Streptococcus thermophilus
Le streptocoque thermophile (ou Streptococcus thermophilus[1],[2]) est une bactérie alimentaire, thermophile (optimum de croissance à 43 °C), présente seulement dans la fermentation du lait, où elle est notamment utilisée en association avec la bactérie Lactobacillus delbrueckii subsp. bulgaricus pour la fabrication du yaourt.
| Règne | Bacteria |
|---|---|
| Division | Firmicutes |
| Classe | Bacilli |
| Ordre | Lactobacillales |
| Famille | Streptococcaceae |
| Genre | Streptococcus |
Streptococcus salivarius subsp. thermophilus (Orla-Jensen, 1919) Farrow et Collins, 1984
Nom équivalent :
- Streptococcus salivarius thermophilus

Cette bactérie lactique appartient à la famille des Streptococcaceae, comportant deux genres phylogénétiquement très proches Streptococcus et Lactococcus. Suivant une classification récente de Moschioni et als[3] (2010), basée sur les séquences de l'ARNr 16 S, on peut distinguer cinq groupes majeurs de Streptococcus. La bactérie S. thermophilus fait partie du « groupe salivarius » qui comprend aussi Streptococcus peroris, Streptococcus salivarius et Streptococcus vestibularis, des streptocoques commensaux de la cavité buccale humaine[4].
Description
S. thermophilus se présente[5] :
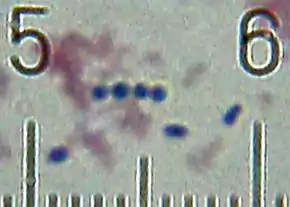
- sous forme de cocci (coque arrondie), de 0,7−1 µm, formant des chaînes ou des paires
- à coloration de Gram positive
- sa température optimale de croissance se situe entre 37 °C et 60 °C, selon les souches. Ne croît pas à 15 °C mais toutes les souches croissent à 45 °C et la plupart à 50 °C
- bactérie homofermentaire stricte (produisant du L-lactate), microaérophile
- non pathogène
- sa culture demande des vitamines B et quelques acides aminés.
Utilisation
L'intérêt économique du streptocoque thermophile est majeur, compte tenu de son utilisation dans la fabrication du yaourt et de nombreux fromages, qu'ils soient artisanaux ou industriels[6].
Il est à noter que, dans la réglementation française, le terme « yaourt » s'applique exclusivement aux préparations issues de la fermentation de laits à partir de Streptococcus thermophilus et Lactobacillus delbrueckii subsp. bulgaricus. Ainsi, un produit laitier sur lequel ne figure pas le terme « yaourt » est un lait fermenté par d'autres bactéries lactiques que celles du yaourt proprement dit.
Outre son usage pour fabriquer des yaourts, S. thermophilus intervient dans la production de nombreux fromages[7]. Il est présent dans les fromage à pâte pressée cuite à température assez élevée (45 °C) (emmental, comté, parmesan, asiago...) mais il entre aussi souvent dans les levains des fromages à pâte molle (camembert, brie...), ou des fromages frais comme la mozzarella. Cette bactérie lactique garantit une acidification rapide du lait lors de la fermentation lactique. Elle assure aussi une saveur particulière au produit en produisant du formiate, de l'acétoïne (odeur de beurre), du diacétyle (note de beurre également) et de l'acétaldéhyde (note de fraîcheur). C'est pourquoi des levains thermophiles[8] à base de S. thermophilus et Lactobacillus delbrueckii subsp. bulgaricus sont utilisés pour acidifier le lait.
Le streptocoque thermophile produit de l'acétaldéhyde à partir du lactose. Mais alors que les lactocoques peuvent réduire l'acétaldéhyde par l'alcool déshydrogénase, le streptocoque thermophile qui ne possède pas cette enzyme, en est incapable. Aussi l'acétaldéhyde est excrétée dans le produit final. En outre, le streptocoque thermophile possède une seconde voie de production de l'acétaldéhyde à partir de la thréonine[9]. Il peut en résulter une production excessive d'acétaldéhyde pouvant être responsable de défaut de flaveur.
Il a été observé une variation importante du nombre de streptocoques thermophiles au cours de la fabrication du gruyère. Cette bactérie, apportée par le levain croît très rapidement les premières heures. Toujours présente en grand nombre à la fin de l'égouttage, elle disparaît rapidement durant l'affinage. On suppose que cette disparition résulte de la lyse des cellules bactériennes. Pour certaines souches, telles que CNRZ 1358, la lyse des cellules est déclenchée par la déplétion du lactose[10]. En présence de Lactobacillus helveticus, la lyse de cette dernière induit celle de S. thermophilus durant l'affinage[11].
Origine

La proche parenté entre Streptococcus thermophilus et Streptococcus salivarius a conduit des chercheurs à émettre l'hypothèse selon laquelle S. thermophilus dériverait de S. salivarius, à partir de l'élevage des ruminants et de l'utilisation du lait de ceux-ci dans l'alimentation humaine. S. salivarius (présent dans la flore microbienne buccale normale) se serait adaptée, par mutation, à un développement dans le lait. L'Homme, au cours des âges, aurait ainsi "domestiqué" ce micro-organisme par les fermentations, avec la fabrication notamment du yaourt. La bactérie se serait adaptée à son nouveau milieu et dériverait donc de S. salivarius pour devenir S. thermophilus. Or, cette dérive, selon les systèmes de classification, aurait débuté il y plus de 9000 ans av. J.-C., ce qui corroborerait la relation avec l'apparition de l'élevage[12].
L'évolution de S. thermophilus s'est traduite par la perte de gènes ayant un pouvoir pathogène présents chez S. pneumoniae et S. pyogenes. On évalue à 10 à 11 % la proportion de gènes de S. thermophilus devenus non fonctionnels (pseudogènes) qui pourraient résulter de son adaptation à l'environnement laitier[13].
Ainsi, avec S. thermophilus, les chercheurs auraient à leur disposition un modèle d'évolution spontanée pour en étudier les mécanismes, éventuellement transposables à d'autres micro-organismes. Par ailleurs, S. thermophilus sert aussi de modèle d'étude à des chercheurs ayant identifié un mécanisme moléculaire s'apparentant, conceptuellement, à un système immunitaire bactérien à médiation moléculaire : le CRISPR[N 1]. Il s'agit d'un mécanisme de défense ciblant la multiplication des phages en empêchant indirectement l'incorporation de gènes supplémentaires.
Métabolismes des glucides

Contrairement à beaucoup d'autres bactéries Gram-positives, S. thermophilus préfère le lactose au glucose comme source d'énergie. Elle utilise la principale source de carbone du lait, le lactose, pour le convertir rapidement en lactate par fermentation et récupérer l'énergie (sous forme d'ATP), nécessaire à sa croissance.
Tout d'abord le lactose du milieu extérieur est transporté à l'intérieur de la cellule grâce à une « perméase à lactose » LacS qui lui fait franchir la membrane cellulaire sous une forme libre. Une fois à l'intérieur le lactose (un diholoside) est hydrolysé par une ß-galactosidase LacZ en glucose et galactose. La plupart des souches de S. thermophilus ne métabolisent ensuite que le glucose ; le galactose est alors excrété dans le milieu extérieur, souvent en échange de l'entrée du lactose. En fermentation laitière, le galactose s'accumule dans le lait ou le caillé et peut causer des défauts[14].
Bien que S. thermophilus soit apte à fermenter sur cinq types de sucre différents (à savoir le lactose, le saccharose, le glucose, le galactose et le fructose[15],[16]), c'est sur lactose qu'il est le plus apte à fermenter. Le saccharose et le fructose sont transportés à l'intérieur de la cellule mais avec une efficacité moindre que celle du lactose (et avec un autre type de transporteur, nommé PTS phosphotransférase). Enfin le galactose et fructose ne sont fermentés que par un nombre limité de souches alors que toutes les souches transforment le glucose en acide lactique.
Chaque molécule de glucose provenant de l'hydrolyse du lactose, est ensuite métabolisée par la voie glycolytique, en deux molécules de pyruvate, 2 ATP et 2 H2O :
En milieu anaérobie ou simplement pauvre en oxygène, une enzyme, la lactate déshydrogénase (Ldh), catalyse la transformation du pyruvate en L-lactate, tout en régénérant le NAD+ :
Plusieurs tentatives d'inactivation du gène ldh (codant Ldh) ayant conduit à des échecs, il est donc probable que cette inactivation est létale en raison de l'absence d'autre voie partant du pyruvate qui permettrait de régénérer le NAD+
S. thermophilus est donc une bactérie homolactique obligatoire produisant très majoritairement (à plus de 95 %) du L-lactate comme produit final du métabolisme des sucres.
Mais elle peut donner aussi dans des fermentations alternatives[16] de l'α-acétolactate, de l'acétoïne, de l'acétaldéhyde, du formiate et de l'acétate.
Seulement trois enzymes de dissipation du pyruvate ont été trouvées chez S. thermophilus :
La production d'exopolysaccharides
Les bactéries synthétisent plusieurs types de polysaccharides. Parmi ceux-ci, ceux qui sont excrétés à l'extérieur de la cellule, sont nommés exopolysaccharides (EPS). Ils peuvent rester attachés à la membrane bactérienne (pour les EPS capsulaires) ou être relâchés dans le milieu (pour les EPS libres). Ils sont probablement impliqués dans la protection de la cellule bactérienne contre les conditions extérieures défavorables.
Les EPS de S. thermophilus sont des polymères essentiellement de D-galactose, D-glucose et L-rhamnose mais aussi des polymères d'N-acétylgalactosamine, fucose et N-acétylglucosamine. Ils sont produits en petites quantités et de manière variable suivant les souches et les milieux de culture (source de carbone, pH, température, etc.). Dans le lait et sur 26 souches de S. thermophilus étudiées[17], seules 4 ont fabriqué plus de 100 mg/L d'EPS et 12 en ont synthétisé entre 20 et 100 mg/L.
La production d'EPS durant la fermentation lactée a la capacité d'influencer les propriétés rhéologiques (texture, viscosité, élasticité) et organoleptiques du produit fini. En général, pour le yaourt, une texture visqueuse est recherchée telle qu'elle soit capable de donner en bouche une sensation d'onctuosité au produit.
Lors de la fermentation, avec l'abaissement du pH, on observe une agrégation des protéines (de type caséines) pour former un réseau tridimensionnel, semblable au niveau microscopique, à une éponge enfermant dans ses cavités le sérum et les lipides du lait. En présence d'EPS, les interactions protéines-EPS forment des agrégats plus denses, qui retiennent mieux l'eau et diminuent la tendance à la synérèse[N 2].
Notes
- Clustered Regularly Interspaced Short Palindromic Repeats
- il s'agit de la séparation d'un liquide de son gel, en l'occurrence ici, l'expulsion du lactosérum de la matrice protéique et de la remontée de celui-ci à la surface du gel.
Références
- (en) Gerald W. Tannock, Probiotics And Prebiotics : Scientific Aspects, Horizon Scientific Press, 2005, p. 43
- Bacteria Collection : Streptococcus thermophilus, The Health Protection Agency Culture Collections
- Monica Moschioni, « Adhesion determinants of the Streptococcus species », Microbial Biotechnology, vol. 3, no 4, , p. 370–388 (ISSN 1751-7915, DOI 10.1111/j.1751-7915.2009.00138.x, lire en ligne, consulté le )
- Christine delorme, « Génomique comparative chez les Streptocoques »(Archive • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ), INRA Jouy-en-Josas
- (en) W. H. N. Holzapfel, B. J. Wood, The Genera of Lactic Acid Bacteria, Springer-Verlag New York Inc., , Softcover reprint of the original 1st ed. 1995 éd. (ISBN 978-1-4613-7666-8 et 1-4613-7666-1)
- L'UMR Génétique et Microbiologie à la une !, INRA Nancy, 2005
- (en) Patrick F. Fox, Paul L. H. McSweeney Dr, Timothy M. Cogan, Timothy P. Guinee, Cheese : Chemistry, Physics and Microbiology : General Aspects, Academic Press, , 3e éd. (ISBN 0-12-263652-X)
- (en) E. Parente, T.M. Cogan, Cheese : Chemistry, Physics and Microbiology, vol. Volume 1, Academic Press, , 123-147 p. (ISSN 1874-558X, lire en ligne), « Starter cultures: General aspects »
- André Eck, Jean-Claude Gillis (coord.), Le fromage, De la science à l'assurance-qualité, TEC&DOC, lavoisier, , 891 p. (ISBN 978-2-7430-0891-8), chap. 10 (« Les phénomènes microbiens par C. Choisy, Desmazeaud, Gueguen, Lenoir, Schmidt, Tourneur »)
- Clara Husson-Kao, « Characterization of Streptococcus thermophilus strains that undergo lysis under unfavourable environmental conditions », International Journal of Food Microbiology, vol. 55, nos 1–3, , p. 209-213 (ISSN 0168-1605, DOI 10.1016/S0168-1605(00)00166-5, lire en ligne, consulté le )
- Stéphanie-Marie Deutsch, « Lysis of lysogenic strains of Lactobacillus helveticus in Swiss cheeses and first evidence of concomitant Streptococcus thermophilus lysis », International Dairy Journal, vol. 12, no 7, , p. 591-600 (ISSN 0958-6946, DOI 10.1016/S0958-6946(02)00051-1, lire en ligne, consulté le )
- [PDF] Xavier Bellanger, Transfert, accrétion et mobilisation des éléments intégratifs conjugatifs et des îlots génomiques apparentés de Streptococcus thermophilus : un mécanisme clé de l'évolution bactérienne ?, thèse à l'université Henri-Poincaré en génomique, 2009, p. 59[60]
- Benoît Thevenard, Implication des systèmes à deux composants dans les réponses de Streptococcus thermophilus à des changements environnementaux, dont la coculture avec Lactobacillus bulgaricus, AgroParisTech, (lire en ligne)
- R Hutkins, « Galactokinase activity in Streptococcus thermophilus. », Applied and Environmental Microbiology, vol. 50, no 4, , p. 777-780 (ISSN 0099-2240, lire en ligne, consulté le )
- Vaillancourt, K., Moineau, S., Frenette, M., Lessard, C. & Vadeboncoeur, C., « Galactose and Lactose Genes from the Galactose-Positive Bacterium Streptococcus salivarius and the Phylogenetically Related Galactose-Negative Bacterium Streptococcus thermophilus: Organization, Sequence, Transcription, and Activity of the gal Gene Products. », Journal of Bacteriology, vol. 184, , p. 785-793
- Pascal Hols, « New insights in the molecular biology and physiology of Streptococcus thermophilus revealed by comparative genomics », FEMS microbiology reviews, vol. 29, no 3, , p. 435-463 (ISSN 0168-6445, DOI 10.1016/j.femsre.2005.04.008)
- Vaningelgem F., Meulen R.V.D., Zamfir M., Adriany T., Laws A.P., Vuyst L.D., « Streptococcus thermophilus ST111 produces a stable high-molecular-mass exopolysaccharide in milk-based medium, », International Dairy Journal, vol. 14, no 10, , p. 857-864
Article connexe
Liens externes
- Taxon:1308 sur MyHits
- Streptococcus thermophilus sur UniProt
- (en) Référence NCBI : taxon 1308 (taxons inclus)
- Premier séquençage d’une bactérie du yaourt : Streptococcus thermophilus - Nature Biotechnology de décembre 2004
 Portail de la microbiologie
Portail de la microbiologie