Méthanogenèse
La méthanogenèse est un ensemble de voies métaboliques produisant du méthane chez certains microorganismes, qualifiés de méthanogènes. De tels microorganismes n'ont été identifiés qu'au sein du domaine des archées, un groupe de procaryotes distincts des bactéries et des eucaryotes d'un point de vue phylogénétique, bien que de nombreuses archées vivent étroitement associées à des bactéries anaérobies. La production de méthane est une manifestation importante et très répandue du métabolisme microbien. C'est bien souvent la dernière étape de la décomposition de la biomasse, participant ainsi au processus plus général de méthanisation.
Certaines études réalisées au début du 21e siècle indiqueraient que les feuilles des plantes vivantes produiraient elles aussi du méthane[1], tandis que d'autres recherches établiraient plutôt que les plantes rejetteraient par les feuilles du méthane absorbé au niveau des racines[2]. Il se peut que des processus inconnus produisent du méthane chez les végétaux, mais ceci n'est en aucun cas attesté.
Biochimie de la méthanogenèse
La méthanogenèse microbienne est une forme de respiration anaérobie[3], l'oxygène étant un inhibiteur de croissance chez les méthanogènes. Dans la respiration aérobie, O2 est l'accepteur final d'électrons et il se forme de l'eau ; dans le cas des méthanogènes, l'accepteur final d'électrons est le carbone de petites molécules organiques.
On peut diviser les organismes méthanogènes en trois classes en fonction de leur substrat : les hydrogénotrophes, les méthylotrophes et les acétotrophes[4].
| Réactions globales pour la méthanogenèse | ΔG°’ [kJ/mol CH4][4] | Organisme |
| Hydrogénotrophe | ||
| CO2 + 4 H2 → CH4 + 2 H2O | −135 | la plupart des méthanogènes |
| 4 HCOOH → CH4 + 3 CO2 + 2 H2O | −130 | beaucoup de méthanogènes hydrogénothrophes |
| CO2 + 4 CH3CHOHCH3 → CH4 + 4 CH3COCH3 + 2 H2O | −37 | quelques méthanogènes hydrogénothrophes |
| 4 CO + 2 H2O → CH4 + 3 CO2 | −196 | Methanothermobacter et Methanosarcina |
| Méthylotrophe | ||
| 4 CH3OH → 3 CH4 + CO2 + 2 H2O | −105 | Methanosarcina et autres méthanogènes méthylothrophes |
| CH3OH + H2 → CH4 + H2O | −113 | Methanimicrococcus blatticola et Methanosphaera |
| 2 (CH3)2S + 2 H2O → 3 CH4 + CO2 + 2 H2S | −49 | quelques méthanogènes méthylothrophes |
| 4 CH3NH2 + 2 H2O → 3 CH4 + CO2 + 4 NH3 | −75 | quelques méthanogènes méthylothrophes |
| 2 (CH3)2NH + 2 H2O → 3 CH4 + CO2 + 2 NH3 | −73 | quelques méthanogènes méthylothrophes |
| 4 (CH3)3N + 6 H2O → 9 CH4 + 3 CO2 + 4 NH3 | −74 | quelques méthanogènes méthylothrophes |
| 4 CH3NH3Cl + 2 H2O → 3 CH4 + CO2 + 4 NH4Cl | −74 | quelques méthanogènes méthylothrophes |
| Acétotrophe | ||
| CH3COOH → CH4 + CO2 | −33 | Methanosarcina et Methanosaeta |
Réduction du dioxyde de carbone
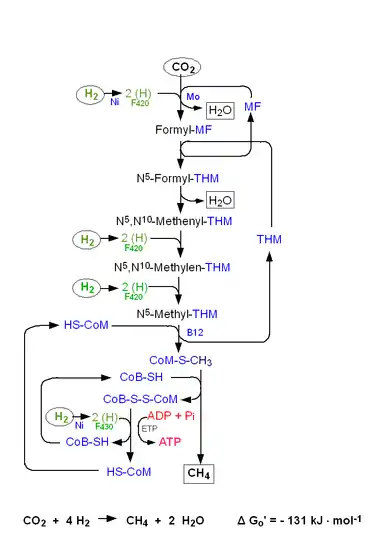
La réduction d'une molécule de dioxyde de carbone en méthane s'écrit de manière globale :
Cette voie de synthèse fait intervenir successivement plusieurs enzymes, coenzymes et cofacteurs en sept étapes majeures en utilisant le dihydrogène comme donneur d'électrons principal, la plupart des méthanogènes hydrogénotrophes pouvant également utiliser le formate comme donneur d'électrons[4].
- La formylméthanofurane déshydrogénase (EC ) catalyse la fixation d'une molécule de dioxyde de carbone CO2 sur son cofacteur méthanofurane pour donner le formylméthanofurane. Cette enzyme recourt également au cofacteur à molybdène, contenant du molybdène et de la molybdoptérine. La source d'électrons est le dihydrogène entrainant la réduction de la coenzyme F420.
- CO2 + méthanofurane + accepteur d'électrons réduit H2O + formylméthanofurane + accepteur d'électrons oxydé.

- La formylméthanofurane:tétrahydrométhanoptérine N-formyltransférase (EC ) transfère le groupe formyle sur l'azote no 5 de sa coenzyme, la tétrahydrométhanoptérine, pour former la formyltétrahydrométhanoptérine :
- formylméthanofurane + 5,6,7,8-tétrahydrométhanoptérine méthanofurane + 5-formyl-5,6,7,8-tétrahydrométhanoptérine.
- Le groupe formyle se condense alors au sein de la molécule pour donner la méthényl-THM+.
- La méthényl-THM+ est ensuite réduite en méthylène-THMPT[5] par une méthylènetétrahydrométhanoptérine déshydrogénase (EC ) avec la coenzyme F420 comme source d'électrons :
- 5,10-méthényl-5,6,7,8-tétrahydrométhanoptérine + coenzyme F420 réduite 5,10-méthylène-5,6,7,8-tétrahydrométhanoptérine + coenzyme F420 oxydée.
- La méthylène-THM est ensuite convertie en méthyl-THM par une méthylènetétrahydrométhanoptérine réductase (EC ) avec la coenzyme F420 comme source d'électrons :
- 5,10-méthylènetétrahydrométhanoptérine + coenzyme F420 réduite 5-méthyl-5,6,7,8-tétrahydrométhanoptérine + coenzyme F420 oxydée.
- La méthyl-THM transfère son méthyle à la coenzyme M sous l'action de la tétrahydrométhanoptérine S-méthyltransférase[3] (EC ) pour régénérant la tétrahydrométhanoptérine en formant la méthyl-coenzyme M :
- 5-méthyl-5,6,7,8-tétrahydrométhanoptérine + coenzyme M 5,6,7,8-tétrahydrométhanoptérine + méthyl-coenzyme M.
- La méthyl-coenzyme M réagit enfin avec la coenzyme B pour donner un hétérodisulfure en libérant le méthane sous l'action de la méthyl-coenzyme M réductase (EC ) :
Le composé disulfure est ensuite réduit à l'aide de la coenzyme F420 pour régénerer les deux thiols.
Réduction de l'acide acétique

La réduction d'une molécule d'acide acétique en méthane s'écrit de manière globale :
Cette voie de synthèse fait intervenir successivement plusieurs enzymes, coenzymes et cofacteurs en quatre étapes majeures[4].
- Un groupe acétyle est transféré sur la coenzyme A à partir d'une molécule d'acétate pour former l'acétyl-coenzyme A. Ceci peut se faire de deux manières[6] :
- - à l'aide du système acétate kinase (EC )- phosphate acétyltransférase (EC )
- EC : acétate + ATP phosphate d'acétyle + ADP
- EC : phosphate d'acétyle + coenzyme A acétyl-CoA + phosphate
- - en utilisant l'acétyl-coenzyme A synthétase (EC )
- acétate + ATP + coenzyme A acétyl-CoA + AMP + diphosphate
- L'acétyl-coenzyme A synthase (EC ) transfère un groupe méthyle de l'acétyl-coenzyme A sur la tétrahydrométhanoptérine pour donner la méthyl-THM en libérant une molécule de monoxyde de carbone qui est oxydée en dioxyde de carbone.
- La méthyl-THM transfère son méthyle à la coenzyme M sous l'action de la tétrahydrométhanoptérine S-méthyltransférase[3] (EC ) pour régénérer la tétrahydrométhanoptérine en formant la méthyl-coenzyme M.
- La méthyl-coenzyme M réagit enfin avec la coenzyme B pour donner un hétérodisulfure en libérant le méthane sous l'action de la méthyl-coenzyme M réductase (EC ).
Le composé disulfure est ensuite réduit à l'aide de la coenzyme F420 pour régénérer les deux thiols.
Risques d'inhibition de la méthanogenèse
Une température trop basse ou trop élevée peut inhiber ce processus.
Les microbes responsables de la méthanogénèse sont également vulnérables à certains inhibiteurs, dont : certains antibiotiques ou biocides, cations, métaux lourds, sulfates, ammoniaque et acides gras volatils (dont les effets sont encore cependant controversés : certains auteurs (Aguilar et al., 1995) estiment qu'il faudrait 10 g/L de chaque acide pour avoir une inhibition significative[7] mais selon d'autres comme Yuen et al. (1995) estiment que dès 3 g/L d’AGV la méthanogénèse peut être inhibée[8]. On s'accorde néanmoins sur l'importance du suivi de ce paramètre lors de la dégradation anaérobie pour la stabilité du processus (Bolzonella et al., 2003) et sur le fait qu'en cas d'inhibition de la méthanisation les AGV s’accumulent[9],[10].
Valorisation industrielle
La méthanation qui cherche notamment à catalytiquement "reconvertir" du CO2 anthropique en méthane exploitable s'avère plus complexe qu'on ne le pensait mais elle progresse.
Parallèlement la R&D universitaire et industrielle explorent des voies plus biotechnologiques, dites de « catalyse microbienne »[11] ou biométhanation[12]. D'une manière plus ou moins biomimétique, la biométhanation industrielle cherche à s'inspirer des processus commun dans certains milieux naturels où certaines archées (groupe de procaryotes aujourd'hui distinct de celui des bactéries) utilisent le dioxyde de carbone et le dihydrogène abondants dans leur environnement pour produire du méthane (il s'agit d'une méthanogénèse naturelle, dont la première étape consiste à réduire le CO2)[13]. La plus grande partie du méthane terrestre est produit de cette manière[13].
Les électrons produits par oxydation du dihydrogène ne sont pas naturellement assez énergétiques pour spontanément réduire le CO2, mais au cours de l'évolution, certains groupes d'archées ont appris à utiliser des complexes métalloenzymatiques (Méthyl-coenzyme M réductase et coenzyme B) pour produire CoM-S-S-CoB et du méthane en dynamisant la moitié des électrons dérivés du H2 (au détriment de l'autre moitié)[13].
Wagner et al. ont montré (en 2017 dans le Journal Science) comment chez certaines archées les électrons reçoivent la poussée d'énergie nécessaire : grâce à la structure cristalline du complexe metalloenzymatique qui contient une chaîne de transfert d'électrons en forme de T, qui divise le flux d'électrons d'un seul donneur vers deux accepteurs (un accepteur doit se lier à une paire de nouveaux groupes fer-soufre à réduire)[13],[14].
Pour des usages locaux, dans les pays en développement notamment, on cherche à produire des bioréacteurs rustiques de conception et entretien simple[15]. Un prototype récent associe deux bioréacteurs solides montés en série reliés par un système d'alimentation et de recirculation[15]. Il a pu produire jusqu'à 6,35l de méthane par litre de réacteur (pour un débit d'hydrogène de 25,2 l/l de réacteur), avec une source nutritive simple (déchets organiques liquides), et acceptant un milieu de culture simple (vermiculite et grains de perlite dans l'expérimentation) ; le taux de conversion pour l'hydrogène a été de 100 %, mais le fonctionnement de ce type de réacteur reste instable et doit être amélioré[15].
Le principe du lit fluidisé a été aussi été testé (en Allemagne en 2015-2017) pour une culture anaérobie d'archées thermophiles afin obtenir un réacteur de biométhanation à haute performance non pressurisé[16]. Le prototype (d'une contenance de 58,1 litres) a pu produire 5.4 m3 de CH4/(m3·d) avec plus de 98 % de conversion en CH4. Il a aussi montré l'importance du contrôle du pH, des nutriments pour une stabilité de la production. Dans ce cas l'inoculum original était mésophile, et il s'est rapidement adapté aux conditions thermophiles du réacteur dit ATBR (pour « anaerobic trickle bed reactor »). Le fonctionnement du réacteur s'est montré sensible au phénomène de dilution par l'eau métabolique produite par la communauté microbienne. Ce type de réacteur peut être ensemencé (inoculé) simplement par des boues digérées : leur biodiversité intrinsèque les rendent capables de fournir une population microbienne qui va bien et rapidement s'adapter aux conditions thermophiles du réacteur. Dans ce cas les auteurs n'ont jamais observé de biofilm macroscopique en conditions thermophiles (même après 313 jours d'opération). Ils concluent que cette technologie est « très efficace » et a un « potentiel élevé d'utilisation comme moyen de conversion et de stockage de l'énergie ».
Un nouveau type de réacteur à flux continu a aussi été récemment (publication 2017) prototypé et testé : il contient un biofilm microbien anaérobie mésophile mixte. Le temps de séjour y est lent mais le système est simple. Dans les 82 jours après mise en route des tests, des rendements de conversion (du CO2 en CH4) de 99 % et de 90 % ont été atteints pour des débits de gaz totaux qui étaient respectivement de 100 et 150 v/v/j. Pour un débit d'entrée de gaz de 230 V/V/J, les vitesses d'évolution du méthane ont atteint 40 V/V/J (record à ce jour pour « une biométhanation par biofilm fixe », avec peu de sources de consommation "parasite" d'énergie grâce à un nouveau concept d'écoulement par bouchons faisant alterner des phases liquides (avec nutriment), solides (croissance du biofilm) et gazeuses (absorption du CO2) très différente du réacteur CSTR qui nécessite en outre un refroidissement constant et important).
Ce travail a aussi été l'occasion d'inventer un type de réacteur qui pourrait avoir d'autres usages biotechnologiques connexes[17].
En 2017 on cherche à encore améliorer l'efficience énergétique de ces voies nouvelles de biométhanation[18], qui pourrait par exemple être intégrée au processus d'épuration des eaux usées[19].
Notes et références
- (en) F. Kepler et al., « Methane emissions from terrestrial plants under aerobic conditions », Nature, vol. 439, no 7073, , p. 187-191 (PMID 16407949, DOI 10.1038/nature04420)
- (en) Science Now – 14 janvier 2009 « Methane Emissions? Don't Blame Plants ».
- (en) R. K. Thauer, « Biochemistry of Methanogenesis: a tribute to Marjory Stephenson », dans Microbiology, 1998, 144, 2377-2406.
- (en) Yuchen Liu et William B. Whiteman, « Metabolic, Phylogenetic, and Ecological Diversity of the Methanogenic Archaea », Annals of the New York Academy of Sciences, vol. 1125, , p. 171.189 (DOI 10.1196/annals.1419.019)
- (en) M. Korbas, S. Vogt, W. Meyer-Klaucke, E. Bill, E. J. Lyon, R. K. Thauer, et S. Shima, « The Iron-Sulfur Cluster-free Hydrogenase (Hmd) Is a Metalloenzyme with a Novel Iron Binding Motif », dans Journal of Biological Chemistry, 2006, 281, 30804-30813.
- (en) J. Lengeler, G. Drews et H. Schlegel, Biology of the Prokaryotes, Stuttgart, G. Thieme, , 955 p. (ISBN 978-0-632-05357-5), p. 291
- Aguilar A., Casas C. & Lema J.M. (1995). Degradation of volatile fatty acids by differently enriched methanogenic cultures- kinetics and inhibition. Wat. Res, 29(2), 505-509.
- YUEN S.T.S., STYLES J.R. & MCMAHON T.A. (1995). An active landfill management by leachate recirculation – a review and an outline of a full-scale project. In : T.H. Christensen, R. Cossu & R. Stegmann (Eds). Proceedings Sardinia 95, Fifth International Landfill Symposium, 1-17.
- AHRING B.K., SANDBERG M. & ANGELIDAKI I. (1995). Volatile fatty acids as indicators of process imbalance in anaerobic digestors. Aplied Microbiology and Biotechnology, 43, 559-565.
- WENS P., VERCAUTEREN T., DE WINDT W. & VERSTRAETE W. (2001). Factors Inhibiting Anaerobic Degradation in a Landfill. In : T.H. Christensen, R. Cossu & R. Stegmann (Eds). Proceedings Sardinia 2001, Eighth International Waste Management and Landfill Symposium, S. Margherita di Pula, Cagliary, Italy, CISA.
- Lecker, B., Illi, L., Lemmer, A., & Oechsner, H. (2017) Biological hydrogen methanation–A review. Bioresource Technology, 245, 1220-1228.
- Serge R. Guiot, « Biométhanation du gaz de synthèse par couplage avec la digestion anaérobie », L'Eau, l'Industrie, les Nuisances, (lire en ligne).
- Tristan Wagner, Jürgen Koch, Ulrich Ermler & Seigo Shima (2017), Methanogenic heterodisulfide reductase (HdrABC-MvhAGD) uses two noncubane [4Fe-4S] clusters for reduction |Science | 18 aout 2017 | Vol. 357, Issue 6352, pp. 699-703 | DOI: 10.1126/science.aan0425 | résumé
- Holger Dobbek (2017) Playing marble run to make methane| Science 18 Aug 2017:Vol. 357, Issue 6352, pp. 642-643 |DOI: 10.1126/science.aao2439 | résumé
- Alitalo A, Niskanen M & Aurab E (2015) Biocatalytic methanation of hydrogen and carbon dioxide in a fixed bed bioreactor Bioresource Technology |Volume 196, Novembre 2015, Pages 600–605 | résumé
- Strübing, D., Huber, B., Lebuhn, M., Drewes, J. E., & Koch, K. (2017). High performance biological methanation in a thermophilic anaerobic trickle bed reactor. Bioresource Technology, 245, 1176-1183|résumé
- Savvas, S., Donnelly, J., Patterson, T., Chong, Z. S., & Esteves, S. R. (2017). Biological methanation of CO 2 in a novel biofilm plug-flow reactor: A high rate and low parasitic energy process. Applied Energy, 202, 238-247. résumé
- Strübing, D., Huber, B., Lebuhn, M., Drewes, J. E., & Koch, K. (2017). High performance biological methanation in a thermophilic anaerobic trickle bed reactor. Bioresource Technology, 245, 1176-1183.
- Patterson, T., Savvas, S., Chong, A., Law, I., Dinsdale, R., & Esteves, S. (2017). Integration of Power to Methane in a waste water treatment plant–A feasibility study. Bioresource Technology, 245, 1049-1057.
Voir aussi
Articles connexes
Bibliographie
- Strübing, D., Huber, B., Lebuhn, M., Drewes, J. E., & Koch, K. (2017). High performance biological methanation in a thermophilic anaerobic trickle bed reactor. Bioresource Technology, 245, 1176-1183|résumé.
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire  Portail de la biochimie
Portail de la biochimie  Portail de la chimie
Portail de la chimie