Psilocybe semilanceata
Psilocybe semilanceata, psilocybe lancéolé ou psilocybe fer de lance, est une espèce de champignons qui contient des substances psychoactives qui sont la psilocybine et la baeocystine. Il est qualifié comme un champignon hallucinogène ou champignon psychédélique. Il est à la fois un des champignons à psilocybine les plus présents dans la nature, et un des plus puissants.

| Règne | Fungi |
|---|---|
| Division | Basidiomycota |
| Classe | Agaricomycetes |
| Sous-classe | Agaricomycetidae |
| Ordre | Agaricales |
| Famille | Strophariaceae |
| Genre | Psilocybe |
Le Psilocybe semilanceata est l'espèce type du genre Psilocybe[1]. Ce petit champignon très répandu dans le monde est constitué d’un chapeau visqueux ne dépassant pas 2,5 cm et surmonté d’un mamelon proéminent. Son chapeau forme un dégradé de jaune à brun, couvert de rainures radiales lorsqu'il est humide, et prend une couleur plus claire au fur et à mesure qu'il vieillit et fane. Le stipe a tendance à être mince et long, ainsi que de la même couleur ou légèrement plus clair que le chapeau. La fixation des lamelles au stipe est adnexée (étroitement attachée). Les lamelles sont d'abord de couleur crème avant de se colorer en pourpre dès que les spores mûrissent. Les spores sont en masse d'un brun violacé sombres, de forme ellipsoïde, et mesurent 10,5-15 par 6,5-8,5 micromètres.
Ce champignon pousse dans les prairies herbeuses, et habitats similaires, particulièrement humides, qui sont fertilisés par du bétail. C'est une espèce saprophyte qui se nourrit des cellules moribondes de racines de différentes espèces d'herbes. À l'inverse du Psilocybe cubensis, cette espèce ne pousse pas dans des déjection d'animaux ni ne s'en nourrit. Il est largement répandu dans les régions à climat tempéré de l’hémisphère Nord, particulièrement en Europe.
Le cas le plus ancien connu d'intoxication au Psilocybe semilanceata a été constaté en 1799 à Londres. Sa consommation volontaire a commencé dans les années 1960 lorsque le champignon a été déclaré la première espèce européenne contenant de la psilocybine.
La possession ou la vente de champignons à psilocybine est illégale dans beaucoup de pays.
Taxinomie et étymologie

En 1838, Elias Magnus Fries décrit l'espèce pour la première fois dans son Epicrisis Systematics Mycologici avec comme nom binominal original Agaricus semilanceatus[2]. En 1871, Paul Kummer transfère l'espèce dans le genre Psilocybe après avoir élevé au rang de genre de nombreux subdivisions d'Agaricus[3]. Le taxon Panaeolus semilanceatus décrit par Jakob Emanuel Lange dans des publications de 1936 et 1939 est considéré comme un synonyme[4],[5]. Selon la base de données taxinomique MycoBank, plusieurs taxons ayant été considérés des variétés de P. semilanceata sont depuis devenus synonymes de Psilocybe strictipes[6] : c'est le cas des variétés caerulescens décrite par Pier Andrea Saccardo en 1887 (originellement nommée Agaricus semilanceatus Var. coerulsecens par Mordecai Cubitt Cooke en 1881), microspora décrite par Rolf Singer en 1969, et obtusata décrite par Marcel Bon en 1985[7],[8],[9].
Plusieurs études de phylogénétique moléculaire publiées au cours des années 2000 ont démontré que le taxon Psilocybe, tel qu'il se définissait alors, était polyphylétique[10],[11],[12]. Ces études allaient en faveur d'une division du genre en deux clades distincts, l'un regroupant les espèces hallucinogènes qui bleuissent dans la famille Hymenogastraceae, et l'autre les espèces non-hallucinogènes et ne bleuissant pas au sein de la famille Strophariaceae. Cependant, le lectotype[13] du genre le plus communément accepté était alors Psilocybe montana, une espèce non-hallucinogène et ne bleuissant pas. Si ces espèces non-hallucinogènes étaient classées dans un autre taxon, le genre avec les espèces hallucinogènes aurait été dépourvu de nom valide. Afin de contrer ce dilemme, plusieurs mycologues proposent en 2005 de conserver le nom de genre Psilocybe avec comme espèce-type P. semilanceata. Ainsi, la conservation du nom permettrait d'éviter un grand nombre de modifications nomenclaturales au sujet de ce groupe de champignons dont de nombreuses espèces se retrouvent liées « à l'archéologie, à l'anthropologie, à la religion, aux modes de vie alternatifs, à la science forensique, au droit et à la légalité de la consommation »[14]. Le choix de P. semilanceata comme espèce-type trouvait également une justification dans son acceptation en tant que lectotype par de nombreux auteurs au cours de la période 1938-68. Cette proposition de conserver le nom Psilocybe avec l'espèce P. semilanceata comme type est acceptée à l'unanimité par le Comité de nomenclature mycologique (en) en 2009[1].
En français, les noms vernaculaires du champignon, psilocybe lancéolé ou psilocybe fer de lance, sont de simples traductions du nom scientifique. En anglais, le nom vernaculaire, liberty cap (soit « chapeau de la liberté») fait référence au bonnet phrygien auquel il ressemble[15] ; il partage ce nom vernaculaire avec P. pelliculosa, une espèce à l'apparence extrêmement proche[16],[17]. Le mot latin pour désigner le bonnet phrygien est pileus, ce mot est depuis utilisé dans le vocabulaire spécialisé pour désigner le chapeau du sporophore (ou carpophore), c'est-à-dire le fruit du champignon. Au cours du XVIIIe siècle, les bonnets phrygiens étaient placés sur des poteaux de la liberté, notamment au cours de la guerre d'indépendance des États-Unis : ces poteaux ont une apparence comparable au stipe du champignon. Le nom du genre est dérivé du grec ancien psilos (ψιλός) (« doux » ou « nu ») et du grec byzantin kubê (κύβη) (« tête »)[18],[19]. L'épithète spécifique vient du latin semi (« moitié ») et lanceata, de lanceolatus, qui signifie « en forme de lance »[20].
Description

Le chapeau de P. semilanceata mesure entre 5 et 25 mm de diamètre et de 6 à 22 mm de hauteur. Il adopte une forme conique et pointue ou bien une forme de cloche, il est alors dit campanulé ; la forme reste plus ou moins la même au cours de la vie d'un spécimen. Une papille, l'umbo, est souvent visible au sommet du chapeau. La bordure du chapeau est enroulée vers l'intérieur avant de se déplier voire de s'enrouler vers l'extérieur au fur et à mesure que le spécimen mûrit. Le chapeau est hygrophane : sa teinte se modifie selon son niveau d'humidité. Humide, le chapeau prend une teinte ocre, brun pâle ou noisette ; le centre est plus sombre avec des nuances proches du vert d'eau. C'est également lorsque le champignon est humide que les rainures radiales correspondant aux lames s'aperçoivent sur la cuticule. Sec, le chapeau est beaucoup plus pâle, d'une teinte marron jaunâtre[21]. La cuticule d'un spécimen humide est collante en raison d'un film gélatineux ; la cuticule est dite glutineuse[22]. Ce film devient visible lorsqu'une partie du chapeau est cassée et pend, tendant ainsi le film. Quand le chapeau sèche après une exposition au soleil, le film blanchit et ne pèle plus[23].
La partie inférieure du chapeau renferme entre 15 et 27 lames étroites regroupées sans être collées les unes aux autres ; ces lames sont adnées, c'est-à-dire rattachées au stipe par une jointure très fine voire inexistante et semblent faire corps avec lui. Les lames des jeunes spécimens sont brun pâle et deviennent gris sombre à marron violacé quand les spores parviennent à maturité. Le stipe est élancé et d'une teinte marron jaunâtre : il mesure entre 45 et 140 mm de long pour 1 à 3,5 mm de diamètre ; le diamètre est souvent plus important à proximité de la base[21]. Le champignon porte un voile fin ressemblant à une toile d'araignée qui ne le couvre pas entièrement et qui disparaît rapidement. Un anneau est parfois présent sur la partie supérieure du stipe ; celui-ci est assombri par l'empreinte de spores[22]. La chair est fine et membraneuse, d'une couleur similaire à celle de la cuticule. Son odeur et son goût sont comparables à de la farine fraîchement moulue. Toutes les parties du champignon prennent une teinte bleuâtre si abîmées ou manipulées ; cette teinte survient parfois naturellement avec l'âge[21].
Caractéristiques microscopiques
L'empreinte de spores adopte une teinte quelque part entre le marron-violet et le rouge profond. L'utilisation d'un microscope optique permet d'obtenir plus de détails : observés frontalement, les spores ont une forme oblongue ou ovale ; ils sont longs de 10,5 à 15 µm et larges de 6,5 à 8,5 μm. Les basides[24] mesurent entre 20 et 31 μm de long pour 5 à 9 μm de large, ils disposent de petites pointes permettant à un baside unique de porter quatre spores. Les basides ne sont pas présents sur le bord stérile des lamelles. Les cheilocystides (cystide située sur l'arête d'une lame) mesurent de 15 à 30 μm de long pour 4 à 7 μm de large ; elles ont une forme de flacon avec un goulot long et large d'environ 1 à 3 μm. P. semilanceata ne porte pas de pleurocystides, c'est-à-dire des cystides présentes sur la façade d'une lame. La cuticule du chapeau peut atteindre une épaisseur de 90 µm, elle se compose d'une couche de tissu appelée l'ixocutis : il s'agit d'une couche d'hyphe gélatineux et s'étendant parallèlement à la surface du chapeau. Les noyaux d'hyphe abritant l'ixocutis sont cylindriques hyalins ; ils sont larges de 1 à 3,5 mm. Le subpellis, une autre couche du chapeau, se trouve juste en-dessous de la cuticule : il se compose de noyaux d'hyphe larges de 4 à 12 μm aux parois rugueuses et de couleur marron-jaunâtre. Des anses d'anastomose sont formées dans chacun des tissus du champignon comprenant des noyaux d'hyphe[22].
| Caractéristiques microscopiques | |||||||||
| |||||||||



Cas de polymorphisme
Le champignon P. semilanceata est sujet au polymorphisme génétique, c'est-à-dire qu'il se présente sous différentes formes. La forme anamorphique de P. semilanceata est une étape de la reproduction asexuée du cycle de vie du champignon qui est impliquée dans le développement de diaspores mitotiques : les conidies. Cultivé dans une boîte de Petri, le champignon prend la forme d'une boule cotonneuse de mycélium : la teinte adoptée est blanche à orange pâle. Les conidies ainsi obtenues sont droites ou courbées : elles mesurent de 2 à 8 μm par 1,1 à 2 μm et contiennent une ou plusieurs petites structures intracellulaires[25]. Si cette étape asexuée de la vie du champignon est peu connue en dehors de la culture en laboratoire, la morphologie de ces structures asexuées sert tout de même de caractéristique classique lors d'analyses phylogénétiques afin de comprendre les relations évolutives entre des groupes de champignons liés[26].
Le mycologue écossais Roy Watling a décrit des spécimens gastéroïdes (similaires à des truffes) et sécotioïdes de P. semilanceata trouvés poussant en association avec des fructuations à la forme habituelle. Ces spécimens disposent d'un chapeau allongé, long de 20 à 22 cm et large de seulement 0,8 à 1 cm à la base ; la bordure inférieure est tournée vers l'intérieur du chapeau et rejoint le stipe par l'intermédiaire d'excroissances membraneuses. Les lames sont étroites, très peu espacées les unes par rapport aux autres et comportent de nombreuses anses d'anastomose pouvant suggérer un réseau de veines. Ces lames sont de couleur sépia, elles arborent une coulure bordeaux et une bordure blanche. Les stipes des fructuations sont longs de 5 à 6 cm et larges de 0,1 à 0,3 cm ; le stipe est couvert par l'anneau qui forme l'extension du chapeau sur environ 2 cm. Les spores ont une paroi épaisse : ils mesurent entre 12,5 et 13,5 μm par 6,5 à 7 μm. Malgré les différences morphologiques importantes, des analyses moléculaires ont démontré que la version sécotioïde appartenait bien à la même espèce que le morphotype original[27].
Espèces proches

Plusieurs espèces issues du genre Psilocybe peuvent se confondre avec P. semilanceata en raison d'une apparence extérieure approchante. P. strictipes est une espèce qui pousse en prairie ; elle est fine mais de distingue visuellement de P. semilanceata par l'absence d'une papille proéminente. P. mexicana est également d'une apparence similaire mais ne se rencontre que dans des prairies subtropicales mexicaines au sol riche en fumier. Ses spores sont légèrement plus petits que ceux de P. semilanceata avec une taille de 8 à 9,9 μm par 5,5 à 7,7 μm[28]. L'espèce P. samuiensis pousse en Thaïlande dans des sols argileux et bien pourvus en fumier ou au milieu de rizières. Ce champignon se différencie de P. semilanceata par un chapeau plus petit, jusqu'à 1,5 cm de diamètre seulement, et par ses spores rhomboïdes[29]. L'espèce la plus ressemblante à l’œil nu est P. pelliculosa ; seule une mesure précise des spores permet de différencier les deux espèces entre elles : les spores de P. pelliculosa sont plus petits, avec une taille de 9 à 13 μm par 5 à 7 μm[17].

P. semilanceata présente également des risques de confusion avec l'espèce Inocybe geophylla qui contient de la muscarine, un alcaloïde toxique[30]. Ce champignon est blanchâtre, le chapeau est soyeux, les lames ont une teinte comprise entre le marron jaunâtre et le gris clair, le dépôt de spores est marron-jaunâtre terne[31].
Écologie et habitat
Les fruits de P. semilanceata poussent seuls ou en groupes sur des sols riches et acides[22],[32] : on les trouve sur des pelouses, des prairies, des pâturages ou sur du gazon[33],[34],[35],[36]. Ils se rencontrent souvent sur des prairies fertilisées par les excréments de moutons ou de vaches sans pour autant pousser directement sur les bouses[22],[34].

Comme d'autres espèces de champignons hallucinogènes telles que P. mexicana, P. tampanensis, ou Conocybe cyanopus, P. semilanceata développe parfois une phase sclérote, c'est-à-dire une forme dormante du champignon qui le protège des incendies et d'autres catastrophes naturelles[35]. Le mycologue allemand Jochen Gartz est parvenu à reproduire la phase de sclérote dans de la gélose à l'extrait de malt[37].
P. semilanceata, comme toutes les autres espèces du genre Psilocybe, est un champignon saprophyte : il obtient ses nutriments en décomposant la matière organique[38],[39]. Le champignon est également rencontré en association avec des laîches dans les zones humides des champs, et a été observé poussant sur des racines en décomposition[22],[40],[41].
Des expériences en laboratoire ont prouvé la capacité de P. semilanceata à empêcher l'expansion de l'oomycète Phytophtora cinnamoni, un protiste pathogène virulent qui entraîne une pourriture racinaire chez les plantes qu'elle attaque[42]. Lorsque l'espèce est cultivée en laboratoire en compétition avec d'autres espèces de champignons saprophytes également isolés de la rhizosphère de leur habitat naturel, P. semilanceata réduit significativement leur croissance. Cette propriété antifongique est au moins partiellement liée à deux composés phénoliques sécrétés par l'espèce. Cela lui permet une compétitivité accrue face aux autres espèces fongiques dans la recherche de nutriments à partir de la matière de plantes décomposées[43]. Une autre expérience de laboratoire s'appuyant sur des tests de sensibilité aux antibiotiques (en) a permis de prouver l'efficacité de P. semilanceata contre le staphylocoque doré résistant à la méticilline (SARM), c'est-à-dire une variation de ce pathogène caractérisée par sa résistance à cet antibiotique. L'origine de cette activité antimicrobienne est inconnue[44].
Distribution
Gastón Guzmán, le mycologue faisant autorité sur le genre Psilocybe, dans sa monographie sur les champignons à psilocybine parue en 1983, estime que P. semilanceata est le champignon contenant de la psilocybine possédant la distribution la plus large puisque sa présence avait alors été confirmée dans 17 pays[45]. En Europe, P. semilanceata est très largement répandu : il se rencontre en Islande, aux îles Féroé, en Irlande, au Royaume-Uni, sur les îles Anglo-Normandes, en France, en Espagne, en Italie en Belgique, en Autriche, en Allemagne, en Suisse, en République Tchèque, en Slovaquie aux Pays-Bas, au Danemark, en Suède, en Norvège, en Finlande, en Grèce, en Hongrie, en Bulgarie, en Géorgie, en Estonie, en Pologne, en Lituanie et en Russie[46]. Il est généralement accepté que l'espèce est originaire d'Europe[47]. Watling a démontré qu'il existe très peu de différences entre les spécimens récoltés en Espagne et ceux récoltés en Écosse, tant aux niveaux morphologique que génétique[27].
Le champignon est également largement distribué en Amérique du Nord. Au Canada, sa présence est enregistrée en Colombie-Britannique, au Nouveau-Brunswick, en Terre-Neuve-et-Labrador, en Nouvelle-Écosse, sur l'île-du-Prince-Édouard, en Ontario et au Québec[46]. Aux États-Unis, il est commun dans le Nord-Ouest Pacifique, à l'ouest de la chaîne des Cascades, où il pousse abondamment en automne et au début de l'hiver ; des fructifications éparses ont également été observées au printemps[22]. Charles Horton Peck a affirmé que le champignon était présent dans l'État de New York au début du XXe siècle : de nombreuses publications scientifiques émises postérieurement ont donc indiqué que l'espèce se rencontrait aussi à l'Est des États-Unis. Dans sa monographie de 1983 sur les Psilocybe, Gastón Guzmán étudie le spécimen se trouvant dans l'herbier de Peck et conclut à une confusion avec l'espèce aujourd'hui connue comme Panaeolina foenisecii[47],[48]. En Amérique du Sud, l'espèce est bien moins commune mais sa présence est tout de même enregistrée au Chili[46],[47]. En Océanie, le champignon est présent en Australie, où il pourrait être une espèce introduite[27], et en Nouvelle-Zélande ; il pousse sur les prairies de haute altitude[49]. En 2000, la présence du champignon est remarquée à Golaghat, dans l'État d'Assam, en Inde[50].
Utilisation psychoactive

Le premier témoignage faisant état avec certitude d'une intoxication à Psilocybe semilanceata remonte à 1799 et concerne une famille britannique ayant préparé un repas à base de champignons récoltés au Green Park, à Londres. Selon le chimiste Augustus Everard Brande, le père et ses quatre enfants ont subi les symptômes habituels associés à l'ingestion, parmi lesquels la dilatation des pupilles, le fou rire spontané et les phases de délire[51]. L'identification de l'espèce concernée est rendue possible en 1803 grâce à l'ouvrage de James Sowerby Coloured Figures of English Fungi or Mushrooms : ce livre contient une description du champignon, alors connu sous le nom Agaricus glutinosus, selon la publication de 1780 de Moses Ashley Curtis[52]. Le mycologue allemand Jochen Gartz considère que la description de l'espèce est « absolument compatible avec le savoir actuel concernant Psilocybe semilanceata »[53].
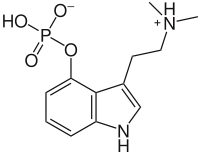
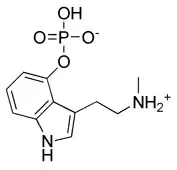
Au début des années 1960, le scientifique suisse Albert Hofmann, principalement connu pour avoir synthétisé le LSD, réalise une analyse chimique de fructuations de P. semilanceata récoltés en Suisse et en France par le botaniste Roger Heim. Grâce à la technique de la chromatographie sur papier, Hofmann relève une concentration de 0,25 % de psilocybine (par rapport à la masse du fruit séché) à partir de spécimens séchés. En 1963, la publication qui fait connaître ces résultats est la première à prouver la présence de psilocybine dans une espèce de champignons européenne ; jusque-là, cette molécule était seulement connue dans les espèces de Psilocybe originaires du Mexique, d'Asie et d'Amérique du Nord[33]. Cette découverte est confirmée à plusieurs reprises : d'abord à la fin des années 1960 avec des spécimens venus d'Écosse et d'Angleterre[54],[55], de Tchécoslovaquie en 1973[56], d'Allemagne en 1977[57], de Norvège en 1978[40], et de Belgique et de Finlande en 1984[58],[59]. En 1965, des champignons saisis auprès d'étudiants de Colombie-Britannique subissent une analyse de chimie forensique (en) : l'identification de ces champignons comme P. semilanceata fait de cette affaire le premier cas d'utilisation récréative et intentionnelle de l'espèce au Canada[60],[61]. En 1977, une étude affirme la présence de la béocystine, un analogue structurel de la psilocybine[57]. D'autres études publiées à la fin du XXe siècle et au cours des années 2000 indiquent qu'il n'existe qu'une très faible variation de la concentration de psilocybine selon l'origine des spécimens de P. semilanceata[62],[63],[64],[65].
Propriétés
La quantification des composés hallucinogènes se trouvant dans les fruits de P. semilanceata a fait l'objet de plusieurs études. En 1993, Gartz indique une concentration moyenne de psilocybine de 1 % (toujours par rapport à la matière sèche) : la concentration minimale est à 0,2 %, la concentration maximale s'élève à 2,37%, ce qui est la concentration en psilocybine la plus forte jamais rencontrée dans un champignon[66]. En 1985, Tjakko Stijve et Thom Kuyper avaient relevé une concentration élevée dans un unique spécimen (1,7 %) à laquelle s'ajoutait une concentration assez forte de béocystine (0,36 %)[67]. Les spécimens plus petits disposent souvent des concentrations les plus élevées, mais les valeurs absolues les plus hautes se rencontrent dans des spécimens de grande taille[68]. Une étude finnoise a essayé les concentrations de psilocybine dans des spécimens issus de vieux herbiers : elle conclut que la concentration décroît de façon linéaire au cours du temps mais qu'elle reste relativement stable. Ainsi, la molécule a été détectée dans des spécimens âgés de 115 ans[69]. Une analyse de spécimens de la côte pacifique nord-ouest menée par Michael Beug et Jeremy Bigwood a permis de relever des concentrations comprises entre 0,62 % et 1,28 %, avec une moyenne a 1.0 ±0.2 %. Beug et Bigwood concluent que l'espèce est l'une des plus puissantes, ainsi que la plus constante dans les niveaux de concentration en psilocybine[70]. Dans une publication de 1996, Paul Stamets propose d'établir une « échelle d'évaluation de la puissance » reposant sur la valeur des composés psychoactifs (psilocybine, psilocine, béocystine) et s'attachant à douze espèces de Psilocybe. Cette proposition permet de comparer de façon basique la puissance de différentes espèces ; cette technique est imparfaite puisqu'elle ne fait pas la différence entre les propriétés psychoactives des différents composés chimiques. Malgré sa petite taille, P. semilanceata est considéré comme un champignon hallucinogène "modérément actif à extrêmement puissant" : cela correspond à un pourcentage combiné des composés psychoactifs compris entre 0,25 % et plus de 2 %[22]. Selon l'échelle de Stamets, des douze espèces comparées, seules trois sont plus puissantes que P. semilanceata : il s'agit de P. azurescens, P. baeocystis et P. bohemica[71]. En 1995, Gartz affirme que P. semilanceata est l'espèce psychoactive la plus répandue en Europe[36].
Plusieurs rapports scientifiques visant à documenter les effets de la consommation de P. semilanceata ont été publiés. Les symptômes habituels comprennent des distorsions visuelles attachées à la couleur, la profondeur et la forme ; cela peut aller jusqu'à des hallucinations visuelles. Les effets sont comparables à ceux qui résultent de la consommation de LSD, bien que plus faibles[72]. Les effets secondaires de l'ingestion du champignon consistent en une dilatation des pupilles, un rythme cardiaque accru, une mauvaise humeur, et une exagération des réflexes. Comme généralement, les symptômes liés à l'ingestion de champignons à psilocybine varient grandement selon les individus : « l'effet sur l'humeur en particulier dépend des traits de personnalité du sujet précédemment à l'ingestion », et « des doses identiques de psilocybine peuvent entraîner des effets très différents chez des individus différents »[73]. La consommation n'entraîne pas d'addiction mais des troubles psychologiques peuvent se déceler[74]. À ce sujet, afin de réduire les risques lors des recherches expérimentales, les chercheurs en psychopharmacologie de l'université Johns-Hopkins ont établi un protocole qui déconseille l'usage de la psilocybine chez les personnes souffrant de schizophrénie ou ayant des cas de psychoses au niveau des deux premiers degrés de parenté. Il exclut aussi les personnes atteintes de troubles bipolaires ou les individus présentant des traits d'humeur trop labiles ou au contraire trop rigides[75].
La grande majorité des intoxications se résout sans incident mais de rares cas font état de conséquences sévères, principalement en raison d'un dosage élevé ou d'un usage persistant. En Pologne, en 1998, un jeune homme de 18 ans a développé le syndrome de Wolff-Parkinson-White, de l'arythmie, et a souffert d'un infarctus du myocarde après avoir ingéré P. semilanceata fréquemment sur la période d'un mois. Il est suggéré que les causes ayant entraîné les dommages cardiaques et l'infarctus du myocarde sont soit une vasoconstriction coronaire, soit une hyper-agrégation des plaquettes sanguines et une obstruction des petites artères coronaires[76].
En 1998, une étude prouve la présence de la phényléthylamine dans des spécimens récoltés en Suède. Les taux de concentration de la phényléthylamine, une substance active décarboxylisée issue de l'acide aminé phénylalanine, sont très largement variables ; un spécimen se détache avec une concentration de 146 µg par gramme de champignon (sur un spécimen encore frais et humide)[77]. Ce composé intéresse grandement la recherche psychiatrique puisqu'il pourrait s'agir d'un neuromodulateur de synapses aminergiques : c'est-à-dire de neurones qui utilisent les monoamines comme neurotransmetteurs. La phényléthylamine posséderait également des propriétés énergisantes, améliorerait l'humeur et encouragerait l'agression[78]. Les auteurs de l'étude suggèrent l'existence d'une interaction entre la phényléthylamine et la psilocine (un composé chimique simple issu de la décomposition de la psilocybine) à travers l'inhibition compétitive d'enzymes monoamines oxydases[77].
L'un des dangers principaux de la tentative de consommation de champignons hallucinogènes ou d'autres champignons sauvages, surtout pour les cueilleurs novices, est la possibilité de confusion avec des espèces toxiques entraînant l'empoisonnement. Ainsi, un jeune Australien en bonne santé a confondu P. semilanceata avec le champignon vénéneux Cortinarius rubellus (en). Le jeune homme a souffert d'insuffisance rénale et a nécessité une greffe de rein[79]. Dans un autre cas, un jeune homme a développé des anormalités cardiaques similaires à celles rencontrées dans le syndrome de tako-tsubo et se caractérisant par un affaiblissement temporaire mais soudain du myocarde[80]. Un procédé de réaction en chaîne par polymérase est mis en place par des scientifiques polonais en 2007 afin d'identifier spécifiquement P. semilanceata[81].
| Structures chimiques des composés | |||||||||
| |||||||||

Statut légal
Le statut légal des champignons à psilocybine est différent selon les pays. La psilocybine et la psilocine sont listées dans la Convention sur les substances psychotropes de 1971 de l'ONU[82]. Dans les pays ayant ratifié ce texte, la possession et l'utilisation de champignons à psilocybine, notamment P. semilanceata, sont donc interdites. Si plusieurs pays d'Europe avaient maintenu la tolérance vis-à-vis de l'usage et de la possession des champignons hallucinogènes même après l'interdiction américaine, la législation s'est faite plus sévère au cours des années 2000. Alors que les champignons étaient auparavant vendus légalement dans les coffee shops et smartshops licenciés, les Pays-Bas sont le dernier pays européen à interdire la possession et l'usage de champignons psychédéliques en 2008[83].
Annexes
Bibliographie
- (en) Guzmán G., The genus Psilocybe: A Systematic Revision of the Known Species Including the History, Distribution and Chemistry of the Hallucinogenic Species, vol. 74, Berlin, J. Cramer, (ISBN 3-7682-5474-7).
- (en) A. Bresinsky et H. Besl, A Colour Atlas of Poisonous Fungi : a Handbook for Pharmacists, Doctors, and Biologists, Londres, Royaume-Uni, Manson Publishing Ltd., (ISBN 0-7234-1576-5).
- (en) P. Stamets, Psilocybin Mushrooms of the World : An Identification Guide, Berkeley, Californie, Ten Speed Press, (ISBN 0-89815-839-7).
- (en) J. Gartz, Magic Mushrooms Around the World, Los Angeles, Californie, LIS Publications, , 136 p. (ISBN 978-0-9653399-0-2).
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Psilocybe semilanceata » (voir la liste des auteurs).
- (en) L. Norvell, « Report of the Nomenclature Committee for Fungi: 15 », Mycotaxon, vol. 110, , p. 487–92 (DOI 10.5248/110.487, lire en ligne[archive du ] [PDF]).
- (la) Fries EM, Epicrisis Systematis Mycologici : Seu Synopsis Hymenomycetum, Uppsala, Suède, Typographia Academica, (lire en ligne), p. 231.
- (de) Kummer P., Der Führer in die Pilzkunde, Zerbst, Germany, C. Luppe, , p. 71.
- (en) J.E. Lange, « Studies in the Agarics of Denmark, Part XI. Psathyra, Panaeolus, Psilocybe, Gomphidius, Phylloporus, Cantharellus, Schizophyllum », Dansk botanisk Arkiv, vol. 9, no 1, , p. 1–46.
- (en) J.E. Lange, Flora Agaricina Danica, vol. 4, Copenhague, Danemark, Danish Botanical Society, , p. 84.
- (en) « Psilocybe strictipes Singer & A.H. Sm. 1958 », sur MycoBank (consulté le ).
- (en) Saccardo PA, Sylloge Hymenomycetum, Vol. I. Agaricineæ, vol. 5, Padoue, Italie, , p. 1051.
- (en) R. Singer, « Mycoflora australis », Beihefte zur Nova Hedwigia, vol. 29, , p. 247.
- M. Bon, « Novitates – Validations de taxons », Documents Mycologiques, vol. 16, no 61, , p. 46.
- (en) J.M. Moncalvo, R. Vilgalys, S.A. Redhead, J.E. Johnson, T.Y. James, M. Catherine Aime, V. Hofstetter, S.J. Verduin, E. Larsson, T.J. Baroni, R. Greg Thorn, S. Jacobsson, H. Clémençon et O.K. Miller Jr., « One hundred and seventeen clades of euagarics », Molecular Phylogenetics and Evolution, vol. 23, no 3, , p. 357–400 (DOI 10.1016/S1055-7903(02)00027-1).
- (en) K.G. Nugent et B.J. Saville, « Forensic analysis of hallucinogenic fungi: a DNA-based approach », Forensic Science International, vol. 140, nos 2–3, , p. 147–57 (DOI 10.1016/j.forsciint.2003.11.022).
- (en) P.B. Matheny, J.M. Curtis, V. Hofstetter, M.C. Aime, J.M. Moncalvo, Z.W. Ge, J.C. Slot, J.F. Ammirati, T.J. Baroni, N.L. Bougher, K.W. Hughes, D.J. Lodge, R.W. Kerrigan, M.T. Seidl, D.K. Aanen, M. DeNitis, G.M. Daniele, D.E. Desjardin, B.R. Kropp, L.L. Norvell, A. Parker, E.C. Vellinga, R. Vilgalys et D.S. Hibbett, « Major clades of Agaricales: a multilocus phylogenetic overview », Mycologia, vol. 98, no 6, , p. 982–95 (DOI 10.3852/mycologia.98.6.982, lire en ligne [PDF]).
- Spécimen type désigné plus tard par un autre biologiste que l'auteur quand ce dernier n'a pas désigné de type.
- (en) S.A. Redhead, J.M. Moncalvo, R. Vilgalys, P.B. Matheny, L. Guzmán-Dávalos et G. Guzmán, « (1757) Proposal to conserve the name Psilocybe (Basidiomycota) with a conserved type », Taxon, vol. 56, no 1, , p. 255–57 (lire en ligne [PDF]).
- (en) A. Weil, The Marriage of the Sun and Moon : Dispatches from the Frontiers of Consciousness, Boston, Massachusetts, Houghton Mifflin, (ISBN 0-618-47905-8, lire en ligne), p. 77.
- (en) S.H. Pollock, « Liberty caps: recreational hallucinogenic mushrooms », Drug and Alcohol Dependence, vol. 1, no 6, , p. 445–47 (DOI 10.1016/0376-8716(76)90010-7).
- (en) G. Guzmán, J. Ott, J. Boydston et S.H. Pollock, « Psychotropic mycoflora of Washington, Idaho, Oregon, California and British Columbia », Mycologia, vol. 68, no 6, , p. 1267–72 (DOI 10.2307/3758763, lire en ligne).
- (en) « psilocybin, n. », sur Oxford Living Dictionary (consulté le ).
- (en) Cornelis S., Schrevelius' Greek Lexicon, Translated into English with Numerous Corrections, Londres, Royaume-Uni, Baldwin, Craddock, & Joy, (lire en ligne), p. 358.
- (en) R.S. Mehrotra et K.R. Aneja, An Introduction to Mycology, Columbia, Missouri, South Asia Books, , 766 p. (ISBN 81-224-0089-2, lire en ligne), p. 540.
- (en) C. Bas, Th. W. Kuyper, E.C. Vellinga et J. van Os, Flora Agaricina Neerlandica, vol. 3, Boca Raton, Floride, CRC Press, (ISBN 90-5410-616-6), p. 45.
- Stamets 1996, p. 142-145
- (en) B. Harris, Growing Wild Mushrooms : A Complete Guide to Cultivating Edible and Hallucinogenic Mushrooms, Berkeley, Californie, Ronin Publishing, , 96 p. (ISBN 1-57951-066-3, lire en ligne), p. 83.
- Ce sont les organes portant les spores et formant l'hyménium.
- (en) G. Walther et M. Weiss, « Anamorphs in the Strophariaceae (Basidiomycota, Agaricales) », Botany-Botanique, vol. 86, no 6, , p. 551–66 (DOI 10.1139/B08-036).
- (en) G. Walther, S. Garnica et M. Weiß, « The systematic relevance of conidiogenesis modes in the gilled Agaricales », Mycological Research, vol. 109, no 5, , p. 525–44 (DOI 10.1017/S0953756205002868).
- (en) R. Watling et M.P. Martin, « A sequestrate Psilocybe from Scotland », Botanical Journal of Scotland, vol. 55, no 2, , p. 245–57 (ISSN 1359-4869, DOI 10.1080/03746600308685009).
- Stamets 1996, p. 129
- Stamets 1996, p. 140
- Bresinsky et Besl 1989, p. 115-116
- (en) J. Ammirati, J.A. Traquair et P.A. Horgen, Poisonous Mushrooms of the Northern United States and Canada, Ottawa, Canada, Fitzhenry & Whiteside in cooperation with Agriculture Canada, (ISBN 978-0-88902-977-4, lire en ligne), p. 149.
- Guzmán 1983, p. 361
- A. Hofmann, R. Heim et H. Tscherter, « Phytochimie – présence de la psilocybine dans une espèce européenne d'agaric, le Psilocybe semilanceata », Comptes rendus hebdomadaires des séances de l'Académie des sciences, vol. 257, no 1, , p. 10–12.
- Gartz 1997, p. 21
- Stamets 1996, p. 24
- Gartz 1997, p. 18
- Gartz 1997, p. 56
- Guzmán 1983, p. 22
- Stamets 1996, p. 16
- (en) K. Høiland, « The genus Psilocybe in Norway », Norwegian Journal of Botany, vol. 25, no 2, , p. 111–22.
- (en) S.M. Keay et A.E. Brown, « Colonization by Psilocybe semilanceata of roots of grassland flora », Mycological Research, vol. 94, no 1, , p. 49–56 (DOI 10.1016/S0953-7562(09)81263-X).
- (en) A.R. Finlay et A.R. McCracken, Phytophthora: Symposium of the British Mycological Society, the British Society for Plant Pathology, and the Society of Irish Plant Pathologists held at Trinity College, Dublin, September 1989, Cambridge, Angleterre, Published for the British Mycological Society by Cambridge University Press, , 447 p. (ISBN 0-521-40080-5, lire en ligne), « Microbial suppression of Phytophthora cinnamoni », p. 387.
- (en) S.M. Keay et A.E. Brown, « Interactions between Psilocybe semilanceata and fungi of its habitat », Mycological Research, vol. 93, no 4, , p. 554–56 (DOI 10.1016/S0953-7562(89)80054-1).
- (en) I. Suay, F. Arenal, F.J. Asensio, A. Basilio, M.A. Cabello, M.T. Díez, J.B. García, A. González del Val, J. Gorrochategui, P. Hernández, F. Peláez et M.F. Vicente, « Screening of basidiomycetes for antimicrobial activities », Antonie van Leeuwenhoek, vol. 78, no 2, , p. 129–39 (DOI 10.1023/A:1026552024021).
- Guzmán 1983, p. 44-45
- (en) G. Guzmán, J.W. Allen et J. Gartz, « A worldwide geographical distribution of the neurotropic fungi, an analysis and discussion », Annali del Museo civico di Rovereto, vol. 14, , p. 198–280 (lire en ligne [PDF]).
- (en) Watling R., Supernatural : Meetings With the Ancient Teachers of Mankind, New York, New York, Disinformation Company, , 480 p. (ISBN 978-1-932857-84-9 et 1-932857-84-2), « Psilocybe semilanceata – a hallucinogenic mushroom native to Europe », p. 404.
- Guzmán 1983, p. 363-364
- (en) P.R. Johnston et P.K. Buchanan, « The genus Psilocybe (Agaricales) in New Zealand », New Zealand Journal of Botany, vol. 33, , p. 379–88 (DOI 10.1080/0028825X.1995.10412964, lire en ligne [PDF]).
- (en) B. Barthakur, P. Gogoi et P.K. Barua, « Agaricales of Nambar reserve forest, Golaghat, Assam, India », Advances in Plant Sciences, vol. 13, no 2, , p. 609–13.
- (en) Brande E., « Mr. E. Brande, on a poisonous species of Agaric », The Medical and Physical Journal: Containing the Earliest Information on Subjects of Medicine, Surgery, Pharmacy, Chemistry and Natural History, vol. 3, , p. 41–44 (lire en ligne).
- (en) Sowerby J., Coloured Figures of English Fungi or Mushrooms, vol. 3, Londres, J.Davis, , 248–49 p. (lire en ligne).
- Gartz 1997, p. 16
- (en) R.G. Benedict, V.E. Tyler et R. Watling, « Blueing in Conocybe, Psilocybe and a Stropharia species and the detection of psilocybin », Lloydia, vol. 30, no 2, , p. 149–157.
- (en) P.G. Mantle et E.S. Waight, « Occurrence of psilocybin in sporophores of Psilocybe semilanceata », Transactions of the British Mycological Society, vol. 53, no 2, , p. 302–304 (DOI 10.1016/s0007-1536(69)80066-5, lire en ligne).
- (de) M. Semerdžieva et F. Nerud, « Hallucinogene Pilze in der Tschechoslowakei », Česká Mykologie, vol. 27, , p. 42–47.
- (en) D.B. Repke et D.T. Leslie, « Baeocystin in Psilocybe semilanceata », Journal of Pharmaceutical Sciences, vol. 66, no 1, , p. 113–14 (DOI 10.1002/jps.2600660130).
- (en) R. Vanhaelen-Fastré et M. Vanhaelen, « Qualitative and quantitative determination of hallucinogenic components of Psilocybe mushrooms by reverse-phase high-performance liquid chromatography », Journal of Chromatography, vol. 312, , p. 467–72 (DOI 10.1016/s0021-9673(01)92800-6).
- (en) J. Jokiranta, S. Mustola, E. Ohenoja et M.M. Airaksinen, « Psilocybin in Finnish Psilocybe semilanceata », Planta Medica, vol. 50, no 3, , p. 277–78 (DOI 10.1055/s-2007-969703).
- (en) R. Heim, K. Genest, D.W. Hughes et G. Belec, « Botanical and chemical characterization of a forensic mushroom specimen of the genus Psilocybe », Journal of the Forensic Science Society, vol. 6, no 4, , p. 192–201 (DOI 10.1016/S0015-7368(66)70336-3).
- (en) R. Metzner, Sacred Mushroom of Visions : Teonanácatl : A Sourcebook on the Psilocybin Mushroom, Rochester, Vermont, Park Street Press, (ISBN 1-59477-044-1).
- Gartz 1997, p. 26
- (en) N. Anastos, S.W. Lewis, N.W. Barnett et D.N. Sims, « The determination of psilocin and psilocybin in hallucinogenic mushrooms by HPLC utilizing a dual reagent acidic potassium permanganate and tris(2,2'-bipyridyl)ruthenium(II) chemiluminescence detection system », Journal of Forensic Sciences, vol. 51, no 1, , p. 45–51 (DOI 10.1111/j.1556-4029.2005.00033.x).
- (en) N. Anastos, N.W. Barnett, S.W. Lewis, N. Gathergood, P.J. Scamells et D.N. Sims, « Determination of psilocin and psilocybin using flow injection analysis with acidic potassium permanganate and tris(2,2′-bipyridyl)ruthenium(II) chemiluminescence detection respectively », Tatlanta, vol. 67, no 2, , p. 354–59 (DOI 10.1016/j.talanta.2004.11.038).
- (en) R. Brenneisen et S. Borner, « The occurrence of tryptamine derivatives in Psilocybe semilanceata », Zeitschrift für Naturforschung C, vol. 43, nos 7–8, , p. 511–14 (ISSN 0939-5075).
- (en) J. Gartz, « New aspects of the occurrence, chemistry and cultivation of European hallucinogenic mushrooms », Annali del Museo Civico di Rovereto, vol. 8, , p. 107–23 (lire en ligne).
- (en) T. Stijve et T.W. Kuyper, « Occurrence of psilocybin in various higher fungi from several European countries », Planta Medica, vol. 51, no 5, , p. 385–87 (DOI 10.1055/s-2007-969526).
- (de) J. Gartz, « Quantitative Bestimmung der Indolderivate von Psilocybe semilanceata (Fr.) Kumm. », Biochemie und Physiologie der Pflanzen, vol. 181, no 2, , p. 117–24 (DOI 10.1016/s0015-3796(86)80079-8).
- (en) E. Ohenoja, J. Jokiranta, T. Mäkinen, A. Kaikkonen et M.M. Airaksinen, « The occurrence of psilocybin and psilocin in Finnish fungi », Journal of Natural Products, vol. 50, no 4, , p. 741–44 (DOI 10.1021/np50052a030).
- (en) M.W. Beug et J. Bigwood, « Psilocybin and psilocin levels in twenty species from seven genera of wild mushrooms in the Pacific Northwest, U.S.A. », Journal of Ethnopharmacology, vol. 5, no 3, , p. 271–85 (DOI 10.1016/0378-8741(82)90013-7).
- Stamets 1996, p. 39-41
- (en) C. Hyde, G. Glancy, P. Omerod, D. Hall et G.S. Taylor, « Abuse of indigenous psilocybin mushrooms: a new fashion and some psychiatric complications », British Journal of Psychiatry, vol. 132, , p. 602–604 (DOI 10.1192/bjp.132.6.602).
- (en) N.R. Peden, K.E.C. Macaulay, A.F. Bissett et J. Crooks, « Clinical toxicology of 'magic mushroom' ingestion », Postgraduate Medical Journal, vol. 57, no 67, , p. 543–45 (DOI 10.1136/pgmj.57.671.543).
- (en) Roland Griffiths, « Griffiths psilocybin », sur hopkinsmedicine.org, (consulté le ).
- (en) M.W. Johnson, W.A. Richards et R.R. Griffiths, « Human hallucinogen research: guidelines for safety », Journal of Psychopharmacology, vol. 22, no 6, , p. 603-620 (ISSN 0269-8811, lire en ligne, consulté le ).
- (en) K.S. Borowiak, K. Ciechanowski et P. Waloszczyk, « Psilocybin mushroom (Psilocybe semilanceata) intoxication with myocardial infarction », Journal of Toxicology: Clinical Toxicology, vol. 36, nos 1–2, , p. 47–49 (DOI 10.3109/15563659809162584).
- (en) O. Beck, A. Helander, C. Karlson-Stiber et N. Stephansson, « Presence of phenylethylamine in hallucinogenic Psilocybe mushroom: possible role in adverse reactions », Journal of Analytical Toxicology, vol. 22, no 1, , p. 45–49 (DOI 10.1093/jat/22.1.45).
- (en) H.C. Sabelli et J.I. Javaid, « Phenylethylamine modulation of affect: therapeutic and diagnostic implications », Journal of Neuropsychiatry and Clinical Neurosciences, vol. 7, no 1, , p. 6–14.
- (en) M. Franz, H. Regele, M. Kirchmair, J. Kletzmayr, G. Sunder-Plassmann et W.H. Hörl, « Magic mushrooms: hopes for a 'cheap high' resulting in end-stage renal failure », Nephrology, Dialysis, Transplantation, vol. 11, no 11, , p. 2324–27 (DOI 10.1093/oxfordjournals.ndt.a027160, lire en ligne).
- (en) H.M. Nef, H. Möllmann, P. Hilpert, N. Krause, C. Troidl, M. Weber, A. Rolf, T. Dill, C. Hamm et A. Elsässer, « Apical regional wall motion abnormalities reminiscent to Tako-Tsubo cardiomyopathy following consumption of psychoactive fungi », International Journal of Cardiology, vol. 134, no 1, , e39–e41 (DOI 10.1016/j.ijcard.2007.12.064).
- (pl) A. Adamczyk, A. Sadakierska-Chudy, J. Janoszka, A. Rymkiewicz et T. Dobosz, « Halucynogenne grzyby—lysiczki (Psilocybe). Czesc II. Identyfikacja Psilocybe semilanceata przy pomocy techniki PCR », Archiwum Medycyny Sądowej i Kryminolologii, vol. 57, no 3, , p. 285–88.
- (en) « List of psychotropic substances under international control » [PDF], sur International Narcotics Control Board, .
- (en) G. Marley, Chanterelle Dreams, Amanita Nightmares : The Love, Lore, and Mystique of Mushrooms, White River Junction, Vermont, Chelsea Green Publishing, , 263 p. (ISBN 978-1-60358-214-8 et 1-60358-214-2, lire en ligne), p. 178.
Références taxinomiques
- (en) Référence Index Fungorum : Psilocybe semilanceata (+ MycoBank)
 Portail de la mycologie
Portail de la mycologie  Portail de la pharmacie
Portail de la pharmacie