Gène engrailed
Engrailed est une homéoprotéine facteur de transcription[1],[2]. Le gène qui produit cette protéine est ce qu’on appelle un gène de polarité segmentaire[3],[4], c’est-à-dire qu’il permet de définir les cellules antéro-postérieures lors de la segmentation[5].
Engrailed est exprimé dans la partie postérieure des segments lors du développement embryonnaire. Ce gène peut être essentiel pour plusieurs animaux soit pour la segmentation, pour la neurogenèse, pour le développement des appendices ou pour une combinaison de ces éléments. En fait ce gène est essentiel pour obtenir l’axe antéro-postérieur[6] du corps embryonnaire d’un animal. Engrailed, les gènes Hox[1] et plusieurs autres gènes sont essentiels au bon développement de cet axe antéro-postérieur. Ce gène important est d’ailleurs présent chez les annélides, chez les arthropodes, chez les mollusques, chez les échinodermes et chez les chordés[7]. Cet homéogène est donc très important en biologie évolutive du développement étant donné son rôle au stade embryonnaire. Plusieurs études et recherches ont été faites au sujet d’engrailed afin de mieux comprendre son importance dans le développement des animaux.
Origine
Ce gène a d’abord été découvert chez la drosophile (Drosophila melanogaster). Christiane Nüsslein-Volhard et Eric F. Wieschaus[8] furent dans les premiers à décrire son rôle dans la segmentation de ces mouches. Après la découverte de ce gène et de son rôle dans la segmentation, plusieurs chercheurs ont continué les recherches sur cet homéogène et ont émis des hypothèses sur son rôle originel dans les organismes, c’est-à-dire son rôle chez les ancêtres des animaux d’aujourd’hui.
Certains chercheurs pensent que ce gène serait à l’origine responsable du développement du système nerveux, c’est-à-dire de la neurogenèse. D’autres pensent plutôt qu’il aurait d’abord permis la segmentation. Cette dernière hypothèse est plus controversée, puisque plusieurs recherches montrent qu’engrailed ne sert pas à la segmentation de plusieurs animaux[9]. Ce qui est certain c’est que l’étude d’engrailed et de wingless, un autre gène relié à la segmentation, a permis d’en savoir plus sur les ancêtres des ecdysozoaires et des lophotrochozoaires[3], qui devaient probablement être segmentés.
Les recherches sur engrailed nous permettent aussi de supposer que ce gène serait impliqué dans le patron de développement des vertébrés[10].
Voie de signalisation[4] chez la drosophile

La voie de signalisation d’engrailed commence par la transcription de ce gène, qui peut être amorcée par l'homéoprotéine fushi tarazu[11]. Une fois la transcription commencée, une cascade de réaction permet la formation et le maintien des parasegments. En effet, engrailed est exprimé dans le noyau des cellules antérieures d’un parasegment, qui vont devenir, plus tard dans le développement, les cellules postérieures du segment[9]. La protéine qu’engrailed produit est très importante puisqu’elle est un facteur de transcription permettant l’expression du gène hedgehog. Ce dernier produit une protéine qui sort de la cellule et va se lier à sa protéine réceptrice, Patched, sur une cellule antérieure adjacente. Cette réaction va induire l’expression du gène wingless, qui va lui-même produire une protéine qui va sortir de la cellule et aller se fixer sur sa protéine réceptrice. Cette protéine réceptrice c’est frizzled et elle est située sur la cellule ayant exprimé engrailed. Lorsque la protéine de wingless se lie à frizzled, ceci permet de maintenir activée l’expression d’engrailed et de wingless afin de maintenir les segments durant l’embryogenèse. Donc une fois qu’engrailed est activée une première fois, la cascade de réaction qu’elle provoque lui permet de rester activée dans la partie postérieure des segments tout au long du développement de l’animal[12].
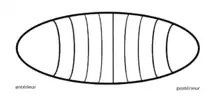
Chez la drosophile
La drosophile est le premier organisme sur lequel a été découvert engrailed. C’est d’ailleurs un organisme modèle dans l’étude de ce gène qui permet le développement des ailes et des segments de la mouche. Ce gène permet, entre autres, le développement normal des ailes des mouches puisqu'une modification dans l’expression de ce gène peut causer un phénotype totalement différent de l’aile et peut parfois transformer la partie postérieure de l’aile en partie antérieure[12]. Cet homéogène est exprimé uniquement dans les parties postérieures des segments et des ailes. En effet, il a été démontré que lors de mutation de ce gène, les parties postérieures des segments se développent en parties antérieures. Ceci en fait un gène très important dans le développement embryonnaire de ces animaux. Chez cet insecte, l’expression du gène possède des limites précises dans le segment. Les cellules exprimant ce gène sont bien délimitées ce qui n’est pas toujours le cas.
Chez les arthropodes
Chez les arthropodes, principalement chez les insectes (dont fait partie la drosophile), engrailed et wingless servent ensemble à la formation de tous les segments, sauf le premier, celui du cerveau, qui n’est formé qu’avec le gène engrailed. Ce dernier est exprimé dans le mésoderme et l’ectoderme de l’animal durant l’embryogenèse[3]. Il permettrait aussi certains patrons de couleurs chez quelques insectes[13].
Chez les araignées et les autres arachnides, engrailed n’a pas tout à fait le même patron d’expression. Deux gènes homologues à engrailed sont exprimés, Cs-engrailed-1 et Cs-engrailed-2. L’expression du premier est très similaire à l’expression d’engrailed dans la drosophile, mais celle du deuxième est suffisamment différentes pour penser qu’elle a un mode d’expression distinct[9]. Le fonctionnement de la segmentation des arachnides n’a pas encore été éclairci et d'autres recherches sont encore nécessaires pour mieux comprendre cette étape essentielle du développement[14].
Chez les onychophores[3]

Les onychophores, un groupe frère des arthropodes, se développent selon le même processus de segmentation que les arthropodes. Tout comme chez ces derniers, le gène engrailed est exprimé dans la partie postérieure des segments lors de l’embryogenèse. Cet animal possède 18 segments, 3 pour la tête et 15 pour le corps, et engrailed est exprimé dans tous ces segments. Pour les onychophores, ce gène sert à la segmentation, au développement des membres et au développement du système nerveux. Chez ces animaux, l’expression d’engrailed n’aurait pas de limite cellulaire précise dans les segments, mais serait assurément dans la partie postérieure. Les études d’engrailed sur ces animaux nous laissent croire que ceux-ci seraient les ancêtres des arthropodes.
Chez les mollusques[15]
Chez les mollusques, la situation est un peu différente puisqu’un nombre différent de copies d’engrailed sont retrouvés selon la classe du mollusque : Chez les bivalves, les chitons et les scaphopodes, il n’y a qu’une copie du gène, alors qu’il y en a deux chez les céphalopodes. Il y a donc des variations du nombre de copies entre les classes, mais parfois aussi au sein d’une même classe[16].

Dans tous les cas, engrailed sert au moins à la délimitation du compartiment, et non à la formation, de la coquille de ces invertébrés, même chez les mollusques dont la coquille est interne et très réduite, comme chez les céphalopodes comme la seiche. Chez les seiches, en plus de délimiter la coquille, engrailed sert aussi au développement de plusieurs autres membres tel que les paupières, les bras et le siphon. Le gène serait donc impliqué dans le développement de nouveautés évolutives[15]. Mais engrailed ne serait pas impliqué dans le développement du système nerveux des mollusques, contrairement à celui d'autres animaux.
Chez les annélides
Chez les annélides, ht-en, l’homologue d’engrailed, serait responsable du développement du système nerveux et dans le développement général, mais ne semble pas impliqué dans la segmentation[9]. Ces animaux ont été relativement peu étudiés et beaucoup de recherches sont encore nécessaires pour mieux comprendre le rôle d’engrailed dans leur organisme.
Chez les échinodermes
Chez les échinodermes les gènes homologues à engrailed varient selon la classe. En effet, chez les oursins[7], le gène homologue à engrailed est SpEn. Ce gène sert à la neurogenèse, c’est-à-dire au développement du système nerveux de l’animal. Ce gène est, entre autres, exprimé dans une cellule ou deux de chaque côté de la bouche des larves, là où des petits cils vont se développer. Chez les étoiles de mer[10], engrailed est exprimé dans le système nerveux et le cœlome, mais au stade larvaire, il est seulement exprimé dans le cœlome. Son expression dans le système nerveux commence dans l’anneau nerveux central des étoiles de mer et plus tard dans le développement, cette expression se propage dans les nerfs radiaux. Engrailed serait aussi exprimé dans les pieds ambulacraires de ces échinodermes.
Chez les vertébrés
Plusieurs homologues d’engrailed sont retrouvés chez les vertébrés. Ce gène est d’ailleurs très conservé au cours de l’évolution des vertébrés[17]. Chez la souris et l’humain, on retrouve En-1 et En-2. Ces gènes servent au développement du cervelet[6]. En présence de mutation de En-1, celui-ci ne se développe pas normalement[18]. En effet, les homologues d’engrailed sont vraiment très importants à l’organisation tridimensionnelle du cervelet[19].
On retrouve aussi des homologues du gène engrailed chez le poisson-zèbre. Cet animal possède au moins deux homologues d’engrailed dont l'un, ZF-En, est très semblable à En-2 présent chez la souris. La similitude entre ces deux gènes permet de supposer qu’il y a une très forte conservation de la fonction de ce gène dans ces organismes[20]. Chez ces poissons, le gène serait exprimé après la gastrulation, alors qu’il est exprimé avant la gastrulation chez les drosophiles. Il sert au développement des somites de ces poissons.
Notes et références
- (en) Brian Gebelein, Daniel J. McKay et Richard S. Mann, « Direct integration of Hox and segmentation gene inputs during drosophila development », Nature, vol. 431, , p. 653-659
- (en) Eric. C. R. Reeve et Isobel Black, Encyclopedia of genetics, London/Chicago, London: Fitzroy Dearborn, , 952 p. (ISBN 1-884964-34-6, lire en ligne), p.167-169.
- (en) Bo Joakim Eriksson, Noel N. Tait, Graham E. Budd et Michael Akam, « The involvement of engrailed and wingless during segmentation in the onychophoran Euperipatoides kanangrensis (Peripatopsidae : Onychophora)(Reid 1996) », Development Genes Evolution, vol. 219, , p. 249-264
- (en) Bruce S. Heming, Insect development and evolution, Ithaca : Comstock Pub. Associates, , 444 p. (ISBN 0-8014-3933-7), p.160-161.
- (en) Di Jiang et William C. Smith, « An ascidian engrailed gene », Development Genes Evolution, vol. 212, , p. 399-402.
- (en) Shunsuke yaguchi, Yoko Nakajima, Diana Wang et Robert D. Burke, « Embryonic expression of engrailed in sea urchins », Gene Expression Paterns, vol. 6, , p. 566-571
- (en) David K. Jacobs et Ruth D. Gates, « Developmental genes and the reconstruction of metazoan evolution – implication of evolutionary loss, limits on inference of ancestry and type 2 errors », Integrative comparative biology, vol. 43, , p. 11-18
- (en) Wim G. M. Damen, « Parasegmental organization of the spider embryo implies that the parasegment is an evolutionary conserved entity in arthrood embryogenesis », Development, vol. 129, , p. 1239-1250
- (en) Maria Byrne, Paula Cisternas, Laura Elia et Bronwyn Relf, « Engrailed is expressed in larval development and in the radial nervous system of Patriella sea stars », Development Genes Evolution, vol. 215, , p. 608-617
- (en) Brian Florence, Antoine Guichet, Anne Ephrussi et Allen Laughon, « Ftz-F1 is a cofactor in Ftz activation of the Drosophila engrailed gene », Development, vol. 124, , p. 839-847
- (en) Alicia Hidalgo, « The roles of engrailed », Trends in genetics, vol. 12, no 1, , p. 1-4
- (en) Andrew D. Peel, Maximilian J. Telford et Michael Akam, « The evolution of hexapod engrailed-family genes : evidence for conservation and concerted evolution », Proceedings of the Royal society B, vol. 273, , p. 1733-1742
- (en) S. Baratte, A. Andouche et L. Bonnaud, « Engrailed in cephalopods : a key gene related to the emergence of morphological novelties », Development Genes Evolution, vol. 217, , p. 353-362
- (en) C.G. Wray, D.K. Jacobs, R. Kostriken, A.P. Vogler, R. Baker et R. DeSalle, « Homologues of the engrailed gene from five molluscan classes », FEBS Letters, vol. 365, , p. 71-74
- (en) Peter W.H. Holland et Nicola A. Williams, « Conservation of engrailed-like homeobox sequences during vertebrate evolution », FEBS letters, vol. 277, nos 1,2, , p. 250-252
- (en) Crystal L. Murcia, Forrest O. Gulden, Natalie A. Cherosky et Karl Herrup, « A genetic study of the suppressors of the Engrailed-1 cerebellar phenotype », Brain Research, vol. 1140, , p. 170-178
- (en) Roy V. Sillitoe, Michael W. Vogel et Alexandra L. Joyner, « Engrailed homeobox genes regulate establishment of the cerebellar afferent circuit map », The journal of neuroscience, vol. 30, no 30, , p. 355-360
- (en) Ander Fjose, Hans G. Eiken, Pål R. Njølstad, Anders Molven et Ivar Hordwik, « A zebrafish engrailed-like homeobox sequence expressed during embryogenesis », FEBS letters, vol. 231, no 2, , p. 355-360
 Portail de la zoologie
Portail de la zoologie