Trichromatisme
Le trichromatisme, la trichromatie ou le trichromaticisme, est le traitement de l'information visuelle au moyen de trois canaux restituant la couleur. La vision est dite trichromatique et l'organisme est dit trichromate : c'est le cas dans l'espèce humaine. Les récepteurs visuels qui différencient les longueurs d'onde de la lumière comportent trois types de cônes qui tapissent la rétine humaine.

Le trichromatisme repose sur la présence, dans la rétine, de trois sortes de photorécepteurs spécifiques à la couleur (nommés cônes chez les vertébrés) et dont le spectre d'absorption est différent. Il est fréquent que des photorécepteurs soient actifs à des niveaux d'intensité lumineuse différents et que leur nombre soit en conséquence supérieur à trois. En effet, chez les vertébrés qui sont dotés de trois sortes de cônes, les bâtonnets peuvent également contribuer à la perception des couleurs en faible lumière. Ces conditions spéciales réservent donc une petite place au tétrachromatisme dans l'espace colorimétrique.
Les humains et autres animaux trichromates
Les humains et quelques autres primates possèdent une vision trichromate, partiellement imputable aux pigments hérités d'organismes vertébrés plus anciens. Chez les oiseaux et les poissons par exemple, on retrouve quatre pigments. Les pigments de cônes récepteurs supplémentaires détectent des longueurs d'onde d'énergie différente, dont les ultraviolets font parfois partie. Le trichromatisme de certains primates résulte d'une perte de deux de ces pigments chez les mammifères placentaires, et de l'apparition d'un autre pigment[1].
Les humains et les primates les plus proches sont des trichromates, au même titre qu'une partie des femelles de la plupart des espèces de singes platyrhiniens (dits du nouveau monde) ainsi que les mâles et les femelles alouates (dits singes hurleurs)[2]. Une étude récente suggère que le trichromatisme pourrait également être répandu parmi les marsupiaux[3]. Une autre étude portant sur le trichromatisme des marsupiaux australiens suggère que les cônes photorécepteurs spécifiques aux longueurs d'onde moyennes de la souris à miel et du dunnart à pieds étroits sont hérités du système rétinien des reptiles. Le possible trichromatisme des marsupiaux résulte d'une évolution potentiellement différente de celle des primates. D'autres expérimentations animales portant sur la biologie et le comportement des marsupiaux pourraient confirmer leur trichromatisme[4].
La plupart des autres mammifères sont aujourd'hui considérés comme des dichromates, c'est-à-dire avec deux sortes de cônes seulement. Les études portant sur les carnivores révèlent généralement un dichromatisme semblable à celui des autres mammifères, comme chez le furet[5] et la hyène[6]. Certaines espèces d'insectes telles que les abeilles sont également des trichromates. Elles sont sensibles aux ultraviolets au bleu et au vert plutôt qu'au bleu au vert et au rouge[2].
Avantages sélectifs
La recherche indique que le trichromatisme serait un avantage sélectif car il permet de distinguer tout ce qui n'a pas la même couleur que celle d'un feuillage mature : les fruits et les jeunes pousses de la végétation[7]. Chez les catarhiniens, les études de la sensibilité spectrale des cônes M et L montrent qu'elle est optimale pour discriminer un objet au milieu d'un feuillage[8].
Une autre hypothèse propose d'envisager que la détection du rougissement de l'épiderme (et des humeurs qui l'accompagnent) pourrait avoir influencé le développement du trichromatisme de la vision chez les primates. La couleur rouge est réputée avoir d'autres effets sur le comportement des primates, humains compris. Cela fait partie de la psychologie des couleurs[9].
Types de cônes propres aux primates
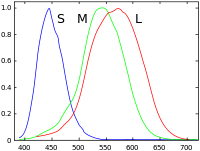
Les primates sont les seuls mammifères placentaires trichromates connus[10]. Leurs rétines comportent trois sortes de cônes. Chaque type de cône contient un photo-pigment différent (opsines) auquel correspond une photosensibilité maximale : couleurs bleuâtres avec un maximum dans les violet-bleus (longueurs d'onde courte, cônes S pour « short » en anglais), couleurs vertes (longueurs d'onde moyenne, cônes M pour « medium » en anglais) et couleur vertes à rouges avec un maximum vert-jaune (grande longueur d'onde, cônes L pour « long » en anglais) du spectre visible (Schnapf et al, 1987).
Les cônes S représentent environ 5 à 10 % des cônes et sont répartis comme une mosaïque régulière. Des cellules bipolaires et ganglionnaires dédiées transmettent le message des cônes S, tandis que nous possédons la preuve qu'ils disposent également d'un canal distinct au travers du thalamus jusqu'au cortex visuel. Au contraire, les cônes M et L sont difficiles à distinguer, tant par leur forme que par les autres moyens anatomiques car leurs opsines ne diffèrent que de 15 acides aminés sur 363 (personne n'est encore parvenu à désigner des anticorps qui leur soient spécifiques). Toutefois, Mollon et Bowmaker ont pu constater que les cônes M et L étaient distribués aléatoirement et dans des proportions semblables[11].
Mécanisme de la perception trichromique
La perception trichromique est la capacité des humains et de certains autres animaux à voir les couleurs différentes par l'intermédiaire de trois types de cônes photorécepteurs. La théorie de la trichromie a débuté avec les constatations des peintres et teinturiers qui constatent qu'on peut recréer une quantité de sensations colorées avec seulement trois couleurs primaires. Thomas Young forme dès la fin du XVIIIe siècle l'hypothèse que la perception des couleurs pourrait résulter de trois différentes sortes de cellules photoréceptrices. Plus tard, Hermann Ludwig von Helmholtz a développé les idées de Young en conduisant des expériences qui démontraient que les personnes dont la vision était normale pouvaient reproduire le spectre des couleurs normales à partir de trois couleurs par un exercice de concordance. Les preuves physiologiques de la trichromie n'ont été établies que bien plus tardivement, par Gunnar Svaetichin, physiologiste suédo-finno-vénézuélien, en 1956[12].
Chacun des trois types de cônes de la rétine humaine abrite une sorte de pigment photorécepteur distincte. Ces pigments sont constitués d'une protéine transmembranaire appelée opsine ainsi que d'une molécule photosensible appelée rétinal. Chaque pigment est particulièrement sensible à une certaine longueur d'onde. Cela signifie qu'un pigment a toutes les chances de produire une réponse cellulaire lorsqu'il est atteint par un photon dont l'énergie spécifique correspond à la longueur d'onde à laquelle le pigment est le plus sensible. Les trois différentes sortes de cônes L, M et S réagissent respectivement aux grandes longueurs d'onde (maximum à 564 nm), aux longueurs d'onde moyennes (maximum à 534 nm) et aux longueurs d'onde courtes (420 nm)[13],[14].
Puisqu'un cône réagit en fonction de la longueur d'onde et de l'intensité de la lumière, le cerveau n'aurait aucun moyen de différencier les couleurs s'il ne disposait que d'un seul type de cônes. Il faut donc au moins deux types de cônes pour pouvoir percevoir les couleurs. À partir de deux types de cônes, le cerveau peut comparer les signaux de chaque type et différencier deux lumières de longueur d'onde dominante différente. Prenons l'exemple d'une stimulation modérée d'un cône M (longueur d'onde moyenne). Cela pourrait indiquer une intense lumière rouge (grande longueur d'onde), ou une lumière d'intensité modérée jaune-vert (longueur d'onde moyenne). La distinction est rendue possible par les cônes L (grande longueur d'onde) qui seraient, si la lumière est rouge, plus fortement stimulés que les cônes M. Si la lumière était bleu-vert, la stimulation du cône M pourrait être identique, mais le cône L réagirait beaucoup moins. Cependant, une lumière jaune ou blanche produiraient exactement la même stimulation. Avec une vision trichrome, les cônes S permettent de faire la différence, en même temps qu'ils étendent la lumière perçue du côté des bleus. La vision trichromique se produit donc par la combinaison des signaux cellulaires.
On estime que chaque type de cône de la rétine humaine peut distinguer environ 100 gradations d'intensités différentes. Si le cerveau humain est effectivement capable de combiner toutes ces variations de toutes les manières possibles, l'humain moyen devrait être en mesure de distinguer environ un million de couleurs différentes[15],[16]. Les estimations basées sur des expériences de différence de couleur arrivent entre 15 000 couleurs identifiables et 500 000 couleurs séparées par une différence juste perceptible. Le million de couleurs semble être une valeur extrême. Ces valeurs sont basées sur des calculs, une partie allant jusqu'à la moitié de ces couleurs théoriques n'existe pas dans la nature ni n'est réalisable par les moyens techniques actuels[17].
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Trichromacy » (voir la liste des auteurs).
- (en) Catherine Arrese, « Trichromacy in Australian Marsupials », Current Biology, vol. 12, , p. 657–660 (PMID 11967153, lire en ligne)
- Rowe, Michael H (2002). "Trichromatic color vision in primates." News in Physiological Sciences. 17(3), 93-98.
- Arrese, CA; Oddy, AY; Runham, PB; Hart, NS; Shand, J; Hunt, DM (2005). "Cone topography and spectral sensitivity in two potentially trichromatic marsupials, the quokka (Setonix brachyurus) and quenda (Isoodon obesulus)." Proceedings of the Royal Society of London Series B. 272(1595), 791-796.
- (en) Catherine Arrese, « Trichromacy in Australian Marsupials », Current Biology, vol. 12, no 8, , p. 657–660 (PMID 11967153, lire en ligne)
- Calderone, JB; Jacobs, GH (2003). "Spectral properties and retinal distribution of ferret cones." Visual Neuroscience. 20(1), 11-17.
- Calderone, JB; Reese, BE; Jacobs, GH (2003). "Topography of photoreceptors and retinal ganglion cells in the spotted hyena (Crocuta crocuta)." Brain Behavior and Evolution. 62(4), 182-192.
- Sharpe et al. (2006). Advantages and disadvantages of human dichromacy. Journal of Vision, 6, 213-223. Retrieved from http://www.journalofvision.org/content/6/3/3.full.pdf+html
- Sumner, P. (2000). "Catarrhine photopigments are optimized for detecting targets against a foliage background". Journal of Experimental Biology
- Diana Widermann, Robert A. Barton, and Russel A. Hill. Evolutionary perspectives on sport and competition. In DOI:10.1093/acprof:oso/9780199586073.001.0001
- (en) Ronald G. Boothe, Perception of the visual environment, Springer, (ISBN 978-0-387-98790-3, lire en ligne), p. 219
- (en) Heinz Wässle, « Colour vision: A patchwork of cones », Nature, vol. 397, , p. 473–475 (DOI 10.1038/17216, lire en ligne)
- Svaetichin,G. (1956). Spectral response curves from single cones, Actaphysiol. scand. 39, Suppl. 134, 17-46.
- (en) Kandel ER, Schwartz JH, Jessell TM, Principles of Neural Science, New York, McGraw-Hill, , 182–185 p. (ISBN 0-8385-7701-6)
- (en) Jacobs GH, Nathans J, « Color Vision: How Our Eyes Reflect Primate Evolution », Scientific American,
- (en) Mark Roth, « Some women who are tetrachromats may see 100,000,000 colors, thanks to their genes », Pittsburgh Post-Gazette, september 13, 2006]
- "Color Vision:Almost Reason for Having Eyes" by Jay Neitz, Joseph Carroll, and Maureen Neitz Optics & Photonics News January 2001 1047-6938/01/01/0026/8- Optical Society of America
- Robert Sève, Science de la couleur : Aspects physiques et perceptifs, Marseille, Chalagam, , p. 229.
Articles connexes
- Œil humain
- Photochromie, trichromie
- Tétrachromatisme
Liens externes
- The Straight Dope: "Are cats and dogs really color-blind? How do they know?"
- Scientific Paper: "Retinal mechanisms for chromatic and achromatic vision"
