Ankylopollexia
Les Ankylopollexia forment un clade éteint de dinosaures ornithischiens qui vivaient du Jurassique supérieur au Crétacé supérieur. Il s'agit d'un clade dérivé d'ornithopodes iguanodontiens[1],[2] et contient le sous-groupe Styracosterna[3]. Le nom vient du mot grec «ankylos», pris à tort pour signifier raide, fusionné (en fait l'adjectif signifie courbé ou courbé ; utilisé des doigts, cela peut signifier accroché), et du mot latin «pollex» , ce qui signifie le pouce. Décrit à l'origine en 1986 par Sereno, cette caractéristique synapomorphique très probable d'une épine conique du pouce définit le clade[4].

Clades de rang inférieur
- †Camptosaurus
- †Cumnoria
- †Draconyx
- †Owenodon?
- †Uteodon
- †Styracosterna
- †Barilium
- †Bayannurosaurus
- †Cedrorestes
- †Dakotadon
- †Fukuisaurus
- †Hippodraco
- †Hypselospinus
- †Iguanacolossus
- †Lanzhousaurus
- †Lurdusaurus
- †Magnamanus
- †Osmakasaurus
- †Planicoxa
- †Proa
- †Theiophytalia
- †Hadrosauriformes
Apparue pour la première fois il y a environ 156 millions d'années, dans le Jurassique, les Ankylopollexia est devenue un clade extrêmement réussi et répandu pendant le Crétacé et a été trouvée dans le monde entier. Le groupe s'est éteint à la fin du Maastrichtien[3]. Même s'ils sont devenus assez grands, comparables à certains dinosaures carnivores, ils étaient universellement herbivores[5].
Taille
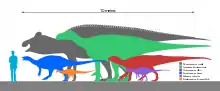
Les ankylopollexiens ont varié considérablement en taille au cours de leur évolution. Le genre jurassique Camptosaurus était petit, ne dépassant pas 5 mètres de long et pesant une demi-tonne[6]. Le plus grand ankylopollexien connu, datant de la fin de la période campanienne (il y a environ 70 millions d'années), appartenait à la famille des hadrosauridés et s'appelle Shantungosaurus. Il mesurait environ 14,7 mètres à 16,6 mètres de long et pesait, pour les plus gros individus, jusqu'à 16 tonnes[7],[8].

Les ankylopollexiens primitifs avaient tendance à être plus petits que les hadrosaures plus grands et plus dérivés. Il existe cependant des exceptions à cette tendance. Une seule trace d'un grand ornithopode, probablement un parent de Camptosaurus, a été signalée dans la Formation de Lourinhã, datant du Jurassique au Portugal. L'animal correspondant avait une hauteur de hanche estimée à environ 2,8 mètres, beaucoup plus grande que le parent contemporain Draconyx[9]. Le Styracosterna primitif Iguanacolossus a été nommé pour sa robustesse distincte et sa grande taille, probablement d'environ 9 mètres de longueur. En ce qui concerne les hadrosaures, l'un des membres les plus basaux des Hadrosauroidea, le genre chinois Bolong, est estimé à environ 200 kilogrammes[10]. Une autre exception à cette tendance est Tethyshadros, un genre plus dérivé des Hadrosauroidea. Estimé à 350 kilogrammes, Tethyshadros n'a été trouvé que sur certaines îles d'Italie. Sa petite taille s'explique par le nanisme insulaire[11].
Classification

Il y a environ 157 millions d'années, Ankylopollexia et Dryosauridae se seraient divisés en branches évolutives distinctes[12]. Initialement nommé et décrit en 1986 par Paul Sereno, les Ankylopollexia recevèrent une définition plus formelle dans un article ultérieur de Sereno en 2005[4]. Dans l'article de 1986, les groupes Camptosauridae et Styracosterna ont été utilisés pour définir le clade, mais dans l'article de 2005, une définition phylogénétique a été donnée : « le dernier ancêtre commun de l'espèce Camptosaurus dispar et Parasaurolophus walkeri et tous ses descendants ».
Le cladogramme ci-dessous suit l'analyse phylogénétique de Bertozzo et al. (2017)[13] :
- Ankylopollexia
- Camptosaurus dispar
- Owenodon hoggii
- Styracosterna
-
- Cedrorestes crichtoni
- Osmakasaurus depressus
-
- Hippodraco scutodens
- Theiophytalia kerri
-
- Iguanacolossus fortis
- Planicoxa venenica
- Dakotadon lakotaensis
-
- Lurdusaurus arenatus
- Lanzhousaurus magnidens
-
- NHMUK R1831
- Kukufeldia tilgatensis
-
- Barilium dawsoni
- Fukuisaurus tetoriensis
-
- Proa valdearinnoensis
-
- Iguanodon bernissartensis
- Hadrosauroidea
- Hypselospinus fittoni
-
- Mantellisaurus atherfieldensis
- NHMUK R3741 (cf. Mantellisaurus)
-
- Ouranosaurus nigeriensis
-
- Altirhinus kurzanovi
- Jinzhousaurus yangi
- Ratchasimasaurus suranareae
-
- Penelopognathus weishampeli
-
- Equijubus normani
- Xuwulong yueluni
-
- Gongpoquansaurus mazongshanensis
-
- Hadrosauromorpha
Paléobiologie
Cerveau

La neurobiologie des ankylopollexiens a été étudiée dès 1871, lorsque John Hulke a décrit un crâne bien conservé (spécimen NHMUK R2501[14]) découvert en septembre 1869 dans le Wealden Group sur l'île de Wight et provisoirement rattaché au genre Iguanodon. Il a noté qu'en raison de la moindre corrélation entre la forme du cerveau et la paroi de la cavité crânienne chez les reptiles, toute déduction de la forme du cerveau de l'animal serait approximative[15]. La référence de ce crâne a été renforcée dans une étude ultérieure, publiée en 1897. On s'est alors demandé si le cerveau du dinosaure pouvait être plus étroitement associé à la cavité que celui des reptiles modernes, et un endocaste a donc été créé et étudié[16]. Ce n'était pas le premier endocaste d'un cerveau ankylopolloxien, car en 1893, le crâne d'un Claosaurus annectens (aujourd'hui rattaché au genre Edmontosaurus[17]) a été utilisé par Othniel Charles Marsh pour créer un moulage de la cavité cérébrale. Certaines remarques de base ont été faites, notamment la petite taille de l'organe, mais il a été noté que l'interprétation des caractéristiques infires de l'organe était difficile[18]. L'article de 1897 notait la similitude des deux endocastes[16].
Les hadrosaures sont réputés avoir le cerveau le plus complexe des ankylopollexiens, et même de l'ensemble des dinosaures ornithischiens. Les cerveaux d'une grande variété de taxons ont été étudiés. John Ostrom, en 1961, a fourni ce qui était alors la revue et le travail le plus complet et détaillé sur la neuro-anatomie des hadrosaures. Ce domaine de l'étude des hadrosaures n'en était alors qu'à ses débuts, et seules les espèces connues aujourd'hui sous les noms d'Edmontosaurus annectens, Edmontosaurus regalis et Gryposaurus notabilis (que l'on considérait à l'époque comme un synonyme de son parent Kritosaurus) disposaient de spécimens appropriés à l'époque pour être examinés (Lambeosaurus était mentionné comme ayant une boîte crânienne brièvement décrite, mais il s'agissait d'une erreur provenant de Lull et Wright (1942))[19],[20]. Ostrom a soutenu l'idée que les cerveaux des hadrosaures et d'autres dinosaures n'auraient probablement rempli qu'une partie de la cavité crânienne, empêchant ainsi la capacité d'apprendre à partir des endocastes, mais il a noté qu'ils étaient encore utiles. Il a noté, comme Marsh, la petite taille prévue de l'organe, mais aussi qu'il était considérablement développé. Il a noté un certain nombre de similitudes avec les cerveaux des reptiles modernes[20].
En 1977, James Hopson a étudié les quotients d'encéphalisation (QE) de divers dinosaures. Trois ornithopodes pour lesquels des endocastes cérébraux avaient été produits précédemment - Camptosaurus, Iguanodon, et Anatosaurus (maintenant connu sous le nom d'Edmontosaurus annectens[17]) - ont été étudiés. On a constaté qu'ils avaient des QE relativement élevés par rapport à de nombreux autres dinosaures (allant de 0,8 à 1,5), comparables à ceux des théropodes carnosauriens et des crocodiliens modernes, mais bien inférieurs à ceux des théropodes coelurosauriens. Ces deux derniers genres, qui ont vécu plus tard que Camptosaurus, avaient des QE un peu plus élevés que le taxon jurassique, qui, se situant à l'extrémité inférieure, était plus comparable au genre cératopsien Protoceratops. Les raisons suggérées pour expliquer leur intelligence comparativement élevée étaient le besoin de sens aigus en l'absence d'armes défensives, et des comportements intraspécifiques plus complexes, comme l'indiquent leurs structures d'affichage acoustique et visuelle[21].

Dans une première pour un vertébré fossile terrestre, Brasier et al. (2017) ont rapporté des tissus mous minéralisés du cerveau d'un dinosaure iguanodontien, provenant de la Formation supérieure de Tunbridge Wells (En) d'âge Valanginien (environ 133 millions d'années) à Bexhill, Sussex. Des restes fragmentaires d'ornithopodes ont été associés au fossile, et bien qu'il n'ait pas été possible d'assigner le spécimen à un taxon avec certitude, Barilium ou Hypselospinus (En) ont été proposés comme candidats probables. Le spécimen se comparait bien aux endocastes de taxons similaires, comme celui d'un Mantellisaurus exposé au Musée d'histoire naturelle de l'Université d'Oxford. Des observations détaillées ont été réalisées à l'aide d'un microscope électronique à balayage. Seules certaines parties du cerveau étaient préservées ; les expansions cérébelleuses et cérébrales étaient les mieux conservées, tandis que les lobes olfactifs et le bulbe rachidien étaient absents ou presque. Les tissus neuronaux semblaient être très serrés, indiquant un CE proche de cinq (les hadrosaures ayant des CE encore plus élevés), correspondant presque à celui des théropodes non aviaires les plus intelligents. Bien qu'il ait été noté que cela correspondait à leur comportement complexe, comme l'avait fait remarquer Hopson, il a été précisé que la densité de l'encombrement pouvait être un artefact de la conservation, et les estimations initiales plus faibles ont été considérées comme plus précises. Certains des comportements complexes attribués peuvent être observés dans une certaine mesure chez les crocodiliens modernes, qui se rapprochent des chiffres originaux[14].

L'avènement de la tomodensitométrie pour la paléontologie a permis une application plus répandue de cette méthode sans qu'il soit nécessaire de détruire les spécimens. Les recherches modernes utilisant ces méthodes se sont largement concentrées sur les hadrosaures. Dans une étude réalisée en 2009 par le paléontologue David C. Evans (En) et ses collègues, les cerveaux des hadrosaures lambéosaurinés des genres Hypacrosaurus (spécimen adulte ROM 702), Corythosaurus (spécimen juvénile ROM 759 et spécimen subadulte CMN 34825), et Lambeosaurus (spécimen juvénile ROM 758) ont été scannés et comparés entre eux (aux niveaux phylogénétique et ontogénétique), à des taxons apparentés et à des prédictions antérieures, ce qui constitue le premier examen à grande échelle de la neurologie de la sous-famille. Contrairement aux premiers travaux, les études d'Evans indiquent que seules certaines régions du cerveau de l'hadrosaure (la partie dorsale et une grande partie du cerveau postérieur) étaient vaguement corrélées à la paroi cérébrale, comme les reptiles modernes, les régions ventrale et latérale étant assez étroitement corrélées. De même, contrairement aux reptiles modernes, le cerveau des jeunes ne semblait pas être plus proche de la paroi cérébrale que celui des adultes. Il faut toutefois noter que les très jeunes individus n'ont pas été inclus dans l'étude[19].
Comme dans les études précédentes, les valeurs de QE ont été étudiées, mais une fourchette de chiffres plus large a été donnée pour tenir compte de l'incertitude concernant la masse cérébrale et la masse corporelle. La fourchette pour l'Hypacrosaurus adulte était de 2,3 à 3,7 ; l'extrémité inférieure de cette fourchette était encore plus élevée que celle des reptiles modernes et de la plupart des dinosaures non maniraptoriens (presque tous ayant des QE inférieurs à deux), mais elle était bien en deçà de celle des maniraptoriens eux-mêmes, qui avaient des quotients supérieurs à quatre. La taille des hémisphères cérébraux a été, pour la première fois, remarquée. On a constaté qu'ils occupaient environ 43 % du volume endocrânien (sans tenir compte des bulbes olfactifs) chez ROM 702. Cette taille est comparable à celle des hadrosaures saurolophinés, mais bien supérieure à celle des ornithischiens en dehors des Hadrosauriformes, et à celle de tous les grands dinosaures saurischiens ; les maniraptors Conchoraptor et Archaeopteryx, un oiseau précoce, avaient des proportions très similaires. Cela renforce l'idée de comportements complexes et d'une intelligence relativement élevée, pour des dinosaures non aviaires, chez les hadrosauridés[19].

Amurosaurus, un proche parent des taxons de l'étude de 2009, a fait l'objet d'un article en 2013 qui s'est à nouveau penché sur un endocaste crânien. Une plage de QE presque identique de 2,3 à 3,8 a été trouvée, et il a de nouveau été noté qu'elle était plus élevée que celle des reptiles vivants, des sauropodes et d'autres ornithischiens, mais différentes estimations de QE pour les théropodes ont été citées, plaçant les chiffres de l'hadrosaurus significativement en dessous même des théropodes plus basaux comme Ceratosaurus (avec une plage de QE de 3. 31 à 5,07) et Allosaurus (avec une fourchette de 2,4 à 5,24, contre seulement 1,6 dans l'étude de 2009[19],[22]) ; les théropodes coelurosauriens plus proches des oiseaux, tels que Troodon, avaient des QE supérieurs à sept. En outre, le volume cérébral relatif n'était que de 30 % chez Amurosaurus, ce qui est nettement inférieur à celui d'Hypacrosaurus, plus proche de celui de théropodes comme Tyrannosaurus (33 %), mais toujours nettement supérieur aux chiffres estimés précédemment pour des iguanodontes plus primitifs comme Lurdusaurus et Iguanodon (19 % chacun). Ceci a démontré un niveau de variation de la neuro-anatomie au sein des Hadrosauridae qui n'avait pas été reconnu auparavant[22].
Paléobiogéographie

Les Ankylopollexiens sont devenus au Crétacé l'un des groupes les plus prospères de la planète, étant à la fois répandu et nombreux dans la nature[23]. À cette époque, les ankylopollexiens se sont répandus en Asie et en Europe. Un premier exemple est le genre chinois Bayannurosaurus, du Berriasien[24]. Le genre le plus ancien, trouvé dans le Wyoming, est Camptosaurus dispar, qui remonte aux alentours du Callovien-Oxfordien, il y a environ 157-156 millions d'années[25].
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Ankylopollexia » (voir la liste des auteurs).
- P. -E. Dieudonné, P. Cruzado-Caballero, P. Godefroit et T. Tortosa, « A new phylogeny of cerapodan dinosaurs », Historical Biology, , p. 1–21 (ISSN 0891-2963 et 1029-2381, DOI 10.1080/08912963.2020.1793979, lire en ligne, consulté le )
- Francisco J. Verdú, Rafael Royo-Torres, Alberto Cobos et Luis Alcalá, « Systematics and paleobiology of a new articulated axial specimen referred to Iguanodon cf. Galvensis (Ornithopoda, Iguanodontoidea) », Journal of Vertebrate Paleontology, , e1878202 (ISSN 0272-4634 et 1937-2809, DOI 10.1080/02724634.2021.1878202, lire en ligne, consulté le )
- (en) A. T. McDonald, « Phylogeny of Basal Iguanodonts (Dinosauria: Ornithischia): An Update », PLoS ONE, vol. 7, no 5, , e36745 (PMID 22629328, PMCID 3358318, DOI 10.1371/journal.pone.0036745, Bibcode 2012PLoSO...736745M)
- Sereno, P.C. (1986). "Phylogeny of the bird-hipped dinosaurs (order Ornithischia)". National Geographic Research 2 (2): 234–56
- (en) Foster, J. (2007). Camptosaurus dispar. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. p. 219-221
- (en) Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 284
- (en) Glut, Donald F. (1997). "Shantungosaurus". Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 816–817. (ISBN 0-89950-917-7).
- (en) Zhao, X.; Li, D.; Han, G.; Hao, H.; Liu, F.; Li, L.; Fang, X. (2007). "Zhuchengosaurus maximus from Shandong Province". Acta Geoscientia Sinica 28 (2): 111–122. doi:10.1007/s10114-005-0808-x
- (en) Octávio Mateus et Jesper Milàn, « Ichnological evidence for giant ornithopod dinosaurs in the Upper Jurassic Lourinhã Formation, Portugal », Oryctos, vol. 8, , p. 47–52 (lire en ligne)
- (en) Wu Wen-hao, Pascal Godefroit, Hu Dong-yu (2010). "Bolong yixianensis gen. et sp. nov.: A new Iguanodontoid dinosaur from the Yixian Formation of Western Liaoning, China". Geology and Resources 19 (2): 127–133
- (en) Dalla Vecchia, F. M. (2009). "Tethyshadros insularis, a new hadrosauroid dinosaur (Ornithischia) from the Upper Cretaceous of Italy". Journal of Vertebrate Paleontology 29 (4): 1100–1116
- (en) Norman, David B.; Weishampel, David B. (1990). "Iguanodontidae and related ornithopods". In Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria. Berkeley: University of California Press. pp. 510–533. (ISBN 0-520-06727-4).
- (en) Filippo Bertozzo, Fabio Marco Dalla Vecchia et Matteo Fabbri, « The Venice specimen of Ouranosaurus nigeriensis (Dinosauria, Ornithopoda) », PeerJ, vol. 5, no e3403, , e3403 (PMID 28649466, PMCID 5480399, DOI 10.7717/peerj.3403)
- (en) Brasier, Martin D., « Remarkable preservation of brain tissues in an Early Cretaceous iguanodontian dinosaur. », Geological Society, London, Special Publications, vol. 448, no 1, , p. 383–398 (DOI 10.1144/SP448.3, Bibcode 2017GSLSP.448..383B, lire en ligne)
- (en) Hulke J.W., « Note on a large reptilian skull from Brooke, Isle of Wight, probably Dinosaurian, referable to the genus Iguanodon », Quart J. Geol Soc, vol. 27, nos 1–2, , p. 199–206 (DOI 10.1144/GSL.JGS.1871.027.01-02.27)
- (en) Andrews, Chas. W., « Note on a cast of the brain-cavity of Iguanodon. », Journal of Natural History, vol. 19, no 114, , p. 585–591 (DOI 10.1080/00222939708680580)
- (en) Benjamin S. Creisler, Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs, Bloomington and Indianapolis, Indiana University Press, , 185–210 p. (ISBN 978-0-253-34817-3), « Deciphering duckbills: a history in nomenclature »
- (en) Marsh, Othniel Charles, « The skull and brain of Claosaurus. », American Journal of Science, vol. 265, no 265, , p. 83–86 (DOI 10.2475/ajs.s3-45.265.83, Bibcode 1893AmJS...45...83M, lire en ligne)
- (en) Evans, David C., « Endocranial Anatomy of Lambeosaurine Hadrosaurids (Dinosauria: Ornithischia): A Sensorineural Perspective on Cranial Crest Function », The Anatomical Record, vol. 292, no 9, , p. 1315–1337 (PMID 19711466, DOI 10.1002/ar.20984)
- (en) Ostrom, John H., « Cranial morphology of the hadrosaurian dinosaurs of North America. », Bulletin of the AMNH, vol. 122, (hdl 2246/1260)
- (en) Hopson, James A., « Relative brain size and behavior in archosaurian reptiles. », Annual Review of Ecology and Systematics, vol. 8, no 1, , p. 429–448 (DOI 10.1146/annurev.es.08.110177.002241)
- (en) Lauters, Pascaline, « Cranial Endocast of the Lambeosaurine Hadrosaurid Amurosaurus riabinini from the Amur Region, Russia », PLOS ONE, vol. 8, no 11, , e78899 (PMID 24236064, PMCID 3827337, DOI 10.1371/journal.pone.0078899, Bibcode 2013PLoSO...878899L)
- (en) MacDonald, Andrew, « New Basal Iguanodonts from the Cedar Mountain Formation of Utah and the Evolution of Thumb-Spiked Dinosaurs », PLOS ONE, vol. 5, no 11, , e14075 (PMID 21124919, PMCID 2989904, DOI 10.1371/journal.pone.0014075, Bibcode 2010PLoSO...514075M)
- (en) Xu, Xing, « A large-sized basal ankylopollexian from East Asia, shedding light on early biogeographic history of Iguanodontia », Science Bulletin, vol. 63, no 9, , p. 556–563 (DOI 10.1016/j.scib.2018.03.016)
- (en) Carpenter, K. and Wilson, Y. (2008). "A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Moument, Utah, and a biomechanical analysis of its forelimb". Annals of the Carnegie Museum 76: 227–263. doi:10.2992/0097-4463(2008)76[227:ansoco]2.0.co;2
 Portail des dinosaures
Portail des dinosaures