Paradoxe de Peto
Le paradoxe de Peto est le constat, effectué par Richard Peto (en), qu'au niveau des espèces, l'incidence du cancer ne paraît pas être en corrélation avec le nombre de cellules de l'organisme[1]. Par exemple, l'incidence des cancers chez l'être humain est bien plus élevée que chez les baleines[2], et ceci malgré le fait qu'une baleine a beaucoup plus de cellules qu'un être humain. Si la probabilité de carcinogenèse était constante pour une cellule donnée, on observerait au contraire une incidence beaucoup plus forte pour la baleine que pour l'homme.
Le paradoxe
Carcinogénèse animale


Le cancer touche probablement toutes les espèces de vertébrés.
On trouve les mêmes types de cancers chez tous les mammifères[3],[4], et les mécanismes de base qui conduisent à ces cancers sont également très similaires.
Par ailleurs, de nombreux mécanismes inhibant l'apparition de cancers, tels que les gènes suppresseurs de tumeurs, sont présents chez tous les mammifères[5].
C'est cette similarité qui permet, pour la recherche sur le cancer, d'utiliser des mammifères comme modèle animal du cancer humain[6].. Que ce soit dans la structure cellulaire et son métabolisme, ou dans les mécanismes de formation et de propagation du cancer, la classe des mammifères est d'une grande homogénéité, depuis la Kitti à nez de porc (Craseonycteris thonglongyai) et ses deux grammes jusqu'à la baleine bleue (Balaenoptera musculus) et ses deux cents tonnes — cent millions de fois plus grosse.
Chez les mammifères, le taux de cancer est relativement constant, variant d'un facteur deux. On a de nombreux exemples de cancers et de morts par cancers rencontrés chez tous les mammifères, et l'on dispose de statistiques fiables sur le taux de cancer pour de nombreuses espèces servant de modèle animal. En revanche, on dispose de relativement peu de données sur les espèces sauvages[5]..
Pour les souris de laboratoire (Mus musculus), l'incidence du cancer est d'environ 46%[7]. Pour le chien (Canis lupus familiaris), le taux est d'environ 20 %[8] et chez l'homme il est de 22 % d'après les statistiques de mortalité aux États-Unis[9]. On rencontre également des cas de cancers chez la baleine bleue (Balaenoptera musculus), et bien qu'il n'y a pas de statistique très précise sur le sujet, on suppose que la plupart des baleines ne meurent pas de cancer[10]. Une étude de 1966 sur 2 000 baleines à fanon (Mysticeti) dans la baie de Saldanha n'a mis en évidence aucune trace de cancer[11]. Mais l'autopsie de 129 des 263 bélugas échoués entre 1983 et 1999 sur les rives du Saint-Laurent a révélé un taux de cancer de 27 %, et dans 18 % des cas, ce cancer avait été la cause principale de la mort[12]. Ce taux paraît très élevé pour des baleines, et il est probable qu'aucune autre population de baleine ne présente un taux aussi élevé. On soupçonne la pollution industrielle et agricole de l'embouchure d'en être la cause[6],[13],[14],[15],[16].
Sensibilité au cancer
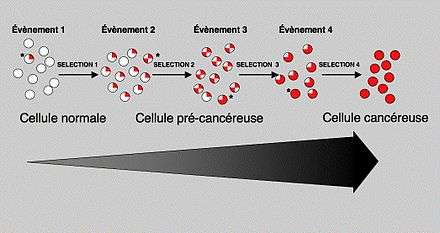
Si l'on en croit la théorie généralement admise sur le développement du cancer (la cancérogenèse), le premier évènement conduisant à la formation d'un cancer est une défaillance au niveau cellulaire, qui transforme une cellule normale en une cellule tumorale maligne. Les tumeurs apparaissent lorsque environ une demi douzaine de gènes participant au contrôle de la croissance cellulaire ont muté. La cellule se multiplie anarchiquement, l'amas se transforme et passe (plus ou moins rapidement, moyennant des temps de latence et des probabilités de transition, suite à des mutations complémentaires) par une série d'étapes, jusqu'aux métastases mortelles[17].
De ce fait, si l'on en croit la théorie (plus controversée) du développement linéaire sans seuil du cancer, il y a une relation linéaire entre la défaillance initiale au niveau cellulaire et l'émergence finale d'un cancer : chaque cellule a une probabilité élémentaire d'induire un cancer, et la probabilité finale au niveau de l'organisme dépend alors linéairement du nombre de ses cellules.
Formulation initiale
Peto, un statisticien épidémiologiste de l'université d'Oxford, publia la première formulation de ce paradoxe en 1977[18]. Alors qu'il préparait un article de synthèse sur le modèle multi-étape de la carcinogenèse (proposé en 1954 par Peter Armitage et Richard Doll), Peto constata que à nombre de cellules équivalents, les êtres humains étaient beaucoup moins sujets au cancer que les souris :
- « Un homme a 1000 fois plus de cellules qu'une souris [...] et vit typiquement trente fois plus longtemps. En soumettant deux organismes similaires au risque de cancer, l'un sur une durée trente fois plus grande que l'autre, on devrait trouver un facteur de l'ordre de 304 ou 306 (de l'ordre du million ou du milliard) sur le risque de carcinome induit par une cellule épithéliale. Et pourtant, il semble que dans la nature, le risque de carcinome induit chez la souris et chez l'homme ne sont pas aussi disparates. Se pourrait-il, alors, que les cellules de notre espèce soient réellement un milliard ou mille milliards de fois plus « cancerifuges » que les cellules de l'espèce murine? Biologiquement parlant, c'est tout à fait improbable. Si l'ADN humain n'est pas plus résistant à la carcinogenèse in vitro que celui de la souris, pourquoi ne mourrons-nous pas tous de multiples cancers à un age précoce? » (Epidemiology and Multistage Models", 1977)[17]
Peto conclut en suggérant que des évolutions adaptatives pouvaient être responsables de la variation interspécifique des taux de carcinogenèse.
Preuves expérimentales du paradoxe
À l'intérieur d'une même espèce, le risque de cancer et la taille corporelle semblent être corrélés positivement, même en tenant compte des autres facteurs de risque[19]. Une étude longitudinale de 25 ans, portant sur 17 738 fonctionnaires britanniques masculins, montra une corrélation positive entre la taille et l'incidence du cancer, avec un haut degré de significativité, même en contrôlant l'effet des autres facteurs de risque comme le tabagisme[20]. Une étude similaire publiée en 2011, portant sur plus d'un million de femmes britanniques, a montré une corrélation forte entre cancer et taille, même après prise en compte d'un grand nombre de facteurs de risque tant socioéconomiques que comportementaux[21]. Une analyse des causes de mortalité de 74 556 chiens domestiques également publiée en 2011 montre que l'incidence de cancer est plus faible dans les petites races, confirmant le résultat d'études antérieures[22].
D'une espèce à l'autre, cependant, cette corrélation n'est plus observée. Une étude de 2015 des observations de nécropsies du zoo de San Diego, portant sur 36 espèces différentes de mammifères allant de la souris rayée de 28 g à l'éléphant de 4 800 kg, près de 100 000 fois plus massif. Cette étude n'a pas trouvé de corrélation entre la taille corporelle et l'incidence de cancer, apportant un soutien expérimental à l'observation initiale de Peto[23].
Étude des mécanisme de contrôle du cancer
Explications possibles
L'explication du paradoxe de Peto fait l'objet de nombreuses hypothèses controversées.
Pour certains scientifiques, le taux de mutation chez les mammifères dépend peut-être de leur taille. Les grands mammifères auraient donc un taux de mutation plus faible que petit mammifère. Les différents taux de mutation viendraient d'une pression sélective liée à la taille[24],[25].
D'autres chercheurs pensent que les mécanismes de réparation et le système immunitaire sont plus performants pour les grands mammifères que pour les petits, de sorte qu'ils auraient une meilleure résistance au cancer[26],[5].
Nécessité pour les multicellulaires
Dans une certaine mesure, l'apparition de la vie multicellulaire a demandé un certain contrôle du taux de cancer[27], et des liens ont été identifiés entre l'origine de la vie multicellulaire et le cancer[28],[29].
Afin de pouvoir constituer des corps plus gros et d'une durée de vie plus grande, les organismes ont eu besoin d'une meilleure répression du cancer. Ces preuves expérimentales suggèrent que les gros organismes comme les éléphants ont plus de traits adaptatifs leur permettant d'échapper au cancer[30]. La raison pour laquelle les organismes de taille intermédiaire n'ont pas ces mécanismes de contrôle est peut-être que l'avantage sélectif que ces gènes induisent par leur contrôle sur le cancer est compensé par d'autres inconvénients — notamment une fertilité réduite[31]
Variété des mécanismes
Le mécanisme pour réduire le risque de cancer varie d'une espèce à l'autre[32].
Un article de Cell Reports de janvier 2015 revendique la découverte chez la baleine boréale (Balaena mysticetus) de gènes qui pourraient être associés à la longévité[33].
À peu près à la même époque, une équipe de chercheurs a identifié sur le rat-taupe nu (Heterocephalus glaber ) un polysaccharide qui semble bloquer le développement des tumeurs[34].
Considérations évolutionnaires
En octobre 2015, deux études indépendantes ont montré que les éléphants avaient vingt copies du gène P53 suppresseur de tumeurs, alors que les hommes et autres mamifères n'en ont qu'une[35]. Des recherches complémentaires ont constaté la présence de 14 copies de ce gène dans l'ADN des mammouths retrouvés congelés, mais un seul chez le lamantin ou le damans, les plus proches parents des éléphants[36].
Ces résultats suggèrent une relation due à la sélection naturelle entre la taille de l'animal et sa capacité à inhiber les tumeurs, confortant l'hypothèse de Peto.
Recherches médicales
La capacité apparente des gros animaux à inhiber le cancer sur un grand nombre de cellules a fait proliférer tout un domaine de recherches médicales[31].
Dans l'une de ces recherches, des souris de laboratoire étaient modifiées génétiquement pour qu'elles expriment les antigènes du P53 de la même manière que chez les éléphants. Les souris mutantes avaient effectivement une meilleure performance dans l'inhibition des cancers, mais présentaient également des signes de vieillissement prématuré[37].
Explications sans mécanisme adaptatif
Hypothèses implicites du modèle de Peto
Le paradoxe de Peto se fonde sur un modèle probabiliste, où l'origine d'un cancer vient d'une défaillance cellulaire, laquelle a une certaine probabilité de se multiplier et de se répandre par la suite. Mais pour que le paradoxe se manifeste, il est nécessaire que les cellules défaillantes ne fassent pas l'objet d'une destruction ciblée, par exemple par le système immunitaire de l'organisme, ce qui entraînerait leur disparition différentielle.
Inversement, si le système immunitaire (ou tout autre mécanisme) est capable de reconnaître et de détruire des cellules atypiques présentes à une certaine densité, le taux de cancer émergent ne dépendra pas uniquement du taux de défaillance des cellules (avec une occurrence de défaillance proportionnelle au nombre de cellules), mais dépendra également de la capacité globale du système immunitaire à s'activer puis combattre efficacement un pourcentage donné de cellules atypiques, présentes dans les tissus.
Dans ce dernier cas, l'incidence de cancer ne dépendra pas directement du nombre de cellules de l'organisme, mais des seuils d'activation et de saturation du système immunitaire face à des cellules atypiques - ce qui dépend du métabolisme de l'organisme dans sa globalité, et de l'efficacité de son système immunitaire, et non de sa taille.
Influence du métabolisme et de la taille cellulaire
Un article de Maciak et Michalak paru en 2014 dans Evolutionary Applications soulignent que « la relation largement négligée que l'on peut constater d'une espèce à l'autre entre la taille cellulaire, le métabolisme cellulaire, et la vitesse de division cellulaire » pourrait être un facteur clef expliquant le paradoxe, concluant que « les grands organismes ont des cellules plus grandes, qui se divisent moins fréquemment et ont un métabolisme moins élevé, facteurs qui tous réduisent la probabilité d'initier un cancer »[38].
Pour Maciak et Michalak, la taille des cellules n'est pas la même chez tous les mammifères, de sorte que la taille corporelle n'est qu'un prédicteur médiocre du nombre de cellules que contient un organisme (par exemple, les globules rouges d'un éléphant sont quatre fois plus gros que ceux d'une musaraigne)[39]. De plus, on observe un rythme de division cellulaire plus rapide sur les petites cellules que sur les grosses ; et l'effet de cette différence dépend exponentiellement de la longévité de l'organisme. Un rythme plus faible de division cellulaire signifie moins d'occasion pour qu'une mutation cancéreuse se manifeste, et les modèles mathématiques de l'incidence des cancers dépendent très fortement de ce rythme[40].
De plus, les organismes plus grands ont généralement un métabolisme de base plus faible, suivant une loi (en inverse du logarithme de la masse) bien connue.
De ce fait, il est normal de s'attendre à un moindre rythme de détérioration cellulaire rapporté à l'unité de masse : ces deux facteurs combinés sont susceptibles d'expliquer en grande partie la paradoxe apparent.
Hypertumeurs
Certains auteurs ont pu suggérer que les tumeurs malignes sont désavantagées dans les organismes plus gros. En particulier, ils ont formulé l'hypothèse d'une compétition entre phénotypes dans la population cancéreuse, où la sélection naturelle tend à favoriser les cellules « tricheuses » agressives qui croissent en formant leur propre tumeur sur la tumeur dont elles sont issues, formant ainsi une « hypertumeur » susceptible d'endommager voire de détruire le néoplasme initial.
Dans un organisme plus gros, il faut plus de temps aux tumeurs pour atteindre une taille létale. Par conséquent, dans les gros organismes, les cancers peuvent être à la fois plus fréquents et en même temps moins létaux[41],[42],[6].
Notes et références
Références
- (en) R. Peto, F. J. C. Roe, P. N. Lee, L. Levy et J. Clack, « Cancer and ageing in mice and men », British Journal of Cancer, vol. 32, no 4, , p. 411–426 (PMID 1212409, PMCID 2024769, DOI 10.1038/bjc.1975.242)
- (en) John D. Nagy, Erin M. Victor et Jenese H. Cropper, « Why don't all whales have cancer? A novel hypothesis resolving Peto's paradox », Integrative and Comparative Biology, vol. 47, no 2, , p. 317–328 (PMID 21672841, DOI 10.1093/icb/icm062)
- F. Galis: Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes, and cancer. In: J Exp Zool 285, 1999, S. 19–26. (Review)
- F. Galis und J. A. J. Metz: Anti-cancer selection as a source of developmental and evolutionary constraints. In: BioEssays 23, 2003, S. 1035–1039.
- A. M Leroi u. a.: Cancer selection. In: Nat Rev Cancer 3, 2003, S. 226–231. (Review)
- J. D. Nagy u. a.: Why don’t all whales have cancer? A novel hypothesis resolving Peto’s paradox. In: Integrative and Comparative Biology 47, 2007, S. 317–328. DOI:10.1093/icb/icm062
- H. B. Andervort et T. B. Dunn: Occurrence of tumors in wild house mice. In: J Natl Cancer Inst 28, 1962, S. 1153–1163.
- J. Morris un J. Dobson: Small Animal Oncology. Blackwell Science, Oxford, 2001. (ISBN 0-632-05282-1)
- R. J. B. King: Cancer Biology. 2. Auflage, Pearson Education, 2000. (ISBN 0-131-29454-7)
- R. B. Landy: Pathology of Zoo Animals. (Editoren: R. J. Montali und G. Migaki), Smithsonian Institution Press, 1980.
- C. J. Uys und P.B. Best: Pathology of lesions observed in whales flensed at Saldanha Bay, South Africa. In: J Comp Pathol 76, 1966, S. 407–412.
- D. Martineau u. a.: Cancer in wildlife, a case study: Beluga from the St. Lawrence Estuary, Quebec, Canada. In: Environ Health Persp 110, 2000, S. 285–292.
- S. De Guise u. a.: Tumors in St. Lawrence beluga whales (Delphinapterus leucas). In: Vet Pathol 31, 1994, S. 444–449.
- S. De Guise u. a.: Possible mechanisms of action of environmental contaminants on St. Lawrence beluga whales (Delphinapterus leucas). In: Environ Health Persp 103, 1995, S. 73–77.
- D. Martineau u. a.: Pathology and toxicology of beluga whales from the St. Lawrence Estuary, Quebec, Canada. Past, present and future. In: Sci Total Environ 154, 1994, S. 201–215. (Review)
- D. C. G. Muir u. a.: Persistent organochlorines in beluga whales (Delphinapterus leucas) from the St. Lawrence River estuary. I. Concentrations and patterns of specific PCBs, chlorinated pesticides and polychlorinated dibenzo-p-dioxins and dibenzofurans. In: Environ Pollut 93, 1996, S. 219–234.
- R. Peto (1977). « Epidemiology, multistage models, and short-term mutagenicity tests » dans Cold Spring Harbor Conferences on Cell Proliferation 4: 1403–1428 p., Cold Spring Harbor Laboratory. Consulté le October 13, 2015.
- Richard Nunney, « The real war on cancer: the evolutionary dynamics of cancer suppression », Evolutionary Applications, vol. 6, no 1, , p. 11–19 (DOI 10.1111/eva.12018)
- (en) Aleah Caulin et Carlo Maley, « Peto’s Paradox: Evolution’s Prescription for Cancer Prevention », Cell Press, vol. 26, no 4, , p. 175–182 (PMID 21296451, PMCID 3060950, DOI 10.1016/j.tree.2011.01.002)
- (en) George Smith et Martin Shipley, « Height and mortality from cancer among men: prospective observational study », BMJ, vol. 317, no 7169, , p. 1351–1352 (PMID 9812932, PMCID 28717, DOI 10.1136/bmj.317.7169.1351)
- (en) « Height and cancer incidence in the Million Women Study: prospective cohort, and meta-analysis of prospective studies of height and total cancer risk », Lancet Oncology, vol. 12, no 8, , p. 785–794 (PMID 21782509, PMCID 3148429, DOI 10.1016/S1470-2045(11)70154-1)
- (en) J.M. Fleming et K.E. Creevy, « Mortality in North American Dogs from 1984 to 2004: An Investigation into Age-, Size-, and Breed-Related Causes of Death », Journal of Veterinary Internal Medicine, vol. 25, no 2, , p. 187–198 (PMID 21352376, DOI 10.1111/j.1939-1676.2011.0695.x, lire en ligne)
- (en) Joshua Schiffman, « Potential Mechanisms for Cancer Resistance in Elephants and Comparative Cellular Response to DNA Damage in Humans », JAMA, vol. 314, , p. 1850 (DOI 10.1001/jama.2015.13134, lire en ligne)
- A. V. Lichtenstein: Cancer as a Programmed Death of an Organism. In: Biochemistry (Moscow) 70, 2005, S. 1055–1064.
- A. V. Lichtenstein: On evolutionary origin of cancer. In: Cancer Cell Int 5, 2005, S. 5. , PMC 555547
- J. Cairns: Mutation selection and the natural history of cancer. In: Nature 255, 1975, S. 197–200.
- A. F. Caulin et C. C. Maley, « Peto's Paradox: Evolution's prescription for cancer prevention », Trends in Ecology & Evolution, vol. 26, no 4, , p. 175–182 (PMID 21296451, PMCID 3060950, DOI 10.1016/j.tree.2011.01.002)
- (en) H Kobayashi et S Man, « Acquired multicellular-mediated resistance to alkylating agents in cancer », Proceedings of the National Academy of Sciences of the United States of America, vol. 90, no 8, , p. 3294–8 (DOI 10.1073/pnas.90.8.3294)
- (en) Tomislav Domazet-Lošo et Diethard Tautz, « Phylostratigraphic tracking of cancer genes suggests a link to the emergence of multicellularity in metazoa », Biomed Central Biology, vol. 8, no 66, (DOI 10.1186/1741-7007-8-66)
- (en) Chi Dang, « Links between metabolism and cancer », Cold Spring Harbor Laboratory Press, vol. 26, , p. 877–90 (PMID 22549953, PMCID 3347786, DOI 10.1101/gad.189365.112)
- (en) Virginia Gewin, « Massive animals may hold secrets of cancer suppression », Nature News, (consulté le 12 mars 2014)
- (en) Carl Zimmer, « Elephants: Large, Long-Living and Less Prone to Cancer », The New York Times, (lire en ligne)
- (en) M. Keane, J. Semeiks, A. E. Webb, Y. I. Li, V. C. Quesada, T. Craig, L. B. Madsen, S. Van Dam, D. Brawand, P. C. I. Marques, P. Michalak, L. Kang, J. Bhak, H. S. Yim, N. V. Grishin, N. H. Nielsen, M. P. Heide-Jørgensen, E. M. Oziolor, C. W. Matson, G. M. Church, G. W. Stuart, J. C. Patton, J. C. George, R. Suydam, K. Larsen, C. López-Otín, M. J. o’Connell, J. W. Bickham, B. Thomsen et J. O. P. De Magalhães, « Insights into the Evolution of Longevity from the Bowhead Whale Genome », Cell Reports, vol. 10, , p. 112–22 (PMID 25565328, DOI 10.1016/j.celrep.2014.12.008)
- (en) T. Xian et J. Azpurua, « INK4 locus of the tumor-resistant rodent, the naked mole rat, expresses a functional p15/p16 hybrid isoform. », Proceedings of the National Academy of Sciences, vol. 112, no 4, , p. 1053–8 (PMID 25550505, PMCID 4313802, DOI 10.1073/pnas.1418203112)
- (en) E. Callaway, « How elephants avoid cancer: Pachyderms have extra copies of a key tumour-fighting gene. », Nature, vol. 526, (DOI 10.1038/nature.2015.18534)
- (en) Vincent Lynch, « TP53 copy number expansion correlates with the evolution of increased body size and an enhanced DNA damage response in elephants », bioRxiv (preprint), (DOI 10.1101/028522, lire en ligne)
- (en) Stuart D. Tyner et Sundaresan Venkatachalam, « p53 mutant mice that display early ageing-associated phenotypes », Nature, vol. 415, , p. 45–53 (PMID 11780111, DOI 10.1038/415045a, lire en ligne)
- (en) S. MacIak et P. Michalak, « Cell size and cancer: A new solution to Peto's paradox? », Evolutionary Applications, vol. 8, , p. 2–8 (DOI 10.1111/eva.12228)
- (en) T. Ryan Gregory, « Mammal erythrocyte sizes », sur Genome Size, (consulté le 13 octobre 2015)
- (en) Peter Calabrese et Darryl Shibata, « A simple algebraic cancer equation: calculating how cancers may arise with normal mutation rates », BMC Cancer, vol. 10, no 3, (DOI 10.1186/1471-2407-10-3, lire en ligne)
- http://m.icb.oxfordjournals.org/content/47/2/317.full
- J. D. Nagy: Competition and natural selection in a mathematical model of cancer. In: Bull Math Biol 66, 2004, S. 663–687.
Bibliographie
- (en) Barbara Natterson-Horowitz et Kathryn Bowers, Zoobiquity: What Animals Can Teach Us about Health and the Science of Healing, Alfred A. Knopf, (ISBN 978-0-307-95838-9)
Voir aussi
- Oncogène
- Gène suppresseur de tumeurs
- Théorie métabolique de l'écologie

[[Catégorie:Paradoxe en biologie[Peto]]