Ovogenèse
Chez les animaux l'ovogenèse, ou gamétogenèse femelle animale, est un processus biologique discontinu aboutissant à la formation d'un gamète femelle animal mature apte à donner par fécondation la cellule œuf. Chez les animaux ce gamète femelle se nomme l'ovocyte (ou ovocyte II ou ovule, sous sa forme fécondable). L'ovocyte II chez la femme, dans sa forme la plus mature, est une cellule sexuelle mesurant entre 110 et 120 µm[1] (30 µm au début de sa croissance). L'ovogenèse commence dans les ébauches embryonnaires des futurs ovaires et se termine théoriquement peu après la fécondation avec expulsion du 2nd globule polaire (dans les trompes de Fallope chez l'humain).
Elle est étroitement liée à la folliculogenèse, qui elle aboutit à la production d'un folicule possédant des fonctions endocrines par la production d'hormones sexuelles et des fonctions paracrines par la production de protéines rythmant la croissance ovocytaire (facteur de croissance ou inhibiteurs).

Chez certains animaux (poissons, amphibiens, reptiles, oiseaux), les ovocytes accumulent lors de l'ovogenèse des réserves énergétiques (vitellus) destinées au développement du futur embryon. Cette accumulation protéique peut aboutir à des ovocytes de plusieurs centimètres (ex : chez la poule, le jaune d'œuf est l'ovocyte).
Notons que l’ovogenèse n'est pas présente chez les végétaux car ces derniers produisent des gamètes femelles différentes, des oosphères.

Ovogenèse humaine
Étapes de l’ovogenèse
| Type cellulaire | Ploïdie | Chromatides | Processus | Début | Diapause | Fin |
|---|---|---|---|---|---|---|
| Cellules germinales | Diploïdes 2N | 2C (46) | Mitose | 3e/4e semaine fœtale | 8e semaine fœtale | |
| Ovogonies | Diploïdes 2N | 2C (46) | Mitose | 8e semaine fœtale | 7e mois fœtal | |
| Ovocyte I | Diploïdes 2N | 4C (92) | Méiose I | 4e à 7e mois fœtal | Prophase I (diplotène) | Pic de LH/Ovulation |
| Ovocyte II + GB I | Haploïdes 1N | 2C (46) | Méiose II | Ovulation
(après la puberté) |
Métaphase II | Fécondation
(moins de 24h après ovulation) |
| Ovule + GB II | Haploïdes 1N | 1C (23) | Méiose II | Minutes après fécondation | 1h après fécondation | |
| Zygote = Ovule + spermatozoïde | Diploïde 2N | 2C (46) | 1h après fécondation | Première division cellulaire | ||
| GB I, globule polaire I; GB II, globule polaire II; l'ovocyte II reprend sa maturation en présence du spermatozoïde dont les chromosomes ne sont pas comptabilisés. Après l'expulsion du GB II, l'ovule a transitoirement 1N, 1C. Alors commence un cycle cellulaire dans lequel les pronuclei de l'ovule et du spermatozoïde agissent de concert. | ||||||
Mise en place des premières cellules germinales chez l'embryon
Les cellules germinales (primordiales) sont complètement formées vers la 4e semaine à partir de l'épiblaste (ou ectoblaste), et migrent dans le lécithocèle (ou vésicule vitelline), plus précisément dans la paroi de l’allantoïde (partie caudale de l'embryon, dans le tissu endodermique), elles deviennent alors des gonocytes (ou gonocytes primordiaux). Les gonocytes sont les cellules précurseurs des gonades (mâle ou femelle). Certains auteurs ne font pas de différence entre cellules germinales primordiales et gonocytes primordiaux.
Vers la 5e semaine, les gonocytes vont entamer leur migration vers les ébauches des gonades appelées à ce stade « crêtes génitales », par des mouvements dits « améboïdes » (attirées par les facteurs chimiotactiques), et les coloniser pour former les futures gonades. Chez la femme, les gonocytes commencent leur évolution en ovogonies (ou gonies femelles) lors de leur migration vers les ébauches des gonades, et la finisse lorsqu'elles y arrivent (entre la 5e et la 8e semaine embryonnaire).
Phase de multiplication mitotique
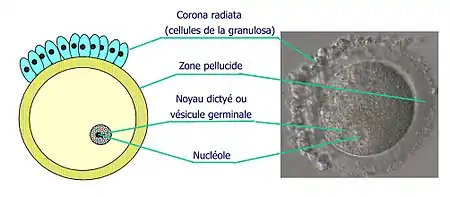
Dès la 5e semaine fœtale ces ovogonies vont faire de nombreuses mitoses successives, aboutissant à une grande augmentation de leur nombre. Leur quantité est estimée à environ 7 millions après cette multiplication, vers le 7e mois fœtal. Ce stock n'est pas renouvelable et déclinera très rapidement par atrésie (à la naissance il ne restera plus que 700 000 I et seulement 400 000 à la puberté). De plus leurs mitoses sont partielles, en effet les ovogonies issues d'une même cellule mère restent connectées par des ponts cytoplasmiques, ce qui permet la synchronisation de leur développement.
Début de la phase de maturation nucléaire méiotique (partielle, jusqu'au diplotène)
Ces ovogonies vont ensuite rentrer en méiose (début possible vers la 12e semaine) mais avec la particularité de rentrer en diapause, c'est-à-dire qu'elles sont bloquées en prophase I au stade dictyé (diplotène). Avec l'entrée en méiose la dénomination de ces cellules germinales va donc changer : les ovogonies deviennent des ovocytes primaires (ou ovocyte I).
Cet arrêt de la méiose est dû à la sécrétion de substance d'origine folliculaire dans l'ovocyte, provenant de la corona radiata :
- OMI (Ovocyte Meiotic Inhibitor)[2] (lien page en anglais)
- Haute concentration d'AMP cyclique, empêchant la synthèse de MPF (Maturation Promoting Factor)
- La présence de purines comme l'hypoxanthine et la guanosine
Cette maturation est interrompue jusqu'à l'ovulation.
Phase de croissance cytoplasmique

Cette phase de croissance ovocytaire se déroule après la naissance, elle est très longue (10 semaines) et se termine en même temps que la maturation finale du follicule (en follicule tertiaire de 10 mm). On assiste à une croissance plutôt rapide jusqu’à la formation de l’antrum chez le follicule (au moment de la formation de l’antrum, l’ovocyte a déjà atteint 80-90 % de sa taille finale), puis une croissance plutôt lente jusqu’à l’ovulation (voir folliculogénèse).

De la puberté à la ménopause, des petits groupes d'ovocytes I (et leur follicule primordial associé) vont être activés mais un seul ovocyte I va survivre et ne pas dégénérer (par atrésie folliculaire). La survie de l'ovocyte est conditionné par la survie de son follicule associé, ce sont les étapes de sélections de la folliculogenèse. De plus pendant cette phase de croissance l'ovocyte régule (avec la production de GDF9 et BMP15) la croissance folliculaire.
Pendant sa croissance l'ovocyte va voir :
- Son volume augmenté de 3 à 4 fois, passant de 30/35/40 µm de diamètre à 120 µm,
- Son nombre de mitochondries augmenté,
- Le développement de son appareil de Golgi,
- Une synthèse protéique spécifique : ZP1, ZP2 et ZP3 (associé à la zone pellucide), des protéines du cycle cellulaire (lié à la méiose), etc ….
- La formation de granules corticaux (exocytés pendant la fécondation, ils modifient les glycoprotéines de la zone pellucide qui deviennent "imperméable" aux autres spermatozoïdes),
- L'accumulation de ribosomes (1000 fois plus que dans une cellule somatique) associée à une forte transcription (avec accumulation d’ARN (300 fois plus que dans une cellule somatique) dont la plupart ne sont pas traduits),
A la fin de la phase de croissance on assiste à une réorganisation de la chromatine (en péri-nucléolaire) et un arrêt de la transcription.
Reprise de la phase de maturation nucléaire méiotique

La reprise de la maturation est déclenchée par le pic pré-ovulatoire de LH. Il se produit de nombreuses étapes de maturation juste après le pic de LH (fenêtre de prélèvement pour les FIV) et après l'ovulation (l'ovulation survient 35 à 36 heures après la décharge de LH), qui sont :
- Blocage de la transmission de OMI par une rupture des jonctions perméables qui lient la corona radiata à l'ovocyte,
- Libération de calcium intracellulaire,
- Activation de MPF et synthèse de stérols activateurs de méiose (MAS)[3] par les cellules du cumulus.
Ceci à comme conséquences pour l'ovocyte I de :
- Réaliser une maturation nucléaire :
- 15 heures après le pic il y a reprise de la méiose I, puis 20h après le pic il va finir complètement sa méiose I (aboutissent à l’expulsion du premier globule polaire)
- 35 heures après le pic il va commencer sa 2ème division méiotique (méiose II) sans la finir, et va rester bloquer en métaphase II par les facteurs cytoplasmiques CSF (kinase bloquant la dégradation des cyclines de MPF).
- Réaliser une maturation cytoplasmique :
- Migration des granules corticaux en périphérie
- Production de facteur de décondensation de la tête du spermatozoïde (glutathion)
- Arrêt de synthèse d'ARN, avec diminution de la synthèse protéiques
- Apparition de récepteurs à l'inositol triphosphate (IP3)
L'ovocyte mature (ou ovocyte II) rejoint les trompes utérines, entouré de la corona radiata (voir d'une partie du cumulus ophorius), dans sa partie 1/3 externe. Deux cas sont alors possibles :
- Si à ce moment il y a fécondation (c'est-à-dire rencontre avec le gamète mâle ou spermatozoïde) dans les 24h suivant l'ovulation (temps de survie de l'ovocyte) il y a réveil un physionomique de l'ovocyte II, sa méiose se termine aboutissant à l'expulsion du second globule polaire (GPII) et à la formation d'une cellule œuf. C'est la fin de la maturation de l'ovocyte et donc de l'ovogenèse.
- Dans le cas contraire (pas de gamète mâle rencontré, donc pas de fécondation) l'ovocyte II ne termine pas sa méiose et va être éliminé par les menstruations (phénomène cyclique), caractérisées par la désagrégation de l'endomètre.
Evolution de la réserve ovarienne/folliculaire


La réserve folliculaire (lié avec la réserve ovarienne) est composée à plus de 95 % par des follicule primordiaux. Cette réserve ne peut être régénérée, elle ne cesse de décroître.
Entre la cinquième et septième semaine du développement embryonnaire, on compte plus de 10 000 follicules. Il y a environ 7 millions de follicules dans les ovaires au bout de la 20ème semaine du développement embryonnaire, puis seuls 1 à 2 millions seront encore présents à la naissance, et le stock disponible à la puberté ne sera plus que d’environ 400 000 follicules. Parmi ces 400 000 follicules, seuls 400 vont arriver à maturité et aboutir à l'ovulation. Le reste des follicules va subir une atrésie c'est-à-dire une mort cellulaire. La maturation de ces follicules débute lors des premières menstruations.
À partir de quarante ans il y a accélération de la dégénérescence des follicules primordiaux (atrésie folliculaire), et comme tous les ovocytes sont liés à un follicule ceci entraîne en parallèle la chute du nombre d'ovocytes. Lorsque ce stock de follicules est épuisé (vers 1000 follicule restant), on parle de ménopause. Cette atrésie folliculaire est régulée par le facteur pro-apoptotique Bax et le facteur anti-apoptotique Bcl2. L'inactivation de Bax entraîne la multiplication par 3 du nombre de follicules primordiaux, tandis que l'inactivation de Bcl2 entraîne leur diminution.
Pour calculer la réserve ovarienne, une mesure du taux de AMH est réalisée. L'AMH est corrélée au nombre de follicules antraux et est donc un bon marqueur du pool folliculaire.
Mais cette description n'est pas tout à fait juste, puisque que des chercheurs ont mis au jour des cellules souches ovariennes d'abord chez la souris en 2004, puis chez la femme en 2012, ouvrant ici le champ à des traitements contre l’infertilité pathologique et physiologique féminine[4],[5].
Ovogenèse générale
L'ovocyte est rempli de substance de réserve (vitellus) pour permettre le développement embryonnaire. À l'exception des mammifères, il n'y a en général pas d'apport nutritif durant le développement embryonnaire (exception : certains requins, certains amphibiens (Salamandra atra ou salamandre noire), certains reptiles).
Maturation ovocytaire
Au cours de la vie intra-utérine, l'ovogonie subit une « pré-méiose » : elle s'arrête au stade diplotène (mais exactement dictyotène) de la prophase de la méiose I, puis elle subit un petit accroissement (son volume cellulaire augmente). On parle alors d'ovocyte I. À la puberté, cet ovocyte I entrera dans une phase de grand accroissement puis dans une phase d'accroissement rapide, et enfin une phase de maturation.
La maturation se fait grâce à la méiose (réductionnelle). Cependant, elle donnera deux cellules de taille différente :
- un ovocyte II qui conservera la majeure partie du cytoplasme ;
- un premier globule polaire de taille réduite.
L'ovulation a lieu quelques heures après cette maturation. L'ovocyte II a déjà engagé sa deuxième division de méiose (équationnelle), mais sera bloqué en métaphase II. Si l'ovocyte II est fécondé par un spermatozoïde il termine sa division et donne naissance à un ovotide et un deuxième globule polaire (GPII).
On considère l'ovotide comme le véritable gamète femelle (haploïde), mais la fécondation entraînant la fusion des deux gamètes, cette cellule est en réalité diploïde. C'est la première cellule du futur individu, qu'on appelle zygote.
Notes et références
- Serge Nef, « Quelle est la taille d'un ovule et d'un spermatozoïde en micromètre et millimètre ? », rts.ch, (lire en ligne, consulté le )
- (en) « Oocyte maturation inhibitor », Wikipedia, (lire en ligne, consulté le )
- Charles Thibault et Marie-Claire Levasseur, La reproduction chez les mammifères et l'homme, Editions Quae, , 928 p. (ISBN 978-2-7298-0417-6, lire en ligne)
- (en) Yvonne A R White, Dori C Woods, Yasushi Takai et Osamu Ishihara, « Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women », Nature Medicine, vol. 18, no 3, , p. 413–421 (ISSN 1546-170X, DOI 10.1038/nm.2669, lire en ligne, consulté le )
- nature video, « Stem cells put women on fertile ground - by Nature Video », (consulté le )
Articles connexes
Liens externes
- Développement des cellules germinales dans l'ovaire sur un site développé par les Universités de Fribourg, Lausanne et Berne
- Définition d'ovogonie sur le site d'Aquaportail
- Présentation sur la croissance et maturation ovocytaire sur le site des Facultés de médecine de Toulouse
- Présentation sur l'ovogenèse et la folliculogénèse sur le site des Facultés de médecine de Toulouse
 Portail de la biologie
Portail de la biologie  Portail de la médecine
Portail de la médecine