Phage lambda
Le phage lambda (Enterobacteria phage λ) est un virus bactériophage qui infecte la bactérie Escherichia coli. Ce bactériophage est un virus à ADN double brin, empaqueté dans une capside icosaédrique prolongée d'une queue et de fibrilles permettant l'ancrage sur la bactérie. Le phage lambda est sujet à deux cycles d'évolution possible : le cycle lytique qui conduit à la réplication rapide du virus et à la mort de la cellule hôte, en l'occurrence Escherichia coli, et le cycle lysogénique durant lequel il insère son génome de manière dormante dans celui de la bactérie et subit des réplications en même temps que le reste du génome bactérien. Il peut ainsi se propager à la descendance de la cellule infectée. Ce cycle aboutit généralement à un cycle lytique et à l'excision du génome phagique hors du génome bactérien. L'existence de la voie lysogénique fait de lambda un phage « tempéré ». L'intégration du génome du virus dans le chromosome bactérien se fait via un mécanisme de recombinaison spécifique, sur un site bien défini entre les opérons gal (utilisation du galactose) et bio (synthèse de la biotine).

| Type | Virus |
|---|---|
| Groupe | Groupe I |
| Ordre | Caudovirales |
| Famille | Siphoviridae |
| Genre | λ-like viruses 7 |
— auteur incomplet —, date à préciser
La biologie moléculaire et la biochimie de ce virus sont particulièrement bien connues, ce qui fait qu'il a été largement utilisé comme vecteur de clonage de l'ADN, ainsi que pour permettre l'insertion de séquences d'ADN spécifiques dans le génome dE. coli, via le mécanisme d'intégration lysogénique du virus.
Description
Généralités
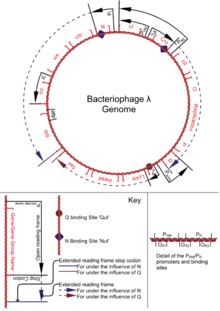
Le phage lambda est une particule virale possédant une tête, qui contient l'ADN du phage, ainsi qu'une queue terminée par des fibrilles. La tête icosaédrique contient de l'ADN linéaire double brin long de 48 502 paires de bases[1], flanquée de part et d'autre de séquences répétées, simple brin de 12 paires de bases. Ces séquences cohésives sont appelées séquences « cos ». Le phage lambda possède un spectre d'hôtes très restreint, principalement limité à Escherichia coli et des espèces proches. Le phage lambda est découvert par Esther Lederberg en 1950 [2], sa recombinaison génétique est ensuite étudiée par Grete Kellenberger-Gujer dès 1961. Il est devenu un organisme modèle et outil très utilisé en biologie moléculaire.
Cycle de vie
Le phage est capable de s'attacher à la surface de son hôte, Escherichia coli. Il injecte alors son ADN dans le cytoplasme de la bactérie. L'ADN phagique est linéaire et sous forme double brin dans la tête du phage. Il se circularise dans la cellule grâce à des séquences répétée « cos ». Le phage circularisé peut alors se répliquer dans la cellule par réplication en thêta, puis en cercle roulant. La réplication en cercle roulant permet la formation d'un concaténaire. Simultanément à cette étape de réplication, le phage produit les protéines de sa capside. Une fois le concaténaire et la tête de la capside obtenus, il y a une interaction spécifique entre un site « cos » situé sur le concaténaire et les protéines de la tête. L'ADN entre dans la capside jusqu'au site « cos » suivant, au niveau duquel il est clivé par les protéines de la tête. Une fois la tête du phage pleine, les protéines de la queue s'assemblent. Les particules virales sont libérées et peuvent infecter d'autres cellules.
Mais une fois sous sa forme circulaire dans la cellule, le phage lambda peut aussi s'intégrer dans le génome de l'hôte, et rester sous forme de prophage. Il réalise alors un cycle lysogénique. Cette intégration dans le génome de l'hôte se réalise par recombinaison site-spécifique, entre un site dit « attP » sur l'ADN phagique et le site « attB » situé sur le chromosome de la bactérie. Le site attB est unique dans le génome d' E. Coli, et est situé entre les gènes gal (gène codant la galactosidase, enzyme de dégradation du galactose) et bio (gène codant la voie de biosynthèse de la biotine). L'intégration dans le chromosome nécessite l'intervention d'une intégrase, codée par le phage mais aussi des protéines de l'hôte, comme la protéine IHF (integration host factor, facteur capable de modifier la topologie de l'ADN). Cette étape permet la création de deux nouveaux sites de part et d'autre du phage appelés attR et attL. Le prophage, sous certaines conditions pourra s'exciser et partir en cycle lytique. Cette excision nécessite l'intégrase produite par le phage, les protéines IHF de l'hôte mais également la protéine XIS (excisionase) codée par le phage et qui permet la reconnaissance des sites attR et attL par l'intégrase. L'excision reconstitue les sites attB et attP.
Cycle lytique ou lysogénique?
Après l'injection de l'ADN phagique dans le cytoplasme, il y a « prise de décision » entre l'entrée en cycle lytique ou la croissance lysogénique, l'intégration dans le génome bactérien sous forme de prophage. Les facteurs influençant cette prise de décision sont mal connus, mais il semble que les conditions physiologique de l'hôte participent à cette étape[3].
Si la prise de décision provoque l'entrée en cycle lysogénique, le phage met en place des systèmes de régulation permettant de se maintenir sous forme de prophage. Pour cela le prophage produit un répresseur Ci qui inhibe son excision du génome hôte en bloquant la transcription de l'intégrase et de la protéine XIS. Sous certaines conditions comme lorsque la survie de la cellule n'est plus assurée, il est cependant nécessaire pour le phage de pouvoir s'exciser et de réaliser un cycle lytique[4]. Ainsi, le represseur Ci est capable d'autoclivage (provoquant son inactivation) lorsque la protéine RecA de la bactérie est sous sa forme activée. Dans la cellule, la protéine RecA a un rôle de signalisation des dommages au niveau de l'ADN. Cette protéine est capable de provoquer l'induction du système SOS de réparation de l'ADN cellulaire[5]. En cas de conditions où la survie de l'hôte est compromise le phage lambda sera donc capable de s'exciser et de produire des particules virales.
La transduction restreinte par Lambda
La transduction est l'apport de matériel génétique dans une cellule en utilisant un phage comme vecteur. Le phage Lambda réalise une transduction restreinte, c'est-à-dire qu'il ne peut transférer qu'un nombre réduit de gènes, soit le gène gal, soit le gène bio. La fréquence de transfert est faible (environ 1 phage transducteur pour 10^12 phages)
Comment former des phages transducteurs?
Une recombinaison illégitime permet une excision du phage ainsi qu'un morceau d'ADN. Cet ADN est ensuite encapsidé et infectera une nouvelle cellule. Cependant, la taille de l'ADN emporté par le phage est réduite par le phénomène dite de « tête pleine », qui consiste en une limite physique de taille d'ADN que peut contenir la tête de la capside. Ainsi, le phage lambda peut emporter soit le gène gal, soit le gène bio.
Devenir de cet ADN bactérien
Le phage transducteur est capable d'infecter une nouvelle cellule, appartenant à son spectre d'hôte (principalement E. coli). Le phage transducteur ayant été excisé par recombinaison illégitime porte le site attR ou attL et non pas le site attP qui permet son intégration dans le chromosome par recombinaison site-spécifique. L'intégration du phage transducteur se réalise par recombinaison homologue entre le gène gal ou bio porté par le phage et les gènes correspondant sur le chromosome bactérien. Cette recombinaison ne permet donc pas l'intégration de nouveaux caractères dans la cellule transduite, mais un mélange d'allèles.
Lectures et documents complémentaires
- James Watson, Tania Baker, Stephen Bell, Alexander Gann, Michael Levine, Richard Losick " Molecular Biology of the Gene (International Edition)" - 6e Edition
- Mark Ptashne and Nancy Hopkins, "The Operators Controlled by the Lambda Phage Repressor", PNAS, v.60, n.4,pp. 1282–1287 (1968).
- Barbara J. Meyer, Dennis G. Kleid, and Mark Ptashne, "Lambda Repressor Turns Off Transcription of Its Own Gene", PNAS, v.72, n.12, pp. 4785–4789 ().
- Brussow, H. and Hendrix, R.W. 2002 "Small is beautiful.", Cell, 108, 13-16
- Dodd, J.B. Shearwin, K.E. and Egan, J.B. 2005 "Revisited gene regulation in phage lambda.", Curr Opin GenetDev, 15, 145-152
- Mc Grath S and van Sinderen D, ed (2007). Bacteriophage: Genetics and Molecular Biology (http:// books. google. com/books?id=35sAile2OnYC& pg=PT266 ) (1st ed.). Caister Academic Press. pp. 266 ff.. (ISBN 1-904455-14-X). .
Notes et références
- F. Sanger, A.R. Coulson, G.F. Hong, D.F. Hill et G.B. Petersen, « Nucleotide sequence of bacteriophage lambda DNA », J. Mol. Biol., vol. 162, , p. 729-773 (PMID 6221115)
- Esther Lederberg, Lysogenicity in Escherichia coli strain K-12 , Microbial Genetics Bulletin, v.1, pp. 5-8 (January 1950); suivi de Lederberg, EM; Lederberg, J (1953). "Genetic Studies of Lysogenicity in Escherichia coli." . Genetics 38 (1): 51–64. . PMC 1209586.
- St-Pierre F, Endy D (2008). "Determination of cell fate selection during phage lambda infection". Proc. Natl. Acad. Sci. U.S.A. 105
- James Watson, Tania Baker, Stephen Bell, Alexander Gann, Michael Levine, Richard Losick (trad. Michel Jacquet et Francis Le Gaillard), Biologie moléculaire du gène, Pearson Education France, 2009, 6e éd. (1re éd. 1965), 688 p. (ISBN 978-2-7440-7348-9)
- "Barbara J. Meyer (), HHMI Interactive (52): 20705–10. doi:10.1073/pnas.0808831105.. PMC 2605630.]
 Portail de la virologie
Portail de la virologie  Portail de la microbiologie
Portail de la microbiologie