui se trouvent à proximité de la lisière des îles ou îlots isolés par le phénomène de fragmentation. Ranney et ses collègues en 1981, et Chen et son équipe en 1992 ou Matlack en 1993 démontrent ce fait, notant par exemple, que des "effets lisières" sont nettement mesurables dans les forêts nord-américaines fragmentées sur 15 à 240 mètres selon les espèces animales et végétales étudiées.
Le drainage et les pompages effectués dans la « matrice » écopaysagère peuvent – à la suite de la baisse de la nappe – bouleverser l'écologie d'un îlot qui semblait a priori épargné, et ce bien au-delà de ses lisières, jusqu'en son cœur et en profondeur[31].
Voir aussi l'article Lisière
Distance entre fragments

La distance entre fragments est un élément important de la théorie modèle de l'insularisation. Elle constituait un des indices de fragmentation[32], pratique pour la cartographie SIG. Néanmoins, dès 1986, Forman & Godron admettent que dans la réalité, l'effet de la distance entre fragments est relatif. Il varie selon les espèces (ubiquistes, volantes, nageant… ou spécialisées et inaptes aux grands déplacements) [13]. Et il est plus ou moins réduit ou à surpondérer selon d'autres facteurs, tels que :
- la nature de la matrice (le milieu qui sépare les îlots est plus ou moins « hostile » aux espèces insularisées) ;
- les habitats qu'elle abrite (habitats de substitutions parfois) ;
- le degré de similarité entre les îlots et les habitats de la matrice (proches ou au contraire très « opposés ») ;
- l'intensité, les dates, et les types d'activités humaines qui s'y déroulent, etc.
Effets du type de structure de réseau
Dans le cas des réseaux artificialisants et fragmentants de canaux, cours d'eau, voies ferrées, routes, etc., le réseau aura un impact très différent (du point de vue du morcellement) selon la longueur totale des infrastructures linéaires, mais plus encore selon le degré de fermeture (interconnexions) du réseau. Les deux extrêmes du point de vue de l'impact par la fragmentation sont :
- un réseau uniquement arborescent dont les branches ne se touchent pas (de nombreuses routes allant des communes littorales vers le trait de côte étaient autrefois ainsi structurées)
- un réseau maillé
C'est le nombre d'intersections (« carrefours ») et non de « nœuds » qui est à prendre en compte pour évaluer le caractère fragmentant d'un réseau.
Taille, superficie et forme des fragments
La taille, la superficie et la forme des fragments influent sur l'intensité des effets de bordure, pour des raisons géométriques notamment, et en fonction de leur exposition au vent, soleil, pluies, etc. Les fragments petits ou étroits seront les plus affectés par les effets de bordure, car ils ne disposent plus en leur centre d'importantes superficies intactes (parfois dites "cœur d'habitat" [13],[33],[14]).
Exemple : Morcellement par les réseaux de transport

Les infrastructures les plus opaques aux déplacements animaux et végétaux ou à haut risque de roadkill (mortalité animales par collision avec véhicules) restent le réseau de transport routier, autoroutier et ferroviaire (ligne TGV). Le réseau routier et autoroutier mondial continue à croître rapidement. Son maillage se densifie, s'interconnecte et les routes sont de plus en plus larges et « artificialisantes ». Le nombre de passages à faune est de très loin inférieur aux besoins. L’impact morcelant des routes a été très sous-estimé, mais on ne s’en est aperçu que très récemment. Faute d’être capable de le quantifier, les études d’impact continuent à le minimiser. Pour les petits mammifères, les insectes, les micro-organismes et certaines plantes, le problème est généralement purement et simplement éludé.
Pollution sonore et pollution lumineuse

- La pollution sonore semble se stabiliser dans les pays riches, mais croît rapidement dans les pays en développement. Contribuant au dérangement des espèces, elle concerne au moins 70 % des espaces verts urbains et jardins particuliers, et espaces naturels bordant les principaux axes de déplacements. Pour des raisons de coût, elle n’est considérée que pour les habitations très proches des zones urbanisées. Des études réalisées en Grande-Bretagne le long des autoroutes ont mis en évidence une disparition progressive des oiseaux chanteurs sur une bande de 2 km environ (et jusqu’à 4 kilomètres sous le vent dominant). Cette disparition est corrélée avec le niveau de décibels, mais varie aussi selon la proximité de l’axe, le relief, le trafic, le vent dominant, la nature du revêtement, la quantité de camions, la nature et la densité de la végétation, etc. À cet effet s’ajoute ceux de la pollution déposée par les véhicules (benzène, plomb, métaux du groupe du platine issus des pots catalytiques, etc).
Il serait intéressant de mesurer si le dérangement lié au bruit est identique sur les voies ferrées où le bruit est souvent plus important en décibels, mais discontinu dans le temps.
Aussi, le trafic aérien augmente de manière continue (de 7 à 10 % par an) dans les dernières années, et que son impact est très mal mesuré.
- La pollution lumineuse par l’éclairage des bâtiments, monuments et infrastructures (dont canaux et cours d'eau qui sont des axes de migration animale) et par les phares des véhicules n’est pas ou très peu analysée. Elle est très importante dans les pays riches. Il semble qu’elle ait des impacts qui ont été également très sous estimés, y compris pour les cours d'eau et la trame bleue qu'ils devraient constituer. Les grandes ou petites migrations de poissons et l'activité planctonique peuvent être perturbée par l'éclairage. Une grande partie des migrations animales se fait en effet de nuit, et la lumière naturelle et/ou artificielle influe fortement sur de nombreux processus du vivant. Elle perturbe les rythmes biologiques et notamment le rythme nycthéméral dont dépendent les modifications saisonnières de métabolisme et de comportement. Beaucoup d'espèces sont également sensibles aux cycles lunaires et à la luminosité de la lune (De nuit, la lumière du soleil (alors invisible) reflétée par la lune peut par exemple inhiber ou au contraire exciter l'activité de certains animaux aquatiques ; en particulier, le rythme nycthéméral influe sur les migrations quotidiennes (déplacement horizontaux et/ou verticaux) et l'activité de nombreuses espèces planctoniques dont les daphnies et autres invertébrés aquatiques et organismes zooplanctoniques). L'éclairage artificiel les perturbe, comme il perturbe des escargots aquatiques[34] ou des poissons migration, reproduction, alimentation)[35]. La lumière peut être source de situation de piège écologique pour les espèces qu'elle attire.
Le morcellement par les routes

Le morcellement par les routes s'exerce de manière plus visible en surface via un effet barrière, le roadkill et la pollution routière, le sel de déneigement ou l'éclairage facteur de pollution lumineuse, mais un effet-barrière existe aussi pour les espèces du sol (ou fouisseuses) et pour d'autres espèces qui utilisent leurs galeries. Les espèces dont les spores, les germes ou les graines, les organismes ou les propagules sont transportées par les petits mammifères ou des animaux du sol (zoochorie) peuvent aussi être affectées. Les fonds de route auxquels on a souvent ajouté de la chaux et/ou du ciment sont homogénéisés et fortement damés, au point de présenter des résistances égales à celle d’un béton. Ils constituent une barrière physique absolue pour les taupes, micro-mammifères, vers de terre, insectes et leurs cortèges de micro-organismes associés. Ces animaux fouisseurs ne peuvent plus exercer leur fonction de « tunneliers » décolmatant, drainant et aérant les sol, qui participe à l’entretien des continuums biologiques souterrains dont on découvre ou redécouvre l’importance pour les arbres notamment. On trouvera dans la semelle routière quelques bactéries, mais dont la mobilité est très réduite. Si de la matière organique a été enfouie, seuls les champignons et quelques micro-organismes circuleront et éventuellement perceront le macadam. Parfois, les racines des arbres perceront le substrat là où il est fragile ou faillé, mais sans compenser l’absence d’un continuum superficiel. Les tunnels offrent un moyen de circuler en préservant des continuités écologiques, mais ils sont coûteux et depuis l’accident du tunnel du Mont-Blanc, les aménageurs les limitent. Des espèces parfaitement volantes peuvent aussi être affectées. Par exemple, une étude récente a montré chez deux espèces de chauve-souris menacées, que le Myotis bechsteinii (spécialiste de la chasse près du sol) et qu'une espèce telle que Barbastella barbastellus (qui chasse plus en hauteur) sont différemment impactées par une autoroute : La première ne traverse que peu l’infrastructure, et seulement en empruntant les souterrains. Elle chasse et se reproduit moins bien aux abords de l'autoroute. Alors que l'autre espèce (B. barbastellus) y est bien moins sensible. En particulier, elle traverse plus volontiers l'autoroute, au-dessus ou en dessous. Ces deux espèces nécessitent des corridors, des mesures compensatoires et conservatoires différentes[36]. Pour certaines espèces, la route se comporte comme un « piège écologique », en les attirant sur un milieu mortellement dangereux pour elles. Ces phénomènes sont mieux étudiés [37] et mieux compensés[38], mais les routes restent de puissants facteurs de morcellement des habitats.
Les barrières chimiques


Les barrières chimiques. Les fonds de route sont en effet parfois constitués de crassiers industriels, de cendres toxiques d’incinérateurs, de mâchefers riches en métaux lourds et autres micro-polluants, de certains déchets de fonderie. Des millions de tonnes de ces produits ont été acceptés avec des teneurs en métaux lourds et/ou en polluants organiques bien supérieures aux normes sol (normes AFNOR). De même, parfois moins officiellement, d’autres déchets dits inertes ou stabilisés et dont on s’aperçoit à l’usage qu’ils ne l’étaient pas, ont servi par exemple à restaurer des berges de rivières ou de canaux. Dans les milieux argileux acides et/ou sableux (souvent naturellement acides) les chantiers sont stabilisés à la chaux et/ou au ciment (jusqu’à plus de 10 kg/m2). Ces produits sont « toxiques » pour un grand nombre d’espèces qui vivent dans les milieux acides. Les laitances et les produits de ruissellement des bétons spéciaux frais, tels que ceux qui servent aux ouvrages de type dalles, fossés, bassins, muret coulé sur place, atteignent un pH de 11. Bien que les adjuvants de fabrication, au dire de leurs utilisateurs soient labellisés NF, ces laitances peuvent présenter un caractère neurotoxique spectaculaire (paralysie et mort immédiate par simple contact) pour la plupart des animaux à sang froid… et ce pendant plusieurs mois.
Des bas côtés, dépendance ou champs proches traités avec des pesticides constituent une barrière supplémentaire et absolue pour de nombreuses espèces, souvent durant la période de culture qui est aussi la période de développement des espèces qui auraient besoin de circuler sur le territoire. Les remembrements et l'intensification agricole qui suivent souvent les constructions de routes ont produit dans presque tous les pays des champs de plus en plus vastes et hostiles à la biodiversité. Les routes ont aussi contribué au recul du bocage et des réseaux de fossés et de chemins qui constituaient autrefois un immense réseau permettant aux espèces de circuler. La densification des transports, et leur moindre coût relatif ont permis l'élevage hors sol, qui s'est traduit par un recul de l’herbage au profit du maïs, ce qui a également nui aux espèces des milieux ouverts et enherbés, et à leurs possibilités de mobilité.
Qualité des sols

Sécheresse et qualité du sol. Le drainage et l’imperméabilisation des routes et leurs abords sont de plus en plus efficaces. L’eau de ruissellement (polluée) est collectée dans des fossés parfois totalement imperméables, vers des bassins eux aussi souvent imperméables et/ou isolés (grillages de sécurité, parois abruptes et/ou plastifiées). Bien entendu, les risques réels de pollution de la nappe phréatique, particulièrement dans les champs-captant justifient la séparation des eaux. Mais l’absence d’eau ou d’humidité dans les fossés et sous les routes est un facteur supplémentaire de disparition d’espèces. Un sous-sol trop sec est par ailleurs inhospitalier et impénétrable par la plupart des espèces fouisseuses. Il s’oppose à la circulation souterraine de la faune. L’effet drainant des routes encastrées s’ajoute à celui de l’agriculture, et la semelle de labour qui s’étend dans les campagnes bloque les espèces fouisseuses dans un volume qui est tout entier exposé au soc de la charrue et aux chocs thermo-hygrométriques.
Le continuum thermo-hygrométrique



Rupture du continuum thermo-hygrométrique Les mesures conjointes de température et d’humidité réalisées en bordure de route ou de zones imperméabilisées montrent que l’effet de tranchée (une route dans une forêt par ex.) et les effets de lisières peuvent se traduire par des chutes importantes et permanentes de l’hygrométrie avec des impacts qui avaient été très sous-estimés :
- Disparition totale et durable de la rosée sur une bande qui peut atteindre 40 m sur le bord de certaines infrastructures exposées (zones de plateaux à nappes perchées, falaises, …)
- Abaissement du plafond de la nappe
- Diverses perturbations hydrologiques et pédologiques peuvent accompagner et renforcer le phénomène.
- Effet desséchant. Les routes larges et linéaires au macadam noir, comme les tranchées forestières, où la dévégétalisation d’un sol exacerbent parfois très fortement l’effet déshydratant du vent et du soleil. Ils entraînent une chute de l’hygrométrie très importante (2 à 3 fois moins d’humidité) avec des chiffres qui peuvent parfois être inférieurs aux moyennes sahariennes, y compris en zone tropicale humide en début de saison des pluies (Guyane française). Ce phénomène a même paradoxalement été observé au-dessus d’une étendue d’eau (barrage EDF-Petit Saut en Guyane, là où sur une grande superficie, les arbres ont été noyés et n’évapotranspirent plus). Nombre d’insectes et certains micro-mammifères manifestent de très nets comportements d’évitement des zones découvertes et dévégétalisées où l’hygrométrie chute, où la température varie brutalement. Pour certaines espèces qui sont pourtant rapides et vives à la course ou au vol, ceci se vérifie de jour comme de nuit et même sur une route fermée à la circulation depuis plusieurs mois. Pour ces espèces, une route semble être l’équivalent d’un mur vertical invisible et infranchissable, dont « l’épaisseur » peut être bien supérieure à la route… Pour ces mêmes espèces, un chemin de terre végétalisée, une route pavée ou un parking semi-végétalisé où l’herbe pousse n’est pas un obstacle. On a montré que même pour une route forestière de 10 m de large où ne passent que 25 voitures/jour, le phénomène existe. L’impact sur les oiseaux est plus complexe et différent selon les espèces. Ce sont les mammifères domestiques (chiens, chats), et les mammifères sauvages (mustélidés, renard, genette, lapin de garenne, chevreuil, sanglier, hérisson, rats, souris, musaraigne) qui semblent les moins gênés (et aussi les amphibiens au moment des migrations) mais ils sont des victimes potentielles des véhicules…
Pour toutes ces raisons le réseau routier interdit la circulation transversale et longitudinale de nombreuses espèces. Ce faisant, il perturbe fortement les diffusions et les flux naturels de gènes indispensables au fonctionnement normal des écosystèmes. La fragmentation des écosystèmes est également responsable de la perte de diversité génétique : les espèces, confinées dans des habitats de plus en plus restreints sont condamnées à l’endogamie, ce qui provoque des problèmes de consanguinité et de dérive génétique.
Les réseaux d’infrastructures et l’urbanisation qu’ils encouragent continuent en outre à consommer de l’espace naturel agricole (l’équivalent surface de deux départements serait déjà imperméabilisé et donc transformé en quasi-désert biologique ou espace écologiquement très appauvris en France ; l’imperméabilisation qui présente quelques avantages pratiques à court terme, se paye en sécheresse, en inondation et en érosion, en pollution et en nuisances de toutes sortes).
Quelques rares espèces de micro-mammifères ou d’oiseaux (pies, corneilles, étourneaux, pigeons, faucons crécerelles) ont trouvé le long des routes des milieux de substitution. Le réseau routier est néanmoins de plus en plus hostile et « imperméable » à la circulation de la faune et de la flore (les fossés de béton, clôtures à petites mailles, murs anti-bruits, bordures, parapets, terre-pleins ou murets centraux étanches à la petite faune, drainage de la route et des berges, tranchées de plus en plus larges, fauchage fréquent, utilisation de pesticides et d’inhibiteurs de croissance, cumulent leurs effets respectifs.)
Certaines espèces sont reconnues comme particulièrement utiles ou importantes pour le fonctionnement des écosystèmes et à ce titre protégées par la loi. Parmi elles, le hérisson et les amphibiens, dont respectivement plus d’un million et plusieurs millions d’individus sont encore chaque année victimes de la circulation, bien que leur nombre ne cesse de se réduire. De la même manière, quelques espèces végétales ont été favorisées par les infrastructures, mais ce sont souvent des plantes banales, ubiquistes voire invasives (renouée du Japon, Balsamine, etc). Les plantations très homogènes (y compris génétiquement) des bords d’infrastructures ont aussi favorisé la diffusion de microbes ou parasites (ex : feu bactérien pour les rosacées qui étaient autrefois plantées sur les bords d’autoroutes).
En Afrique, et dans de nombreux pays en développement des bandes parallèles aux routes étaient utilisées le jour par les marcheurs, les ânes et les chevaux et autres animaux de bât qui longeaient les routes, permettant de nuit un usage comme corridor biologique par certaines espèces sauvages. Souvent, notamment en raison d'accidents nombreux, les autorités tendent à élargir les routes et à supprimer ces accotements végétalisés.
Cartographie
Divers types de cartographies, souvent sur SIG décrivent la fragmentation écologique, notamment pour la cartographie des corridors biologiques.
En France, dans le cadre des accords de Grenelle et de la Trame verte et bleue, le CEMAGREF a produit, sur la base des données européennes (UE – SOeS ; CORINE Land Cover 2006), de l'IGN 2006, et de l'IFN 2010, une carte de La fragmentation des espaces naturels par région forestière départementale[39].
Solutions provisoires
Pour compter les amphibiens et les reptiles qui traversent la route et/ou pour les guider vers un batrachoduc (tunnel leur permettant de traverser sous la route, plus en sécurité), des dispositifs provisoires collecteurs (de type panneaux, bâches ou filets à mailles très fines) peuvent être disposés sur les bords des routes à proximité des points connus de traversée au moment des migrations des amphibiens (généralement des adultes se rendant à leur zone de ponte au printemps, ou en revenant). Ces dispositifs ne sont posés qu'au moment des migrations et les animaux collectés dans des seaux sont transportés de l'autre côté, ou ils sont permanents s'il existe un batrachoduc. Les tritons sachant escalader une paroi verticale lisse, un rebord est souvent prévu sur la partie haute. Des écoducs peuvent ensuite être positionnés aux points judicieux, qu'on peut aussi avoir repéré en comptant les cadavres d'animaux.
Ces dispositifs, s'ils ne sont pas aussi correctement enterrés sont parfois moins efficaces pour les jeunes de l'année qui retournent dans les milieux boisés ou prairiaux, ne mesurant que quelques millimètres, ils passent parfois sous les plaques rigides censées les guider et meurent alors nombreux, déshydratés ou écrasés sur la route.
Une autre possibilité est de capturer des animaux et de les transporter (par camion en général) de l'autre côté d'une infrastructure très fragmentante (ex : autoroute ou TGV clôturé). Cela limite le risque de dérive génétique, mais c'est un facteur important de stress pour les animaux.
 Dispositif guidant les animaux vers un point de traversée (Battrachoduc)
Dispositif guidant les animaux vers un point de traversée (Battrachoduc) Panneau alertant les automobilistes
Panneau alertant les automobilistes
Législation
En France, les accords de Grenelle instituent une Trame verte et bleue, qui a notamment pour objectif [40] « la remise en bon état des milieux nécessaires aux continuités écologiques », avec en particulier « le rétablissement ou l'amélioration de leur fonctionnalité. Elle s'effectue notamment par des actions de gestion, d'aménagement ou d'effacement des éléments de fragmentation qui perturbent significativement leur fonctionnalité et constituent ainsi des obstacles »[40]. Toutefois, ces actions doivent aussi tenir « compte du fonctionnement global de la biodiversité et des activités humaines »[40].
Voir aussi
Articles connexes
- Biodiversité
- Écologie du paysage
- Trame verte
- Route HQE
- quinzième cible HQE
- écoduc
- Intégrité écologique
- Connectivité écologique
- Corridor biologique
- Mortalité animale due aux véhicules
- Pollution lumineuse
- Migration animale
- Liste des animaux migrateurs
- Fragmentation forestière
- Cloisonnement forestier
- Biologie de la conservation
- Théories de la dispersion des individus
- Lombriduc
- Complémentarité des habitats
Liens externes
- (fr) Document IFEN (Oct. 2006) sur certains impacts des routes et véhicules
- (fr) Page du Site PDBFF du projet consacré à l'étude de la fragmentation de la forêt amazonienne brésilienne
- (fr) Note pédagogique du Ministère de l'Environnement Wallon sur la fragmentation écologique
- (fr) Thèse de doctorat intitulée "Mode d'utilisation du milieu fragmenté par une espèce forestière aux habitudes discrètes, la martre des pins Martes martes", par Vincent Pereboom (75 pages)
- La continuité écologique des cours d'eau Agence de l'eau Artois-Picardie
- Ministère de l'environnement La fragmentation des cours d'eau par des obstacles matériels, sur le système CARMEN (mis à jour en 2013) (seuils, barrages, digues, épis en rivière, grille de pisciculture. Les "freins à l'écoulement" ne sont pas hiérarchisés, un vannage non fermé depuis 20 ans est représenté comme une écluse importante ; il faut en outre croiser une telle carte avec les indices de pollution chimique, thermique, lumineuse et peut-être sonore). Une base de données complémentaires est prévue pour aller dans ce sens.
Bibliographie
- (fr) Morcellement du paysage en Suisse « Analyse du morcellement 1885–2002 et implications pour la planification du trafic et l’aménagement du territoire » (Version succincte, Office fédéral de la statistique)
- Léonard A. & Zegel P., 2010. Référentiel des obstacles à l'écoulement version 1 – Descriptif de contenu. ONEMA, 30p
- (en) Amezaga, J.M., Santamaría, L. & Green, A.J. (2002) Biotic wetland connectivity — supporting a new approach for wetland management policy. Acta Oecologica, 23, 213–222.
- (en) Didham R.K, Kapos V & Ewers R.M (2012) Rethinking the conceptual foundations of habitat fragmentation research. Oikos 121, 161–170
- (en) Fischer, J. & Lindenmayer, D. B. 2007. Landscape modification and habitat fragmentation: A synthesis. Global Ecology & Biogeography 16, 265-280 (Résumé).
- (en) Ledevin R & Millien V (2013),Congruent morphological and genetic differentiation as a signature of range expansion in a fragmented landscape ; Ecology and Evolution 3 (10.1002/ece3.2013.3.issue-12), 4172-4182
- (en) Lindenmayer, D. B. & Fischer, J. 2007. Tackling the habitat fragmentation panchreston (Panchreston signifie en anglais "qui explique tout"). Trends in Ecology & Evolution 22, 127-132 (Résumé)
- (en) Lindenmayer, D. B. & Fischer, J. 2006. Habitat fragmentation and landscape change: an ecological and conservation synthesis. Island Press, Washington D.C. (Habitat fragmentation and landscape change: an ecological and conservation Extraits avec Google livre)
- Brown, M. & Dinsmore, J.J. (1986) Implications of marsh size and isolation for marsh bird management. Journal of Wildlife Management, 50, 392–397
- Figuerola, J., Green, A.J. & Michot, T.C. (2005) Invertebrate eggs can fly: evidence of waterfowl mediated gene-flow in aquatic invertebrates. American Naturalist, 165, 274–280.
- Figuerola, J., Green, A.J. & Santamaría, L. (2003) Passive internal transport of aquatic organisms by waterfowl in Doñana, southwest Spain. Global Ecology and Biogeography, 12, 427–436.
- Trakhtenbrot, A., Nathan, R., Perry, G. & Richardson, D.M. (2005) The importance of long-distance dispersal in biodiversity conservation. Diversity and Distributions, 11, 173–181.
Références
- R.T.T.Forman, « Estimate of the area affected ecologically by the road system in the United States », Conservation Biology, vol. 14, , p. 31-35 (DOI 10.1046/j.1523-1739.2000.99299.x, lire en ligne [PDF])
- (en) Lenore Fahrig, « Effects of Habitat Fragmentation on Biodiversity », Annual Review of Ecology, Evolution, and Systematics, vol. 34, no 1, , p. 487–515 (ISSN 1543-592X et 1545-2069, DOI 10.1146/annurev.ecolsys.34.011802.132419, lire en ligne, consulté le )
- M. Kurtis Trzcinski, Lenore Fahrig, Gray Merriam, « Independent effects of forest cover and fragmentation on the distribution of forest breeding birds », Ecological Applications, vol. 9, no 2, , p. 586-593 (DOI 10.1890/1051-0761(1999)009[0586:IEOFCA]2.0.CO;2, lire en ligne [PDF])
- Cristina Banks-Leite, Robert M. Ewers, Jean-Paul Metzger, « Edge effects as the principal cause of area effects on birds in fragmented secondary forest », Oikos, vol. 119, no 6, , p. 918-926 (DOI 10.1111/j.1600-0706.2009.18061.x, lire en ligne [PDF])
- Lindenmayer, D. B. et Fischer, J. 2007. Tackling the habitat fragmentation panchreston. Trends in Ecology & Evolution 22, 127-132 (Résumé)
- source : OFFH ; Observatoire de la faune, de la flore et de ses habitats
- (en) Lenore Fahrig, « Ecological Responses to Habitat Fragmentation Per Se », Annual Review of Ecology, Evolution, and Systematics, vol. 48, no 1, , p. 1–23 (ISSN 1543-592X et 1545-2069, DOI 10.1146/annurev-ecolsys-110316-022612, lire en ligne, consulté le )
- EUROSTAT, Réseaux régionaux de transport ferroviaire et routier Les densités les plus fortes ne sont pas l'apanage des régions des capitales, Statistiques en bref – Transports – Fin de rédaction: 13.03.2008 ; Données extraites le: 15.11.2007; ISSN 1977-0332
- European Environment Agency, Landscape fragmentation in Europe ; Joint EEA-FOEN report, 2011 — 87 pp. — 21 x 29,7 cm ; (ISBN 978-92-9213-215-6) ; EEA Report series: ISSN 1725-9177 ; doi:10.2800/78322, coécrit avec l'Office fédéral suisse pour l'environnement (=Foen)
- Ilkka Hanski, Ilik Saccheri, « Molecular-Level Variation Affects Population Growth in a Butterfly Metapopulation », PLoS Biology, vol. 4, no (5): e129, (DOI 10.1371/journal.pbio.0040129)
- Hanski I (1999) Metapopulation ecology New York: Oxford University Press. 313 p.
Nieminen M, Siljander M, Hanski I (2004) Structure and dynamics of Melitaea cinxia metapopulations. In: Ehrlich PR, Hanski I, editors. On the wings of Checkerspots: A model system for population biology New York: Oxford University Press. p. 63–91. - Hanski, T Pakkala, M Kuussaari ; Metapopulation persistence of an endangered butterfly in a fragmented landscape ; (résumé PDF ; - Oikos, Copenhague, 1995 - JSTOR
- Forman, Richard T. T., et Michel Godron. 1986. Landscape Ecology. Wiley, New York.
- Forman, R.T.T. 1995. Land Mosaics: The Ecology of Landscapes and Regions. Cambridge University Press, Cambridge, Angleterre.
- Vos C.C., Verboom J., Opdam P.F.M. et Ter Braak C.J.F. 2001. Toward ecologically scaled landscape indices. The American Naturalist 183: 24–41.
- Opdam, P. (1991) Metapopulation theory and habitat fragmentation: a review of holarctic breeding bird studies. Landscape Ecology 5 (2): 93-106.
- Haila & Hanski 1984; Sisk & Haddad 2002
- Haila & Hanski 1984; Wilcove et al. 1986; Saunders et al. 1991; Debinski & Holt 1999
- Gascon & Lovejoy 1998; Gilfedder & Kirkpatrick 1998; Ås 1999; Jules et al. 1999; Kemper et al. 1999; Jules & Shahani 2003
- Amezaga, J.M., Santamaría, L. & Green, A.J. (2002) Biotic wetland connectivity — supporting a new approach for wetland management policy. Acta Oecologica, 23, 213–222.
- Lenore Fahrig et Gray Merriam, « Conservation of Fragmented Populations », Conservation Biology, vol. 8, no 1, , p. 50-59 (DOI 10.1046/j.1523-1739.1994.08010050.x, lire en ligne [PDF])
- Bailey, S.A., Duggan, I.C. & van Overdijk, C.D.A. (2003) Viability of invertebrate diapausing eggs collected from residual ballast sediment. Limnology and Oceanography, 48, 1701– 1710.
- Karen M. Alofs et Norma L. Fowler ; Habitat fragmentation caused by woody plant encroachment inhibits the spread of an invasive grass ; Volume 47 Issue 2, Pages 338 - 347 Online:2010/02/17 (Résumé)
- carte : Fragmentation des cœurs de forêts en Europe, AEE, 2010]
- http://pdbff.inpa.gov.br/iprojand4.html#2
- Dai Fukui, Toshihide Hirao, Masashi Murakami et Hirofumi Hirakawa ; Effects of treefall gaps created by windthrow on bat assemblages in a temperate forest ; ; Forest Ecology and Management Volume 261, Issue 9, 1 May 2011, Pages 1546-1552 (Résumé)
- Song sparrow (Melospiza melodia) song varies with urban noise (The Auk 123(3):650–659, 2006 American Ornithologists’ Union, 2006. Biology Department, Reed College, 3203 SE Woodstock Boulevard, Portland, Oregon 97202, USA.
- Zachos F.E., ALlthoff C., Steynitz Y.V., Eckert I., Hartl G.B. 2007. Genetic analysis of an isolated red deer (Cervus elaphus) population showing signs of inbreeding depression. European Journal of Wildlife Research 53 : 61-67 (7 p., 2 fig., 2 tab., 61 réf.). Pour lire l’article : European Journal of Wildlife Research (www.springerlink.com/content/1439-0574/
- Exemple illustré : Fragmentation par les infrastructures dans le nord de la France, et bases du réseau écologique (Trame verte)] sur ce même territoire (La Voix du Nord)
- Patrizia Digiovinazzo, Gentile Francesco Ficetola, Luciana Bottoni, Carlo Andreis, Emilio Padoa-Schioppa, " Ecological thresholds in herb communities for the management of suburban fragmented forests" ; Forest Ecology and Management ; Volume 259, Issue 3, 25 January 2010, Pages 343–349 (Résumé)
- Jean & Bouchard 1993; Findlay & Houlahan 1997
- MacArthur & Wilson 1967; Wilson & Willis 1975; Simberloff & Abele 1976; Gilpin & Diamond 1980
- Soulé 1991
- Fernando Pimentel-Souza, Virgínia Torres Schall, Rodolfo Lautner Jr., Norma Dulce Campos Barbosa, Mauro Schettino, Nádia Fernandes, Behavior of Biomphalaria glabrata (Gastropoda: Pulmonata) under different lighting conditions ; Revue canadienne de zoologie, 1984, 62:2328-2334, 10.1139/z84-340
- Source Jean-marc Elouard et Chritian Lévêque, Rythme nycthéméral de dérive des insectes et des poissons dans les rivières de côte d'Ivoire, Laboratoire d’hydrobiologie, ORSTOM, Bouaké (Côte d'Ivoire)
- KERTH G., MELBER M. [2009]. Speciesspecific barrier effects of motorway on habitat use of two threatened forest-living bat species. Biological Conservation 142(2) : 270-279 (10 p., 2 fig., 3 tab., 51 réf.).
- N. de Saadeler, J.M. Fauconnier, G. Kustjens, G. Berthoud, R.J. Cooper, Études relatives au transport et à la diversité biologique et paysagère (Sauvegarde de la nature no 132) ; Éditions Conseil de l'Europe ; 2003-O1-01 ; 134 pages, (ISBN 978-92-871-5277-0) (Lien)
- Code de pratiques sur la prise en compte de la diversité biologique et paysagère dans les infrastructures de transport (Sauvegarde de la nature no 131), Éditions Conseil de l'Europe, 85p., (ISBN 978-92-871-5114-8) (Lien)
- Commissariat général au développement durable Datar, Observatoire des territoires, Service de l’observation et des statistiques La fragmentation des espaces naturels
- Décret n° 2012-1492 du 27 décembre 2012 relatif à la trame verte et bleue,Art. R. 371-20.-I
{kind=link}
{kind=link}
{kind=link}
 Portail de l’écologie
Portail de l’écologie  Portail de la conservation de la nature
Portail de la conservation de la nature