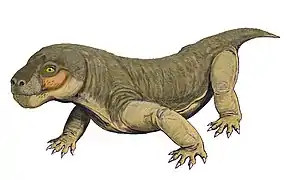
Euchambersia
Euchambersia est un genre éteint de thérapside thérocéphales ayant vécu au Permien tardif, il y a environ 255 millions d'années, dans ce qui est aujourd'hui l'Afrique du Sud et dont une seule espèce est connu : Euchambersia mirabilis, nommée par le paléontologue Robert Broom en 1931 à partir d'un crâne manquant aux mâchoires inférieures; un deuxième crâne, appartenant à un individu immature, a ensuite été décrit. C'est un membre de la famille des Akidnognathidae.
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Synapsida |
| Ordre | Therapsida |
| Sous-ordre | † Therocephalia |
| Infra-ordre | † Eutherocephalia |
| Famille | † Akidnognathidae |

Euchambersia était un petit thérocephale possédant de grandes canines. Cependant, il est notable chez les thérocephales pour posséder des crêtes sur ses canines et une grande indentation sur le côté du crâne. Dans l'hypothèse erronée que les canines sont rainurées au lieu d'être striées, il a été proposé que ces structures supportent un mécanisme de distribution de venin. Plus récemment, la structure interne du crâne d'Euchambersia a été utilisée comme preuve plus forte en faveur de l'hypothèse qu'il était venimeux; d'autres possibilités, comme l'indentation soutenant une sorte d'organe sensoriel, restent encore plausibles.
Description
Taille et morphologie

Euchambersia était de petite taille et un museau relativement court pour un thérocephale, le type de crâne ayant une longueur reconstruite d'environ 116 millimètres, expliquant l'écrasement et la déformation du fossile. Le deuxième crâne connu appartenait à un individu plus petit, d'une longueur de 80 millimètres qui était probablement immature, à en juger par le manque de fusion dans le crâne[1].
Selon la description initiale, l'orbite d'Euchambersia était plutôt petit. Les branches des Os postorbitaire et du jugal qui entourent habituellement l'arrière et le bas de l'orbite oculaire chez les thérocephales semblent être soit très réduites, soit totalement absentes. Pendant ce temps, le haut de l'orbite est formé par l'os préfrontal et l'os frontal est également petit. Le crâne ne porte pas de foramen pinéale. Comme Whaitsia, le ptérygoïde et le palatin du palais ne sont pas séparés du transpalatin, plus loin sur le côté de la mâchoire, par aucune sorte d'ouverture[2].
Venin
Les grandes fosses maxillaires d'Euchambersia ont fait l'objet de débats continus concernant leur fonction. Cependant, la plupart des chercheurs conviennent qu'ils détenaient une sorte de glande sécrétoire. Alors que Broom a initialement soutenu que les fosses pouvaient contenir les glandes salivaires parotides[2], cette proposition a été rejetée par Boonstra et JP Lehman, qui ont noté que les glandes parotides ont tendance à être placées derrière l'œil; ils ont suggéré respectivement que les fosses contenaient des glandes lacrymales modifiées et des glandes de Harderian[1]. Cependant, ce dernier est également peu probable car les glandes de Harder sont généralement placées à l'intérieur de l'orbite. Franz Nopcsa a suggéré que les fosses maxillaires abritaient des glandes à venin (qui peuvent être dérivées de glandes lacrymales), les canines striées et les encoches derrière les canines permettant au venin de s'écouler passivement dans la circulation sanguine de la victime[3]. Cette hypothèse a été largement acceptée tout au long du 20e siècle[4],[5],[6],[7] et la morphologie caractéristique d'Euchambersia a été utilisée pour soutenir d'éventuelles adaptations portant du venin parmi divers autres animaux préhistoriques[8],[9],[10],[11], y compris le thérocéphale Ichibengops[12].
Une grande partie de cette acceptation a été basée sur l'hypothèse erronée que les canines sont rainurées au lieu d'être striées, des canines rainurées d'Euchambersia seraient parallèles aux crocs de divers serpents venimeux ainsi qu'aux incisives libérant du venin des solénodons vivants[7]. Cette interprétation, qui est constamment apparue dans la littérature publiée après 1986, a été déterminée par Julien Benoit comme étant le résultat de la propagation du diagramme trop reconstruit du crâne de Broom, sans le contexte des spécimens réels. Cette ligne de preuve a été soulevée pour soutenir la nécessité d'une réévaluation de l'hypothèse d'une morsure venimeuse d'Euchambersia[13]. De plus, Benoit a fait valoir que les canines cannelées et striées ne sont pas nécessairement associées aux animaux venimeux non plus, comme le montre leur présence dans les hippopotames, les muntjacs et les babouins, dans lesquels ils jouent un rôle dans le toilettage ou l'aiguisage des dents[14], dans les deux derniers, les canines striées sont également accompagnées d'une fosse distincte devant l'œil, qui n'a aucun lien avec le venin[15]. De plus, les dents rainurées et striées des serpents non venimeux sont utilisées pour réduire la traînée d'aspiration lors de la capture de proies glissantes comme les poissons ou les invertébrés[16].

La tomodensitométrie des spécimens connus d'Euchambersia par Benoit et ses collègues a ensuite été utilisée pour fournir un support plus concret en faveur de l'hypothèse du venin. Les canaux menant vers et depuis les fosses maxillaires, comme le révèlent les analyses, auraient principalement soutenu le nerf trijumeau ainsi que les vaisseaux sanguins[17],[18],[19]. Cependant, le fait que les canaux conduisent également directement à la racine des canines suggère qu'ils avaient un rôle secondaire dans l'apport de venin. En tout, Euchambersia semble avoir eu une glande à venin (logée dans les fosses maxillaires), un mécanisme de délivrance du venin (les canaux maxillaires) et un instrument par lequel une blessure pour la livraison de venin peut être infligée (les canines striées), qui répondent aux critères d'un animal venimeux tels que définis par Wolfgang Bücherl[20]. Benoit et al. a noté que cela ne démontre pas de manière concluante qu'Euchambersia était réellement venimeux, en particulier compte tenu des objections précédemment énoncées. De plus, il n'y a pas d'animaux vivants avec un système de distribution analogue au système proposé pour Euchambersia (la plupart délivrent du venin par la mâchoire inférieure[21],[22] tandis que les serpents ont des canaux spécialisés[1],[23]).
Une autre hypothèse suggérée par Benoit et al. implique une sorte d'organe sensoriel occupant la fosse maxillaire. Uniquement parmi les thérapsides[24] le canal à l'intérieur du maxillaire est exposé à l'arrière de la fosse maxillaire, ce qui implique que le canal, portant le nerf trijumeau, se serait probablement étendu à travers la fosse, en dehors du contour du crâne. Benoit et al. a émis l'hypothèse que la fosse pouvait avoir soutenu un organe sensoriel spécialisé analogue à l'organe de fosse des vipères à fosse et d'autres serpents[25], ou alternativement un ganglion de cellules nerveuses. Il est également possible que cet organe remplaçait l'œil pariétal en Euchambersia, comme le fait l'organe de la fosse dans les vipères de la fosse[26] Cependant, un tel organe sensoriel élargi serait sans précédent chez les tétrapodes, et les quelques autres thérocephales qui manquent également d'un œil pariétal n'ont pas non plus de fosse maxillaire[27]. Ainsi, Benoit et al. considéré l'hypothèse du venin comme étant plus plausible[1].
Références
- Benoit, Norton, Manger et Rubidge, « Reappraisal of the envenoming capacity of Euchambersia mirabilis (Therapsida, Therocephalia) using μCT-scanning techniques », PLoS ONE, vol. 12, no 2, , e0172047 (PMID 28187210, PMCID 5302418, DOI 10.1371/journal.pone.0172047, Bibcode 2017PLoSO..1272047B)
- Broom, « Notices of some new genera and species of Karroo fossil reptiles », Records of the Albany Museum, vol. 4, no 1, , p. 161–166
- Nopcsa, « On the biology of the theromorphous reptile Euchambersia », Annals and Magazine of Natural History, 10e série, vol. 12, no 67, , p. 125–126 (DOI 10.1080/00222933308673757)
- Rubidge et Sidor, « Evolutionary Patterns Among Permo-Triassic Therapsids », Annual Review of Ecology and Systematics, vol. 32, , p. 449–480 (DOI 10.1146/annurev.ecolsys.32.081501.114113)
- Watson et Romer, « A classification of therapsid reptiles », Bulletin of the Museum of Comparative Zoology, vol. 114, , p. 35–89 (lire en ligne)
- Van Valen, « Therapsids as Mammals », Evolution, vol. 14, no 3, , p. 304–313 (DOI 10.2307/2405973, JSTOR 2405973)
- Folinsbee, Muller et Reisz, « Canine Grooves: Morphology, Function, and Relevance to Venom », Journal of Vertebrate Paleontology, vol. 27, no 2, , p. 547–551 (DOI 10.1671/0272-4634(2007)27[547:cgmfar]2.0.co;2, JSTOR 30126324)
- Sues, « Venom-conducting teeth in a Triassic reptile », Nature, vol. 351, no 6322, , p. 141–143 (DOI 10.1038/351141a0, Bibcode 1991Natur.351..141S)
- Ivakhnenko, « The First Whaitsiid (Therocephalia, Theromorpha) », Paleontological Journal, vol. 42, no 4, , p. 409–413 (DOI 10.1134/S0031030108040102)
- Sues, « A reptilian tooth with apparent venom canals from the Chinle Group (Upper Triassic) of Arizona », Journal of Vertebrate Paleontology, vol. 16, no 3, , p. 571–572 (DOI 10.1080/02724634.1996.10011340)
- Gong, Martin, Burnham et Falk, « The birdlike raptor Sinornithosaurus was venomous », Proceedings of the National Academy of Sciences of the United States of America, vol. 107, no 2, , p. 766–768 (PMID 20080749, PMCID 2818910, DOI 10.1073/pnas.0912360107, Bibcode 2010PNAS..107..766G)
- Huttenlocker, Sidor et Angielczyk, « A new eutherocephalian (Therapsida, Therocephalia) from the upper Permian Madumabisa Mudstone Formation (Luangwa Basin) of Zambia », Journal of Vertebrate Paleontology, vol. 35, no 5, , e969400 (DOI 10.1080/02724634.2015.969400)
- Benoit, « A review of the "venomous therocephalian" hypothesis and how multiple re-portrayals of Euchambersia have influenced its success and vice versa », Bulletin de la Société Géologique de France, vol. 187, no 4, , p. 217–224 (DOI 10.2113/gssgfbull.187.4-5.217, lire en ligne)
- Mitchell, Heckert et Sues, « Grooves to tubes: evolution of the venom delivery system in a Late Triassic "reptile" », Naturwissenschaften, vol. 97, no 12, , p. 1117–1121 (PMID 21060984, DOI 10.1007/s00114-010-0729-0, Bibcode 2010NW.....97.1117M)
- Orr, Delezene, Scott et Tocheri, « The comparative method and the inference of venom-delivery systems in fossil mammals », Journal of Vertebrate Paleontology, vol. 27, no 2, , p. 541–546 (DOI 10.1671/0272-4634(2007)27[541:TCMATI]2.0.CO;2)
- Vaeth, Rossman et Shoop, « Observations of Tooth Surface Morphology in Snakes », Journal of Herpetology, vol. 19, no 1, , p. 20–26 (DOI 10.2307/1564416, JSTOR 1564416)
- Bellairs, « Observations on the snout of Varanus, and a comparison with that of other lizards and snakes », Journal of Anatomy, vol. 83, no 2, , p. 116–146 (PMID 17105074, PMCID 1273152)
- Abdel-Kader, Ali et Ibrahim, « The Cranial Nerves of Mabuya quinquetaeniata III: Nervus Trigeminus », Life Science Journal, vol. 8, no 4, , p. 650–669 (lire en ligne)
- Leitch et Catania, « Structure, innervation and response properties of integumentary sensory organs in crocodilians », Journal of Experimental Biology, vol. 215, no 23, , p. 4217–4230 (PMID 23136155, PMCID 4074209, DOI 10.1242/jeb.076836)
- W. Bücherl, Venomous Animals and their Venoms, vol. 1, New York, Academic Press, , 9–12 p. (ISBN 9781483229492, DOI 10.1016/B978-1-4832-2949-2.50006-0), « Introduction »
- Fry, Wroe, Teeuwisse et van Osch, « A central role for venom in predation by Varanus komodoensis (Komodo Dragon) and the extinct giant Varanus (Megalania) priscus », Proceedings of the National Academy of Sciences of the United States of America, vol. 106, no 22, , p. 8969–8974 (PMID 19451641, PMCID 2690028, DOI 10.1073/pnas.0810883106, Bibcode 2009PNAS..106.8969F)
- Ligabue-Braun, Verli et Carlini, « Venomous mammals: A review », Toxicon, vol. 59, no 7, , p. 680–695 (PMID 22410495, DOI 10.1016/j.toxicon.2012.02.012)
- S.A. Weinstein, T.L. Smith et K.V. Kardong, Handbook of Venoms and Toxins of Reptiles, Boca Raton, CRC Press, , 65–91 p., « Reptile Venom Glands: Form, Function, and Future »
- Benoit, Manger et Rubidge, « Palaeoneurological clues to the evolution of defining mammalian soft tissue traits », Scientific Reports, vol. 6, , p. 25604 (PMID 27157809, PMCID 4860582, DOI 10.1038/srep25604, Bibcode 2016NatSR...625604B)
- Goris, « Infrared Organs of Snakes: An Integral Part of Vision », Journal of Herpetology, vol. 45, no 1, , p. 2–14 (DOI 10.1670/10-238.1)
- Krochmal, Bakken et LaDuc, « Heat in evolution's kitchen: evolutionary perspectives on the functions and origin of the facial pit of pitvipers (Viperidae: Crotalinae) », Journal of Experimental Biology, vol. 207, no 24, , p. 4231–4238 (PMID 15531644, DOI 10.1242/jeb.01278)
- Benoit, Abdala, Manger et Rubidge, « The Sixth Sense in Mammalian Forerunners: Variability of the Parietal Foramen and the Evolution of the Pineal Eye in South African Permo-Triassic Eutheriodont Therapsids », Acta Palaeontologica Polonica, vol. 61, no 4, , p. 777–789 (DOI 10.4202/app.00219.2015)
 Portail de la paléontologie
Portail de la paléontologie  Portail de l’Afrique du Sud
Portail de l’Afrique du Sud