Séquence régulatrice
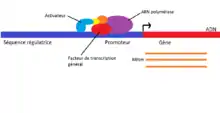
Les séquences régulatrices, appelées aussi séquence-cis, sont une partie de l’ADN non codant (séquences du génome qui ne sont pas traduites en protéines) et qui influent sur le niveau de transcription des gènes. Elles sont reconnues par des facteurs de transcription, appelés facteur-trans, qui agissent de différentes façons, en augmentant ou en diminuant l’expression du gène. Les séquences régulatrices interviennent ainsi au niveau de l’initiation de la transcription dans la régulation de l'expression des gènes. Les promoteurs sont tout d’abord identifiés comme les éléments essentiels à la transcription, étant le lieu d’attache de l’ARN polymérase et de facteurs généraux de la transcription nécessaires à l’initiation. Des régions modulant la transcription ont ensuite été identifiées comme des amplificateurs chez les eucaryotes. Ces derniers sont pour la plupart situés en amont du promoteur, lui-même situé en amont de la séquence codante. Ils sont même parfois placés à une grande distance, atteignant plus de 10 kb du site d’initiation de la transcription[1]. Les facteurs de transcription peuvent ainsi induire des repliements de l’ADN pour assurer un contact avec le complexe initiateur.
La plupart des gènes eucaryotes sont sous le contrôle de multiples éléments régulateurs. Un même élément peut agir sur plusieurs gènes et un gène donné peut être sous le contrôle d’une combinaison d’éléments. Différentes combinaisons de facteurs assurent le contrôle et la coordination de l’ensemble des gènes.
Les séquences régulatrices pourraient représenter environ 17 % du génome humain alors que les régions codantes des gènes (exons) pourraient en représenter 2,9 % selon une estimation du projet ENCODE dont l'objectif est de déterminer les fonctions de toutes les parties du génome humain. Les séquences régulatrices étaient autrefois considérées comme faisant partie de l’ADN non codant puisque leurs fonctions étaient mal comprises.
Caractérisation des séquences régulatrices
Il est possible de fusionner l’ADN des séquences régulatrices à l’ADN codant grâce à un gène rapporteur. On mesure alors l’expression du gène, généralement via l’activité de la protéine produite. Il est possible aussi de remplacer des morceaux d'ADN (linker scanning mutations) dans un promoteur, ce qui permet la caractérisation des éléments impliqués dans la régulation. La mutation de nucléotides spécifiques permet de caractériser précisément les motifs impliqués.
Procaryote
Chez les procaryotes, les principales séquences régulatrices des opérons sont les promoteurs et les opérateurs.
À partir de l'analyse de la séquence de différentes régions promotrices, des séquences consensus ont été établies, dont il existe plusieurs sortes. L'un des promoteurs bactériens les plus connus est formé de deux séquences conservées de 6 nucléotides, séparées par 17-19 nucléotides non conservés. Elles sont appelées boîtes -10 (séquence nucléotidique : TATAAT) et -35 (séquence nucléotidique : TTGACA), les chiffres représentant le nombre de nucléotides en amont du site de démarrage de la transcription. En général, plus la séquence s’approche du consensus, plus le promoteur est fort. Le nombre de transcrits générés à partir de ce promoteur dans un laps de temps donné sera plus élevé puisqu’il a une meilleure affinité pour le facteur sigma. C’est, en fait, le facteur sigma qui lie de façon spécifique le promoteur du gène et attire la polymérase d'ARN au début du gène. Les séquences du promoteur orientent le facteur et déterminent ainsi l’orientation de la polymérase sur l’ADN.
Des répresseurs ou des activateurs qui viennent se lier aux séquences régulatrices, généralement des opérateurs, peuvent aussi agir pour contrôler l’expression génique. Les répresseurs empêchent la liaison de l’ARN polymérase au promoteur ou bien l’immobilisent au site de sorte qu’elle ne puisse commencer la transcription. Les activateurs augmentent l’affinité de la polymérase pour le promoteur et peuvent agir en allostérie.
Un cas de figure bien connue de régulation des gènes chez les procaryotes est l’opéron lactose chez E. coli.
Eucaryote
Chez les eucaryotes, des facteurs de transcription généraux, collectivement appelés TFII, permettent l’association de l’ARN polymérase au promoteur et l’initiation de la transcription. Un complexe médiateur forme aussi un lien entre la polymérase et les facteurs de transcription liés aux séquences régulatrices.
Le promoteur des eucaryotes est généralement composé de 2 ou 3 séquences conservées séparées par des nucléotides non conservés. Les facteurs de transcription généraux reconnaissent ces séquences et jouent collectivement le même rôle que le facteur sigma des bactéries. La formation du complexe de pré-initiation survient à la boîte TATA lorsqu'elle est présente. Il s’agit d’une séquence conservée à -25/-35 bases en amont du site d’amorçage et détermine le site d’initiation de la transcription.
Les sites amplificateurs ont tout d’abord été identifiés par Walter Schaffner en 1981, qui étudiait le promoteur du virus SV40. On découvrit que ces séquences stimulaient des promoteurs autres que ceux de SV40 et que leur effet ne dépendait ni de leur orientation, ni de la distance les séparant du site d’initiation de la transcription[1].
Répresseurs
Les répresseurs peuvent agir de plusieurs façons. Ils peuvent interférer avec les activateurs, soit en les liant directement, soit en liant leurs sites de liaison. Ils peuvent interagir avec le complexe d’initiation en empêchant la polymérase ou un des facteurs généraux de la transcription d’accéder au promoteur. Ils peuvent aussi maintenir la polymérase au promoteur à l’aide d’un domaine de liaison protéine-protéine. Ils peuvent compacter la chromatine et rendre les gènes inaccessibles en recrutant des histones méthyltransférase et des histones désacétylase.

Activateurs
Ils aident au recrutement de la polymérase à un promoteur faible (liaison coopérative) ou stimulent la transcription en provoquant un changement conformationnel dans l’ARN polymérase ou l’ADN du promoteur (allostérie).
Les activateurs sont typiquement modulaires, composés d’un domaine liant l’ADN et d’un domaine d’activation. Les activateurs de la transcription agissent rarement directement sur l’ARN polymérase. Ils peuvent recruter indirectement la polymérase en complexe avec le médiateur ou des facteurs généraux de la transcription. Ils peuvent recruter des facteurs modificateurs des histones tels que des histones acétyltransférase et des complexes remodeleurs, qui altèrent la chromatine pour permettre l’accès à des sites de liaison spécifiques sur l’ADN.
Certaines protéines appelées isolateurs se lient à l’ADN sur des régions situées entre un promoteur et son activateur. Ces protéines empêchent l’activateur d’agir sur un promoteur, mais sans directement inhiber l’un ou l’autre.
Gal4, un facteur de transcription chez la levure, se lie à la séquence SAAG en présence de galactose et active la transcription de GAL1, gène nécessaire au métabolisme du galactose, jusqu’à 1000 fois le taux basal.
Voies de signalisation
Plusieurs voies de signalisation sont vouées en grande partie à la régulation de facteurs de transcription. Une cellule de mammifère typique peut exprimer des récepteurs pour plus d’une centaine de molécules dont la fonction première est de réguler l’activité de facteurs de transcription. Plusieurs de ces voies de signalisation sont régulées par des récepteurs à activité kinase tels que les récepteurs TGFβ et les récepteurs tyrosine kinase (RTK).
Les hormones liposolubles peuvent influencer directement l’activité d’une famille de facteurs de transcription (récepteurs nucléaires) qui produiront des réponses géniques par leur liaison aux séquences régulatrices.
Structure de la chromatine
Les gènes activement transcrits se situent dans les zones décondensées de la chromatine puisque ces régions sont plus accessibles aux facteurs de transcription. En effet, des facteurs du remodelage du nucléosome peuvent démembrer les nucléosomes au niveau de séquences régulatrices, permettant aux facteurs de la transcription d’atteindre le brin d’ADN. Les activateurs empêchent alors le blocage par les histones et permettent le recrutement de l’ARN polymérase au promoteur ainsi que ses facteurs associés. Les régions du promoteur et de l’amplificateur en cours de transcription comportent alors moins de nucléosomes, ce qui les rend plus sensibles à l’ADNase que le reste du gène. De fait, les séquences régulatrices des gènes activement transcrits sont dites sites hypersensibles à l’ADNase. L’ARN polymérase peut alors commencer la transcription du gène tout en désassemblant les nucléosomes. Le pouvoir de désassemblage de la polymérase est possible par l’acétylation des histones et leurs liaisons aux protéines non histones HMG-14 et HMG-17. Ces dernières restreignant la liaison de l’histone H1 responsable de la condensation de la chromatine.
Méthylation de l’ADN
Certains gènes sont maintenus silencieux par méthylation de l’ADN de leur région promotrice. Les séquences méthylées sont reconnues par des protéines liant l’ADN méthylé telles que MeCP2. Cette protéine recrute à son tour des histones désacétylases et des histones méthylases qui modifient la chromatine et rendent la région impossible à transcrire.
La méthylation de l'ADN est l'acteur majeur de la mise en place de l'empreinte parentale, mécanisme par lequel l'expression d'un gène va dépendre de l'origine parentale. L'une des deux copies du gène est alors gardée silencieuse par les groupements méthyles. Un certain nombre de gènes soumis à empreinte parentale est impliqué dans le développement et la croissance de l’embryon et du placenta[2]. D'autres sont impliqués dans le développement post-natal, avec des rôles affectant l'allaitement et le métabolisme[3].
Développement
Les facteurs de transcriptions, en se liant à un amplificateur, permettent de diriger l’expression génique au cours du développement embryonnaire et de la différenciation cellulaire. Les cellules eucaryotes possèdent en effet un grand nombre de facteurs de transcription étant donné que les gènes des organismes pluricellulaires s’expriment de manière élaborée. Leur expression étant distincte à chaque tissu et à chaque étape du cycle cellulaire. C’est notamment le cas des morphogènes qui agissent sur les séquences régulatrices des gènes afin d’induire des réponses cellulaires, ce qui permet de former des axes de polarité dans l’embryon.
Évolution
La majorité de l’ADN intergénique et des portions non codantes des gènes (introns) n’interviennent pas dans la survie et la capacité de reproduction. La conservation de leurs séquences n’est donc pas soumise à la pression sélective. En effet, la sélection naturelle a tendance à éliminer les individus dont le génome porte des mutations dans les séquences codantes de l’ADN ainsi que dans les séquences régulatrices des gènes. Ces éléments ont alors disposition à être conservés. En comparant le génome d’espèces différentes, tel l’humain et la souris, il est possible de déterminer quelles sont ces séquences et déterminer certains éléments fonctionnels du génome.
En étudiant les différences dans les séquences d’ADN et dans l’organisation des gènes, il est aussi possible de déterminer l’origine génétique de certains caractères. En ce qui concerne les séquences régulatrices, leurs modifications peuvent augmenter ou diminuer l’affinité de facteurs de transcription pour leurs sites. Elles peuvent aussi permettre la liaison à de nouveaux facteurs de transcription ou restreindre l’accès aux anciens. Ceci pouvant modifier de façon importante la réponse et le développement de l’organisme.
Les génomes du chimpanzé et de l’homme ont été comparés et il a été estimé que les deux génomes différaient de 4 %, soit des dizaines de millions de différences[4]. Une attention particulière a notamment été portée sur le gène FOXP2, codant un facteur de transcription. La protéine humaine possède en fait deux acides aminées différents de celle du chimpanzé. De plus, selon les calculs, les modifications de ce gène eurent lieu au cours des 120 000 à 200 000 dernières années, époque où semble apparaître l’espèce humaine. Il est alors intéressant de constater que les personnes ayant une mutation sur le gène FOXP2, souffrent de problèmes de la parole et du langage. Ces individus sont notamment incapables d’assurer le mouvement délicat des lèvres et de la langue nécessaire à une communication vocale. Les modifications du gène FOXP2 peuvent donc avoir eu un rôle important dans l’évolution humaine pour l’établissement du langage et de la communication verbale[4].
Notes et références
- (en) Banerji, J., Rusconi, S. et Schaffner, W., « Expression of a β-globin gene is enhanced by remote SV40 DNA sequences », Cell, , p. 299-308 (DOI 10.1016/0092-8674(81)90413-X, lire en ligne).
- (en) Isles, A. R. et Holland, A. J., « Imprinted genes and mother–offspring interactions », Early Human Development, , p. 73-77 (DOI 10.1016/j.earlhumdev.2004.10.006, lire en ligne).
- (en) Tycko, B. et Morison, I. M., « Physiological functions of imprinted genes », Journal of Cellular Physiology, , p. 245-258 (DOI 10.1002/jcp.10129, lire en ligne).
- Gerald Karp (trad. de l'anglais), Biologie cellulaire et moléculaire : Concepts and experiments, Bruxelles/Paris, De Boeck supérieur, , 844 p. (ISBN 978-2-8041-6011-1, lire en ligne).
Voir aussi
Articles connexes
Bibliographie
- (en) B. Alberts, A. Johnson, J. Lewis, J, M. Raff, K. Roberts et P. Walter, Molecular Biology of the Cell, Garland Science, , 4e éd. (présentation en ligne)
- G. M. Cooper, La cellule : Une approche moléculaire, De Boeck supérieur, (lire en ligne)
- E. T. Dermitzakis et A. G. Clark, « Evolution of Transcription Factor Binding Sites in Mammalian Gene Regulatory Regions: Conservation and Turnover », Molecular Biology and Evolution, vol. 19, no 7, , p. 1114‑1121 (lire en ligne)
- G. Karp, Biologie cellulaire et moléculaire : Concepts and experiments, De Boeck supérieur, (lire en ligne)
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire