Agrobacterium
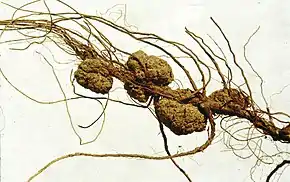
Agrobacterium est un genre de bactéries communes du sol et plus particulièrement du sol parcouru par les racines des plantes, dit sol rhizosphérique. Ces bactéries sont pour la plupart des pathogènes des végétaux (ou phytopathogènes). A. tumefaciens est responsable d'une maladie appelée galle du collet - ou crown-gall selon la dénomination anglo-saxonne - qui se traduit par la formation d'une tumeur au site d'infection. La bactérie a été identifiée à partir de ces tumeurs en 1907 par deux chercheurs américains, E.F. Smith et C.O. Townsend. Le mécanisme de formation des tumeurs s'apparente à une transformation génétique, un fragment d'ADN bactérien étant transféré de la bactérie vers la plante, puis intégré dans le matériel chromosomique végétal. Cette observation constitue le fondement d'une des techniques les plus utilisées en ingénierie génétique des végétaux.
| Règne | Bacteria |
|---|---|
| Embranchement | Proteobacteria |
| Classe | Alpha Proteobacteria |
| Ordre | Rhizobiales |
| Famille | Rhizobiaceae |
Espèces de rang inférieur
- Agrobacterium radiobacter
- Agrobacterium rhizogenes
- Agrobacterium rubi
- Agrobacterium tumefaciens
Il existe d'autres espèces d'Agrobacterium, telles A. vitis, responsable de la maladie du broussin de vigne (galle spécifique de la vigne), ou A. rhizogenes, responsable du hairy root, maladie caractérisée par l'apparition d'un chevelu racinaire au point d'infection. La taxonomie des agrobactéries reste très discutée et évolue constamment.
Description
Le genre Agrobacterium désigne des bacilles d'environ 1 x 3 microns, à coloration de Gram négative (dits gram négatif). Le genre Agrobacterium appartient à la famille des Rhizobioaceae, elle-même incluse dans la classe des alpha-protéobactéries au sein du phyllum des proteobactéries. Les Rhizobiaceae regroupent de nombreuses bactéries capables d'induire la formation de nodosités sur les plantes légumineuses, sièges d'une symbiose permettant la fixation de l'azote.
Les bactéries du genre Agrobacterium sont mobiles, et aérobies stricts pour la plupart. Elles présentent une capacité d'utilisation des glucides très étendue. Leur température optimale de croissance s'étage de 24 à 28°. Dans ces conditions et dans un milieu de culture très favorable, leur temps de génération avoisine 120 minutes. À l'inverse, elles résistent très bien aux carences, aux milieux pauvres et peuvent ainsi survivre plusieurs semaines dans de l'eau pure, à 4 °C. Elles résistent aussi au sels de tellure, et à de nombreux antibiotiques de type bêta-lactame.
Historique
La galle du collet est une maladie connue depuis l'antiquité; ses symptômes ont été décrits par des observateurs grecs et romains. Elle affectait - et affecte toujours - de nombreuses plantes de culture, tels la vigne ou les arbres fruitiers à noyau. En 1907, deux chercheurs américains, E.F. Smith et C.O. Townsend, isolent une bactérie de fragments de galle, qu'ils identifient comme l'agent pathogène responsable de la maladie, nommée Bacterium tumefaciens puis renommée Agrobacterium tumefaciens.
En 1930, Riker et ses collaborateurs, dans le Wisconsin, identifient une bactérie responsable d'une autre maladie des végétaux, le "hairy-root", qui se traduit par l'apparition d'un chevelu racinaire au point d'infection. L'agent responsable sera nommé plus tard Agrobacterium rhizogenes.
En 1942, deux autres américains, P.R. White et A.C. Braun, démontrent que les cellules issues de galles du collet sont des cellules cancéreuses. Débarrassées de la présence d'Agrobacterium, elles prolifèrent facilement et indéfiniment sur des milieux de culture qui ne permettent pas la croissance de cellules saines, en raison de l'absence de régulateurs de croissance végétaux dans ces milieux. A.C. Braun et ses collègues proposent alors l'existence d'un "principe inducteur de tumeur".
À la fin des années 1950, deux groupes de chercheurs français, ceux de Lioret et de Morel, identifient des composés spécifiques des cellules de crown-gall, les opines. Ils proposent en 1972 que la spécificité de synthèse des opines résulte d'un transfert d'informations génétiques entre la bactérie et la plante.
En 1974, l'élément génétique responsable du pouvoir pathogène chez A. tumefaciens est caractérisé par un consortium de chercheurs belges dirigés par J. Schell et M. van Montagu. Il s'agit d'un fragment d'ADN circulaire, ou plasmide, appelé plasmide Ti ou pTi.
En 1977, un autre consortium, américain, dirigé par E. Nester, démontre que la maladie résulte du transfert d'un fragment d'ADN du plasmide Ti, appelé ADN-T ou T-DNA en anglais, de la bactérie vers les chromosomes de la cellule végétale. L'implication d'un plasmide, dit plasmide Ri, et d'un transfert d'ADN sont vérifiés pour A. rhizogenes, à la fin des années 1970 et au début des années 1980.
Enfin, dès 1978, le consortium belge cité plus haut est le premier à proposer que le plasmide Ti pourrait servir de vecteur d'introduction de séquences d'ADN choisies dans le génome végétal, ouvrant ainsi la voie au génie génétique des plantes, et à la création de variétés OGM.
Génome
Le génome d’Agrobacterium tumefaciens se compose de deux chromosomes, l'un linéaire et l'autre circulaire, et dans de nombreux cas de plasmides de haut poids moléculaire. Dans la souche de référence C58, le chromosome 1 est constitué d'environ 2800000 paires de bases (2800 kb) et porte 2833 gènes. Le chromosome 2 est constitué de 2075 kb et porte 1895 gènes. Les plasmides AtC58 et TiC58 comportent respectivement 540 et 210 kb, et regroupent 543 et 198 gènes. Le génome complet est donc constitué de 5675 kb et regroupe 5469 gènes. La séquence nucléotidique du génome de diverses souches d’Agrobacterium est connue et accessible par internet sur le site agrobacterium.org [1].
Listes d'espèces
Selon la LPSN
En 2020 la taxonomie du genre Agrobacterium est en pleine évolution. L'espèce-type Agrobacterium tumefaciens est en cours de reclassement vers une sous-espèce d'Agrobacterium radiobacter dénommée Agrobacterium radiobacter subsp. tumefaciens. Plusieurs autres espèces ont été reclassées vers d'autres genres :
- anc. Agrobacterium atlanticum Rüger & Höfle 1992 : reclassé en Ruegeria atlantica (Rüger and Höfle 1992) Uchino et al. 1999
- anc. Agrobacterium ferrugineum (ex Ahrens & Rheinheimer 1967) Rüger & Höfle 1992 : reclassé en Pseudorhodobacter ferrugineus (Rüger & Höfle 1992) Uchino et al. 2003
- anc. Agrobacterium gelatinovorum (ex Ahrens 1968) Rüger & Höfle 1992 : reclassé en Thalassobius gelatinovorus (Rüger & Höfle 1992) Arahal et al. 2006
- anc. Agrobacterium meteori Rüger & Höfle 1992 : reclassé en Ruegeria meteori (Rüger & Höfle 1992) Hördt et al. 2020
- anc. Agrobacterium rhizogenes (Riker et al. 1930) Conn 1942 : reclassé en Rhizobium rhizogenes (Riker et al. 1930) Young et al. 2001
- anc. Agrobacterium stellulatum (ex Stapp & Knösel 1954) Rüger & Höfle 1992 : reclassé en Stappia stellulata (Rüger & Höfle 1992) Uchino et al. 1999
- anc. Agrobacterium vitis Ophel & Kerr 1990 : reclassé en Allorhizobium vitis (Ophel & Kerr 1990) Mousavi et al. 2016.
Le périmètre actuel délimite onze espèces, dont six ont été décrites après 2000 :
- Agrobacterium arsenijevicii Kuzmanović et al. 2019 sp. nov.
- Agrobacterium cavarae Flores-Felix et al. 2020 sp. nov.
- Agrobacterium fabacearum Delamuta et al. 2020 sp. nov.
- Agrobacterium larrymoorei Bouzar & Jones 2001 sp. nov.
- Agrobacterium nepotum (Puławska et al. 2012) Mousavi et al. 2016
- Agrobacterium pusense (Panday et al. 2011) Mousavi et al. 2016
- Agrobacterium radiobacter (Beijerinck & van Delden 1902) Conn 1942
- Agrobacterium rosae Kuzmanović et al. 2019 sp. nov.
- Agrobacterium rubi (Hildebrand 1940) Starr & Weiss 1943
- Agrobacterium salinitolerans Yan et al. 2017 sp. nov.
- Agrobacterium skierniewicense (Puławska et al. 2012) Mousavi et al. 2016.
Notes et références
Articles connexes
- Pathologie végétale
- Nodule (botanique)
- Agrobacterium radiobacter
 Portail de la microbiologie
Portail de la microbiologie  Portail de la protection des cultures
Portail de la protection des cultures