Oncogène
Les oncogènes (du grec onkos, signifiant vrac, masse ou tumeur et génos signifiant génération, naissance, origine) sont une catégorie de gènes dont l'expression favorise la survenue de cancers. Ce sont des gènes qui commandent la synthèse d'oncoprotéines, protéines stimulant la division cellulaire ou inhibant la mort cellulaire programmée (apoptose), ce qui déclenche une prolifération désordonnée des cellules.
Le terme oncogène peut désigner aussi des virus qui provoquent l'apparition de cancers. On parle d'oncovirus.
Introduction
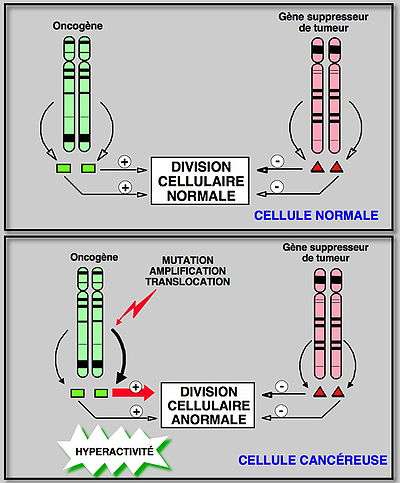
Notre organisme est composé d'environ 1014 cellules réparties dans plus de 200 types cellulaires différents qui composent les tissus (cellules sanguines, nerveuses, germinales, etc.). La prolifération cellulaire au sein de ces tissus est rigoureusement contrôlée au cours de notre vie ; certaines cellules (les neurones) ne nécessitant pas un renouvellement constant, d'autres étant perpétuellement en cours de multiplication (cellules sanguines ou de la peau). Le contrôle de cette multiplication cellulaire normale se fait par l'intermédiaire d'un équilibre permanent entre facteurs activateurs (stimulateurs de la division cellulaire) et facteurs inhibiteurs (freins de la division cellulaire). Toute altération de cet équilibre, ou homéostasie cellulaire, peut faire pencher la balance soit du côté inhibiteur, dans ce cas la cellule meurt et disparaît, soit du côté activateur et la cellule se divise de façon incontrôlée et peut donner naissance à un cancer.
De manière schématique, on peut faire une analogie entre une voiture et une cellule. Lorsque les systèmes qui contrôlent l’accélérateur et les freins de la voiture sont en bon état, celle-ci va donc pouvoir avoir une vitesse de croisière parfaitement contrôlée. Une voiture dans laquelle l’accélérateur serait activé en permanence ou bien ayant une absence totale de frein est donc lancée dans une course folle qui ne peut plus être arrêtée sauf par une catastrophe ! Dans les cellules cancéreuses, cela se passe exactement de la même façon.
Il y a trois grandes catégories de gènes associés aux pathologies cancéreuses : les oncogènes (cet article), les gènes suppresseur de tumeurs et les gènes de réparation de l'ADN.
Les proto-oncogènes sont les régulateurs positifs de la prolifération cellulaire (les «accélérateurs»). Ils deviennent hyper-actifs et leur modification (en oncogène) est dominante car il suffit qu'un des deux allèles soit muté. On a identifié actuellement plus de 100 proto-oncogènes. Parmi les plus connus, on trouve les gènes Ha-ras, myc et abl.
La seconde catégorie comprend les gènes suppresseurs de tumeurs qui sont des régulateurs négatifs de la prolifération cellulaire (les «freins») avec notamment la protéine p53. Son inactivation est l'initiation la plus fréquente de la cancérogenèse, c'est une mutation de type récessif. L'inactivation d'un anti-oncogène ou l'activation d'un proto-oncogène a des effets similaires sur la prolifération cellulaire et la survenue d'anomalies génétiques.
Il existe également les gènes de réparation qui sont capables de détecter et de réparer les lésions de l’ADN qui ont modifié les oncogènes ou les gènes suppresseurs de tumeur. Ces systèmes de réparation sont également inactivés dans les cellules cancéreuses.
Les cancers sont des pathologies génétiques, c’est-à-dire qu’ils ont pour origine une modification quantitative et/ou qualitative des gènes décrits ci-dessus. Comme il s’agit d’altérations génétiques somatiques qui ne sont présentes que dans le tissu malade, la plupart des cancers ne sont donc pas héréditaires. Les cancers familiaux (10 % des cancers humains) sont associés à une altération constitutionnelle (ou germinale) d’un gène. Cette altération est donc présente dans toutes les cellules de notre organisme, gamètes incluses, elle peut donc être transmise à la descendance.
Oncogènes
Sémantique et histoire du terme
La transformation cellulaire correspond au passage d’une cellule eucaryote normale vers un état cancéreux. À ne pas confondre avec la transformation bactérienne qui est un phénomène tout à fait différent.
Les termes c-onc et v-onc ont été les premiers à être utilisés. V-onc (oncogène viral) correspond à une séquence d’ADN (un gène généralement) porté par un rétrovirus transformant (Virus a ARN). Cette séquence v-onc est responsable du pouvoir de cancérisation du rétrovirus. Lorsque D. Stehelin et collaborateurs ont montré en 1976 que ces séquences v-onc correspondent a des gènes cellulaires que le virus a modifié et incorporé dans son génome, ces gènes cellulaires ont été nommés c-onc (oncogène cellulaire ou proto-oncogène). Ces rétrovirus sont spécifiques des animaux (souris poulet ou autre). Aucun rétrovirus associé à une séquence v-onc n a été mis en évidence dans une tumeur humaine. L’analyse de ces rétrovirus ont permis d’isoler de nombreux v-onc et par conséquent d’identifier leurs équivalents cellulaires c-onc. Par la suite, l’analyse génétique plus détaillée des tumeurs humaines a permis de mettre en évidence les faits suivants :
- La plupart des cancers humains ne sont pas dus à des rétrovirus
- La transformation cancéreuse est due à une altération accidentelle de gènes cellulaires dont certains sont les c-onc identifié chez les rétrovirus animaux ;
- Tous les c-onc identifiés chez les rétrovirus ne sont pas associés à des cancers chez l'homme ;
- De nouveaux oncogènes ont été identifiés dans les tumeurs humains sans avoir d’équivalent v-onc dans les rétrovirus. De manière globale, on nomme « oncogène cellulaire » toute séquence d’ADN dont la modification qualitative ou quantitative conduit à la transformation cellulaire.
Oncogènes et cancers
En général, on considère qu'il existe trois grands types de modifications génétiques qui peuvent altérer un proto-oncogène: les mutations, les translocations et les amplifications géniques. Dans tous les cas, on obtient un produit hyperactif qui stimule la division cellulaire de sorte qu'elle échappe à la régulation normale.
- Les mutations sont des évènements ponctuels qui peuvent changer l'une des bases de l’ADN du gène. Cette modification change l'information génétique et la protéine synthétisée à partir de ce gène est modifiée. Un des proto-oncogènes mutés dans les cancers humains est le gène H-ras. Sous sa forme normale (non mutée), la protéine H-ras stimule la multiplication cellulaire uniquement lorsqu’elle reçoit un signal de prolifération. Sous sa forme mutée, le gène H-ras code une protéine qui stimule la multiplication cellulaire indépendamment de tout signal.
- Les amplifications géniques correspondent à l'augmentation du nombre de copies d'un proto-oncogène. Dans les cellules de neuroblastomes, on retrouve plusieurs centaines de copies du gène N-myc alors qu'il devrait n'y avoir que deux copies. Cette amplification aboutit à une surproduction de la protéine myc qui active continuellement la division cellulaire. De même l'amplification du gène erbB2 est fréquente dans les cancers du sein.
- Les translocations chromosomiques sont des modifications de notre génome qui correspondent à des remaniements importants de nos chromosomes. En général, dans les translocations réciproques, un fragment de l'un de nos chromosomes est transféré sur un autre chromosome et vice-versa. Cet évènement peut générer deux types de modifications. Des modifications quantitatives car la translocation change complètement l'environnement génétique des gènes présents sur les fragments de chromosome et dérègle leur expression. Il peut y avoir aussi des modifications qualitatives car la translocation peut engendrer la synthèse de protéines ayant des fonctions aberrantes. Les translocations sont très fréquentes dans les leucémies. Les cas les plus connus sont ceux de la Leucémie Myéloïde Chronique avec une translocation systématique entre les chromosomes 9 et 22 ou la translocation entre les chromosomes 14 et 18 dans les lymphome

Différents oncogènes
Le premier oncogène viral, le gène v-SRC du virus du sarcome de poulet découvert par Rous en 1911, fut cloné en 1975. L'étude des oncogènes viraux (v-onc) a permis de caractériser leurs équivalents cellulaires normaux les proto-oncogènes (c-onc).
| Nom | Cancer (*) | Type d'altération (**) |
|---|---|---|
| HRAS / KRAS et NRAS | Pratiquement tous les cancers humains | Mutations ponctuelles au niveau des codons 12 et 13 |
| Her2 (ErbB2 ou neu) | Cancer du sein (30 %) | Amplification du gène |
| Hdm2 | Sarcomes | Amplification du gène |
| N-Myc | Neuroblastomes (20 %) | Amplification du gène |
| TAL1 | Leucémie a cellules T (30 % des ALL-T) | Réarrangement du gène |
| Fli-1 | Sarcome d'Ewing (80 %) | Translocation chromosomique |
| MLL | Leucémies d'origines diverses | Translocation chromosomique |
| CCND1 (cycline D1) | Cancers du sein (15 %), Tête et cou (30 %) | Amplification du gène |
| RET | Cancer médullaire de la thyroïde (35 %) | Translocation chromosomique |
| PML | Leucémie aigües promyélocitaires | Translocation chromosomique |
| Beta caténine | Cancer du colon | Mutations ponctuelles |
| Abl | Leucémie Myéloïde Chronique (95 %) | Translocation chromosomique |
| c-myc | Pratiquement tous les cancers humains | Translocation chromosomique, amplification du gène |
(*) seuls les cancers les plus fréquents sont indiqués (**) seules les modifications les plus fréquentes sont indiquées
les oncogènes cellulaires comme c-fos, c-src, c-jun (et autres) dont l'association avec des cancers humains n'est pas formellement avérée ne sont pas inclus dans cette liste. De nombreux oncogènes et gènes suppresseurs de tumeurs isolés chez l'animal ne sont pas associés a des pathologies cancéreuses chez l'homme.
Les oncogènes comme cibles pour le développement de nouvelles thérapies
Les oncogènes sont des cibles privilégiées pour le développement de nouvelles molécules anticancéreuses. Les oncogènes ayant un effet activateur sur la prolifération cellulaire, il semble logique de penser que leur inhibition pourrait avoir un effet anticancéreux. D’autre part, de nombreux oncogènes étant différents par rapport aux proto-oncogènes des cellules normales (formes mutées, protéines de fusion) on peut penser que seules les cellules tumorales seront ciblées par ces nouvelles molécules. Parmi les milliers de molécules qui ont été développées par les laboratoires de recherche et l’industrie pharmaceutique, seule une poignée est actuellement utilisée de manière systématique en thérapie anticancéreuse.
L’exemple le plus marquant de cette stratégie thérapeutique a été le développement du STI571 (imatinib, ou Glivec), une molécule qui inhibe spécifiquement l’oncogène Bcr-Abl qui est retrouvé dans près de 100 % des leucémies myéloïdes chroniques. Par la suite, il a été découvert que cette molécule a également une activité thérapeutique très efficace vis-à-vis de certains cancers du tube digestif appelés tumeur stromale gastro-intestinale, qui étaient très résistantes à toute autre thérapie jusqu'à présent. Dans ce cas, l’oncogène ciblé est c-kit.
Deux substances, l'arsenic et l'acide rétinoïque, ciblant spécifiquement l'oncogène de la leucémie aiguë promyélocytaire, ont un effet thérapeutique chez les malades atteints par ce type de leucémie[1].
Notes et références
- (en) J. Zhu et al., « Arsenic-induced PML targeting onto nuclear bodies : implications for the treatment of acute promyelocytic leukemia », revue PNAS, vol. 94, , p. 3978-3983
Voir aussi
Articles connexes
- Une revue scientifique dédiée aux oncogènes et gènes suppresseurs de tumeurs : Oncogene
Liens externes
- (en) Oncogenes and Tumor Suppressor Genes
- « The hallmarks of cancer », Hanahan D, Weinberg RA Cell, no 100, p. 57–70 (2000).
- (en) A genetic model for colorectal tumorigenesis. Fearon ER, Vogelstein B (1900) Cell, no 61, p. 759–767.
- (en) Cancer genes and the pathways they control. Vogelstein B, Kinzler kW (2004) Nat. Med., no 10, p. 789–799.

