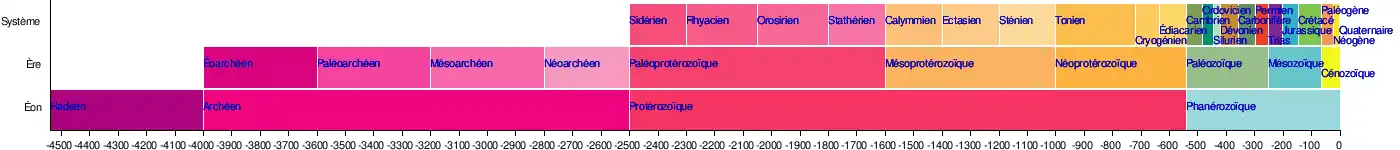
Cryogénien
Le Cryogénien est la deuxième période du Néoprotérozoïque. Elle s'étend de −720 à −635 Ma[1]. Elle suit le Tonien et précède l'Édiacarien. Elle est parfois subdivisée en deux époques : le Sturtien (de −720 à −650 Ma) et le Varangien (de −650 à −635 Ma), des noms de deux des âges glaciaires survenus lors de cette période.
| Notation chronostratigraphique | NP2 |
|---|---|
| Notation RGF | a3b |
| Niveau | Période / Système |
| Érathème / Ère - Éonothème / Éon -- Superéon |
Néoprotérozoïque Protérozoïque Précambrien |
Stratigraphie
| Début | Fin |
|---|---|
| ≃720 Ma |
Cette période a été baptisée Cryogénien parce qu'ayant connu une glaciation générale de la Terre : la glaciation Varanger (le préfixe cryo- signifie « froid »).
Paléobiologie
Les premières traces de vie multicellulaires découvertes datent d'environ −2,1 Ga (fossiles du bassin sédimentaires de Franceville)[2], mais il a fallu un certain temps pour voir apparaitre les premiers animaux, plus complexes.
Des travaux récents[3] laissent penser que cette époque, en dépit de conditions de vie apparemment difficiles, a coïncidé avec une hausse des populations d'algues dans les océans cryogéniques et avec l'émergence de premières formes de vies animales (encore très primitives, avec les premières éponges notamment)[4]. Elles seront localement si abondantes et durant une période si longue que leurs spicules est le principal constituant de certaines roches (Spiculites[5],[6],[7], gaizes et spongolites[8]).
C'est donc une période de transition entre un monde vivant bactérien, puis algo-bactérien en partie photosynthétique[9],[10] vers un monde aussi colonisé par des eucaryotes[3]. La période de 800 à 717 Ma est en effet caractérisée par une croissance de la diversité des microfossiles eucaryotes.
Ceci correspond à l'un des premiers grands bouleversements écologiques, l'un des plus profonds que la vie sur Terre ait connus : avec une totale réorganisation de la distribution du cycle du carbone et des nutriments (phosphore notamment[11],[12]) dans la colonne d'eau et l'augmentation du flux d'énergie vers des niveaux trophiques plus nombreux et complexes[3].
Les progrès de la biogéochimie ont permis l'étude rétrospective de certains biomarqueurs eucaryotes tels que le colestane, l'ergostane, le stigmastane, le dinostane, l'isopropylcholestane, le n-propylcholestane et le cryostane[13]. Elle a confirmé l'importance de cette période géologique pour la radiation évolutive[13]. Et elle apporte des précisions sur la géographie évolutive des premiers eurcaryotes ainsi que sur l'enrichissement taxonomique de ce groupe lors de la grande glaciation de la Terre et jusqu'à l'Édiacarien (635-541 Ma)[13].
Le 26-méthylsterol pourrait avoir protégé des éponges, mais aussi d'autres eucaryotes, contre leurs propres toxines membranyliques (divers protistes sécrètent des toxines lytiques (c'est-à-dire capable de dissoudre (lyser) des parois cellulaires), ce qui leur permet à la fois d'échapper à la prédation et de parasiter ou tuer des proies eucaryotes[13].
Comme les membranes cellulaires construites avec des stérols peuvent être la cible de telles attaques, l'abondance du cryostane plaide en faveur d'une prédation dans le groupe Chuar et plaide pour l'hypothèse d'une eucaryophagie qui se serait généralisée au Cryogénien[13].
Le bilan fossile moléculaire des stéroïdes eucaryotes[14] incite à penser que la biomasse bactérienne était initialement encore la seule source primaires trophique importante dans les océans cryogéniens. Ces stéroïdes se sont ensuite diversifiés et sont devenus très abondants, ce qui est le signe de l'accroissement rapide de la biomasse d'algues planctoniques marines (Archaeplastida), dans un intervalle de temps géologiquement très court, entre les glaciations Sturtian et Marinoan « La Terre boule de neige », il y a 659-645 Ma[3]. Il apporte aussi des indices sur la vie et l'apparence des éponges qui semblent compter parmi les premiers organismes animaux coloniaux importants[15].
Certains géobiologistes pensent que c'est le moment du développement des cyanobactéries[3].
L'explosion du plancton algual et cyanobactérien a donné naissance à de nouvelles chaines alimentaires, et surtout à des transferts d'éléments nutritifs et énergétiques plus efficaces et complexes[16], permettant l'élaboration d'écosystèmes de plus en plus complexes et stables comme le montre l'apparition conjointe de biomarqueurs pour les éponges (qui semblent avoir dû s'adapter à des protistes toxiques[13]) et les rhizariens prédateurs, et le rayonnement subséquent des eumétazoaires dans la période de l'Édiacarien[4].
Les temps géologiques précis de cette transition restent encore à affiner, de même que les liens possibles avec d'une part la hausse du taux d'oxygène atmosphérique[17],[3] (et donc l'apparition d'une couche d'ozone protectrice), et d'autre part avec le début de l'évolution animale[3].
Ampleur de la glaciation
Certains scientifiques estiment que cette glaciation très sévère a concerné la planète entière ; d'autres estiment que la bande équatoriale a été épargnée par la glace.
Paléogéographie
À l'époque du Cryogénien, le supercontinent de Rodinia commence à se fragmenter, un rift séparant le continent en deux grandes masses. C'est cette fragmentation qui serait à l'origine de la glaciation[18]. En effet, le cœur d'un supercontinent est généralement loin de l'influence océanique et donc désertique. Sa fragmentation rend de nouvelles surfaces accessible à l'érosion et au transport : l'érosion des silicates exposés entraîne la capture de CO2 pour former des bicarbonates solubles, et leur arrivée dans l'océan les fait précipiter sous forme de carbonates, fixant ainsi le carbone. La fragmentation d'un supercontinent est donc toujours suivi d'une chute dans l'effet de serre du gaz carbonique, entraînant un refroidissement global.
Notes et références
- Selon l'ICS .
- Sean Bailly, « Les plus anciens organismes pluricellulaires », sur Pourlascience.fr (consulté le )
- (en) Jochen J. Brocks, Amber J. M. Jarrett, Eva Sirantoine, Yosuke Hoshino et Tharika Liyanage, « The rise of algae in Cryogenian oceans and the emergence of animals », Nature, (DOI 10.1038/nature23457, lire en ligne, consulté le )
- (en) N.J. Butterfield, « Macroevolutionary turnover through the Ediacaran transition: ecological and biogeochemical implications », Geol. Soc. Spec. Publ, vol. 326, , p. 55-66.
- Gammon, P. R., James, N. P., & Pisera, A. (2000). Eocene spiculites and spongolites in southwestern Australia: not deep, not polar, but shallow and warm. Geology, 28(9), 855-858
- Cavaroc, V. V., & Ferm, J. C. (1968). Siliceous spiculites as shoreline indicators in deltaic sequences. Geological Society of America Bulletin, 79(2), 263-272.
- Schindler, T., Wuttke, M., & Poschmann, M. (2008). Oldest record of freshwater sponges (Porifera: Spongillina)—spiculite finds in the Permo-Carboniferous of Europe. Paläontologische Zeitschrift, 82(4), 373-384.
- Gammon P.R. (1978). Spiculites and spongolites. In Sedimentology (pp. 1130-1134). Springer Netherlands.
- (en) P. Sánchez-Baracaldo, J.A. Raven, D. Pisani et A.H. Knoll, « Early photosynthetic eukaryotes inhabited salinity habitats », Proc. Natl Acad. Sci. USA, .
- (en) N.J. Butterfield, « Proterozoic photosynthesis – a critical review », Palaeontology, vol. 58, , p. 953–972.
- .
- (en) C.T. Reinhard et al., « Evolution of the global phosphorus cycle », Nature, vol. 541, , p. 386–389
- (en) J.J. Brocks et al., « Early sponges and toxic protists : possible sources of cryostane, an age diagnostic biomarker antedating Sturtian Snowball Earth », Geobiology, vol. 14, , p. 129-149 (résumé).
- (en) G.D. Love et al., « Fossil steroids record the appearance of Demospongiae during the Cryogenian period », Nature, vol. 457, , p. 718-721
- (en) G.D. Love, E. Grosjean, C. Stalvies, D.A. Fike, J.P. Grotzinger, A.S. Bradley, A.E. Kelly, M. Bhatia, W. Meredith, C.E. Snape et al., « Fossil steroids record the appearance of Demospongiae during the Cryogenian period », Nature, vol. 457, no 7230, , p. 718-721.
- (en) A.J. Irwin, Z.V. Finkel, O.M.E. Schofield et P.G. Falkowski, « Scaling-up from nutrient physiology to the size-structure of phytoplankton communities », J. Plankton Res., vol. 28, , p. 459–471.
- (en) Torsvik, « The Rodinia Jigsaw Puzzle », Sciences, vol. 300, no 5624, , p. 1379-1381 (lire en ligne)
{kind=link}