Vie à l'époque archéenne dans la ceinture de Barberton
La ceinture de Barberton[1], dans l'est de l'Afrique du Sud, est une formation géologique qui recèle quelques-uns des plus anciens fossiles témoignant sans conteste de l'existence de la vie à l'époque archéenne. Ces fossiles, de la taille d'une cellule procaryote, ont été trouvés dans des roches vieilles de 3,5 Ga[2]. C'est un excellent endroit pour étudier la vie à l’époque archéenne car des sédiments et des roches métasédimentaires sont exposés.

Étudier les formes de vie les plus anciennes sur notre planète peut apporter des informations précieuses aidant à comprendre comme la vie pourrait avoir évolué sur d'autres planètes. On a ainsi longtemps pensé que la vie pouvait exister sur Mars car l'environnement et la tectonique y sont similaires à ceux de la période archéenne sur Terre[3]. En connaissant l’environnement dans lequel la vie a évolué sur Terre et la façon dont les roches en préservent les traces, les scientifiques peuvent mieux savoir où chercher les traces de vie sur Mars.
Les débuts de la vie sur Terre
Une vie fossile datée de 3,5 Ga a aussi été trouvée dans le craton de Pilbara dans l'ouest australien[4]. Cela, ainsi que les fossiles de Barberton, montrent que la vie cellulaire a existé à ce stade de l'évolution de la Terre. D'autres travaux montrent qu'il y aurait peut-être eu de la vie il y a 3,8 milliards d'années dans ce qui est de nos jours l'ouest du Groenland[5],[6] mais cela est toujours sujet à d'intenses débats. La vie cellulaire existait il y a 3,5 milliards d'années et avait donc commencé à évoluer avant cela. La Terre étant âgée de 4,5 milliards d'années[7], il y eut donc une « fenêtre » d'un milliard d'années pour permettre à la vie d'apparaître à partir d'une Terre qui en était dépourvue à l'origine.
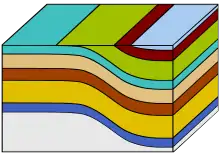
Géologie
Histoire tectonique de la ceinture de Barberton
La ceinture de diorite de Barberton se situe dans le craton du Kaapvaal, qui sous-tend une grande partie de l'Afrique australe ; elle s'est formée par la mise en place de batholites granitoïdes[8]. Le craton du Kaapvaal a fait partie d'un supercontinent appelé Vaalbara, qui comprenait aussi le craton de Pilbara, situé aujourd'hui dans l'ouest australien[8]. Bien que la date exacte soit toujours débattue, il est probable que le Vaalbara ait existé de 3,6 à 2,2 milliards d'années avant le présent, puis qu'il se soit divisé en deux continents différents[9].
Preuves de vie
Les traces de vie conservées dans les roches archéennes ont été altérées par leur histoire de 3,5 milliards d'années et, donc, sont difficiles à discerner. Les parois cellulaires peuvent être préservées mais leur composition change avec le temps et elles se minéralisent. Six critères ont été établis pour permettre de déterminer si une microstructure donnée est susceptible d'être un microfossile[10],[11] :
- Les vrais microfossiles doivent être relativement abondants ;
- Les vrais microfossiles doivent être composés de carbone, ou, s'ils ont été minéralisés, correspondre à un précipité biologique (par exemple, certaines bactéries forment des pyrites dans le cadre de leur processus métabolique[12]) ;
- Les vrais microfossiles doivent présenter une morphologie biologique ;
- Les vrais microfossiles doivent apparaître dans un contexte biologiquement plausible (par exemple, il ne peut y avoir de microfossiles dans une roche magmatique car la vie ne peut exister dans la lave en fusion) ;
- Les vrais microfossiles doivent rentrer dans un contexte évolutionnaire bien établi (par exemple, l'existence de microfossiles complexes il y a 3,5 milliards d'années est hautement improbable car ils doivent avoir eu le temps d'évoluer depuis les cellules simples, leurs ancêtres) ;
- Les vrais microfossiles doivent être différents de la matière carbonée n'ayant pas une origine biologique. Cela s’établit par analyse isotopique.
Morphologie des cellules

Les cellules biologiques sont préservées dans la roche car les parois cellulaires sont faites de protéines qui se transforment en kérogène à la mort de la cellule. Le kérogène est insoluble dans les minéraux acides aussi bien que dans les matériaux basiques et les solvants organiques[13]. Au fil du temps, il se minéralise en graphite ou en carbone de type graphite ou bien se dégrade en hydrocarbures de pétrole et de gaz[14].
Il existe trois types principaux de morphologie cellulaire. Bien qu'il n'y ait pas d'échelle de taille pour chacun d'eux, les microfossiles sphéroïdes peuvent ne faire que 8 μm, les filamenteux ont un diamètre typique inférieur à 5 μm et une longueur allant de 10 à 100 μm et ceux en forme de broche peuvent atteindre 50 μm[2],[15].
Analyse isotopique
Le fractionnement isotopique est un bon moyen pour caractériser et donc distinguer le carbone organique de l'inorganique. On utilise l'indicateur δ13C (delta treize C), ratio des isotopes stables du carbone, 13C / 12C, exprimé en « pour mille » (‰).
L'analyse isotopique du carbone inorganique donne des valeurs pour δ13C allant jusqu'à -10 ‰, avec une fourchette typique variant de -5 à 5 ‰. Le carbone organique, quant à lui, voit le δ13C varier de -20 ‰, pour les bactéries phototropes[16] à -60 ‰ pour les communautés microbiennes utilisant le méthane[17]. Ce large écart pour le carbone organique est lié au métabolisme cellulaire ; un organisme utilisant la photosynthèse (un phototrophe) aura une valeur δ13C différente d'un organisme qui dépend de substances chimiques pour son énergie (un autotrophe).
Conservation des fossiles
Les plus anciens microfossiles de la ceinture de Barberton se trouvent dans le groupe d'Onverwacht, plus précisément dans les formations de Kromberg et d'Hooggenoeg[2]. Ces deux formations sont constituées principalement de roches magmatiques ; les roches sédimentaires sont devenues métamorphiques. Il est pourtant possible de trouver des microfossiles dans les chailles, un type d'évaporite qui se forme dans un environnement sédimentaire. D'après les trouvailles faites dans les chailles et dans les stromatolithes lithifiés, il est probable que la vie ancienne a existé sous forme de « tapis microbiens » et de stromatolithes[2].
Les stromatolithes sont des marqueurs de l'existence de larges colonies de micro-organismes ; on les trouve dans les roches fossiles et, rarement, dans les environnements hypersalins modernes. Un stromatolithe typique est fait d'une alternance de couches de sédiments et de microbes ; les microbes utilisant la photosynthèse, les stromatolithes sont caractéristiques d'environnements aqueux de faible profondeur du fait que les microbes nécessitent d'être dans la zone photique des cours d'eau. Les stromatolithes sont typiquement constitués de microfossiles filamenteux[18]. Les plus anciens stromatolithes ont été datés de 3,5 Ga[19] ; c'est aussi l'âge de ceux de la ceinture de Barberton.
Les microfossiles trouvés dans les chailles datent aussi de 3,5 Ga. Les trois types de morphologie ont été trouvés. Les chailles peuvent avoir diverses couleurs, mais les microfossiles se trouvent principalement dans les chailles noires, cette couleur pourrait donc être l'indicateur d'un matériau organique[2].
Applications futures
Les scientifiques ont établi la date approximative à laquelle apparaissent les premiers fossiles, mais elle ne correspond pas nécessairement à celle à laquelle la vie a commencé à évoluer. Aucun fossile n'a été retrouvé dans des roches plus anciennes, mais des preuves de vie peuvent être trouvées par d'autres voies, comme par exemple les données étendues concernant les isotopes du carbone et la spectroscopie Raman. D'autres travaux sont menés sur le sujet de l'évolution de la vie cellulaire sur la Terre hostile des premiers âges.
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Archean life in the Barberton Greenstone Belt » (voir la liste des auteurs).
- Ou « ceinture de roches vertes de Barberton », ou bien encore « ceinture de diorite de Barberton ».
- (en) M. Walsh, « Microfossils and possible microfossils from the early Archean Onverwacht Group, Barberton mountain land, South Africa », Precambrian Research, vol. 54, , p. 271–293 (PMID 11540926, DOI 10.1016/0301-9268(92)90074-X)
- (en) F. Westall, A. Steele, J. Toporski, M. Walsh, C. Allen, S. Guidry, McKay, Gibson et H. Chafetz, « Extracellular polymeric substances as biomarkers in terrestrial and extraterrestrial materials », Journal of Geophysical Research, vol. 105, no 10, , p. 24511–24527 (DOI 10.1029/2000JE001250, Bibcode 2000JGR...10524511W).
- (en) J.W. Schopf, « Fossil evidence of Archaean life », Philosophical Transactions of the Royal Society, b, vol. 361, , p. 869–885 (PMID 16754604, PMCID 1578735, DOI 10.1098/rstb.2006.1834)
- (en) S.J. Mojzsis, G. Arrhenius, K.D. Keegan, T.H. Harrison, A. P. Nutman et C.L.R. Friend, « Evidence for life on earth 3,800 million years ago », Nature, vol. 384, no 6604, , p. 55–58 (PMID 8900275, DOI 10.1038/384055a0, Bibcode 1996Natur.384...55M)
- (en) K. D. McKeegan, A. B. Kudryavtsev et J.W. Schopf, « Raman and ion microscopic imagery of graphitic inclusions in apatite from older than 3830 Ma Akilia supracrustal rocks, west Greenland », Geology, vol. 35, no 7, , p. 591–594 (DOI 10.1130/G23465A.1, Bibcode 2007Geo....35..591M)
- (en) C. Patterson, « Age of meteorites and the Earth », Geochimica et Cosmochimica Acta, vol. 10, , p. 230–237 (DOI 10.1016/0016-7037(56)90036-9, Bibcode 1956GeCoA..10..230P, lire en ligne)
- (en) E.S. Cheney, « Sequence stratigraphy and plate tectonic significance of the Transvaal succession of southern Africa and its equivalent in Western Australia », Precambrian Research, vol. 79, , p. 3–24 (DOI 10.1016/0301-9268(95)00085-2)
- (en) T.E. Zegers, M. de Wit, J. Dann et S.H. White, « Vaalbara, Earth's oldest assembled continent? A combined structural, geochronological, and palaomagnetic test », Terra Nova, vol. 10, , p. 250–259 (DOI 10.1046/j.1365-3121.1998.00199.x)
- (en) J.W. Schopf et M. R. Walter, « Archean microfossils: new evidence of ancient microbes », dans J. W. Schopf, Earth's Earliest Biosphere, New Jersey, Princeton University Press, , p. 214–239
- (en) R. Buick, « Carbonaceous filaments from North Pole, Western Australia: are they fossil bacteria in Archaean stromatolites? », Precambrian Research, vol. 24, , p. 157–172 (DOI 10.1016/0301-9268(84)90056-1, lire en ligne [PDF])
- (en) H. Ohmoto, T. Kakegawa et D.R. Lowe, « 3.4-Billion-year-old biogenic pyrites from Barberton, South Africa: sulfur isotope evidence », Science, vol. 262, , p. 555–557 (DOI 10.1126/science.11539502, Bibcode 1993Sci...262..555O)
- (en) R.P. Philp et M. Calvin, « Possible origin for insoluble organic (kerogen) debris in sediments from insoluble cell-wall materials of algae and bacteria », Nature, vol. 262, no 5564, , p. 134–136 (DOI 10.1038/262134a0, Bibcode 1976Natur.262..134P).
- (en) E.W. Tegelaar, J. W. deLeeuw, S. Derenne et C. Largeau, « A reappraisal of kerogen formation », Geochimica et Cosmochimica Acta, vol. 53, no 11, , p. 3103–3106 (DOI 10.1016/0016-7037(89)90191-9, Bibcode 1989GeCoA..53.3103T)
- (en) D. Z. Oehler, F. Robert, S. Mostefaoui, A. Meibom, M. Selo et D.S. McKay, « Chemical Mapping of Proterozoic Organic Matter at Submicron Spatial Resolution », Astrobiology, vol. 6, no 6, , p. 838–850 (PMID 17155884, DOI 10.1089/ast.2006.6.838, Bibcode 2006AsBio...6..838O)
- (en) M. Schidlowski, J.M. Hayes, I.R. Kaplan (éd.) et J.W. Schopf, Earth's Early Biosphere, Princeton University Press, , p. 149–186
- (en) M. Schidlowski, « A 3,800-million-year isotope record of life from carbon in sedimentary rocks », Nature, vol. 333, no 6171, , p. 313–318 (DOI 10.1038/333313a0, Bibcode 1988Natur.333..313S)
- (en) G.R. Byerly, D.R. Lowe et M. Walsh, « Stromatolites from the 3,300 to 3,500-myr Swaziland Supergroup, Barberton Mountain Land, South Africa », Nature, vol. 319, no 6053, , p. 489–491 (DOI 10.1038/319489a0, Bibcode 1986Natur.319..489B).
- (en) A. Allwood, B. S. Kamber, M. R. Walter, I. W. Burch et I. Kanik, « Trace elements record depositional history of an Early Archaean stromatolitic carbonate platform », Chemical Geology, vol. 270, , p. 148–163 (DOI 10.1016/j.chemgeo.2009.11.013, lire en ligne [PDF])
 Portail de la géologie
Portail de la géologie  Portail origine et évolution du vivant
Portail origine et évolution du vivant