Cortex préfrontal
Le cortex préfrontal est la partie antérieure du cortex du lobe frontal du cerveau, située en avant des régions prémotrices.
Cette région est le siège de différentes fonctions cognitives dites supérieures (notamment le langage, la mémoire de travail, le raisonnement, et plus généralement les fonctions exécutives). C'est aussi la région du goût et de l'odorat. C'est l'une des zones du cerveau qui a subi la plus forte expansion au cours de l'évolution des primates jusqu'aux hominidés.
Définition
Il existe trois possibilités pour définir le cortex préfrontal :
- le cortex frontal possédant une couche IV granulaire ;
- la zone de projection du noyau médiodorsal du thalamus ;
- la zone du cortex frontal dont la stimulation n'évoque aucun mouvement.
En 1935, Jacobsen utilisa le terme de préfrontal pour désigner la zone du frontal contenant une couche IV granulaire (avec des cellules étoilées et pyramidales) en opposition des aires motrices et prémotrices, agranulaires[1]. Dans la classification de Brodmann, le cortex préfrontal regroupe les aires 8, 9, 10, 11, 44, 45, 46, 47 sur la face latérale, et 12, 32, 24 sur la face médiale (J. Fuster[2]) mais pour compliquer les choses, toutes ces aires ne sont pas strictement granulaires puisque 44 est dysgranulaire, les aires 11 caudale et 47 orbitale sont agranulaires[3]. Pour J. Fuster, un des obstacles majeurs au critère architectonique, est la grande variabilité de la cytoarchitectonique d'espèce à espèce, et entre individus d'une même espèce.
La définition du préfrontal comme zone de projection du noyau médiodorsal du thalamus repose sur les travaux de Rose et Wollsey[4] (1948) qui ont montré que ce noyau se projetait sur les parties antérieures et ventrales du cerveau des non primates. Cette région, qualifiée de orbitofrontale par ces auteurs, fut nommée semble-t-il, la première fois, de préfrontale par Akert, qui en 1964, suggéra que ce critère soit utilisé pour définir les zones homologues du cortex frontal des primates et non primates[5]. Cette définition a l'avantage d'établir des homologies en dépit du manque de cortex granulaire frontal chez les non primates. Elle est encore largement acceptée actuellement, bien que des études de tracés récents aient montré que les projections du noyau médiodorsal ne soient pas restreintes au cortex granulaire du cortex des primates. Pour sauver la définition, il a été suggéré que le cortex préfrontal soit défini comme la région du cortex ayant les connexions réciproques les plus fortes avec le noyau médiodorsal du thalamus[6].
La troisième définition du préfrontal comme la zone du cortex frontal dont la stimulation ne provoque pas de mouvement observable, avait été utilisée par David Ferrier en 1890. Cette définition n'est, elle non plus, pas exempte de défauts, puisque les zones électriquement silencieuses du frontal comprennent des zones granulaires et agranulaires[3].
Subdivisions
Au sein des aires préfrontales, on distingue :
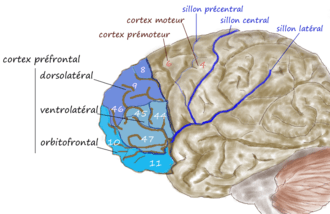
- le cortex préfrontal dorsolatéral (dlPFC) situé sur la face externe (la partie visible du cerveau vu de côté). Cette région dorsolatérale est impliquée dans un réseau qui permet l'élaboration de processus cognitifs. Il joue un rôle majeur dans la planification et les fonctions exécutives ;
- le cortex ventrolatéral ;
- le cortex frontal inférieur (dont le cortex orbitofrontal) qui est situé sous les régions dorsolatérales et au-dessus des orbites oculaires ;
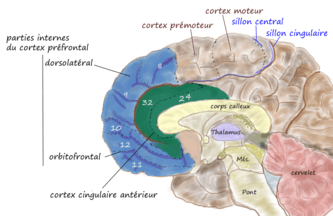
- le cortex frontal médial et cingulaire antérieur qui est situé le long de la scissure interhémisphérique et au-dessus et en avant du corps calleux.
 |
 |
face externe de l'hémisphère gauche |
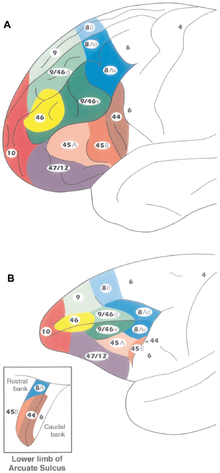
La table ci-dessous donne les différentes subdivisions du cortex préfrontal, face latérale. Le terme dorsolatéral a été utilisé pour renvoyer aux aires 8, 9, 46 au sens restreint, ou à ces mêmes aires plus les aires 44, 45, 47 latérale, au sens large.
| orbitofrontal | cingulaire antérieur | ||||||||||
Roger Gil dans son ouvrage de Neuropsychologie[7] propose un autre découpage du cortex frontal. Il garde l'aire 4 de Brodmann pour l'aire motrice mais étend le cortex prémoteur, puisque outre l'aire 6, il inclut l'aire 8 (aire de coordination oculomotrice), l'aire 44 pars opercularis et 45 pars triangularis du gyrus frontal inférieur, impliquées respectivement dans le traitement phonologique et sémantique du langage (Poldrack et al. 1999) ainsi que l'aire motrice supplémentaire (en) à la face interne de l'hémisphère.
Le cortex préfrontal est caractérisé par le fait que sa lésion entraîne des manifestations désignées sous le nom de « syndrome frontal ». Il est divisé en trois parties :
| dorsolatéral | orbitaire ou ventrale | interne ou mésiale | |||||||
Chez les mammifères
Le cortex préfrontal existe à un état plus primitif chez le rat, mammifère partageant environ 90 % de gènes orthologues stricts avec l'être humain[8], avec une orthologie quasi totale entre humain et rat concernant les « gènes de maladie ». L'étude du cerveau du rat s'avère donc importante pour la conception de médicaments utiles à l'humain. Néanmoins, le cortex préfrontal du rat est moins différencié que celui du primate, et il montre des caractéristiques anatomiques et fonctionnelles qui correspondraient à un mélange des caractéristiques de plusieurs sous-parties du cortex préfrontal des primates[9]. Par exemple, même si certains auteurs considèrent que le cortex préfrontal dorsolatéral (dlPFC) est une zone du cerveau spécifique des primates[10], l'aire prélimbique (PL) du cortex préfrontal du rat présente à la fois des analogies avec le dlPFC en termes d'implication dans la mémoire de travail ou dans le séquencement des tâches comportementales, et à la fois des analogies avec le cortex frontal médian des primates en termes de connexions anatomiques avec d'autres zones du cerveau[9].
Des caractéristiques anatomiques du cortex préfrontal existent également chez le chien[11].
Connexions
Afférences
Le cortex préfrontal établit d'importantes connexions :
- avec les régions sous-corticales : thalamus, sous-thalamus, mésencéphale et système limbique
- avec de nombreuses aires corticales sensorielles associatives
Les voies principales arrivant sur le cortex préfrontal sont celles du noyau médiodorsal du thalamus.
Chez les primates, le noyau médiodorsal se décompose[2] en :
- une composante médiale, dite magnocellulaire, en raison de la présence de cellules de grandes tailles. Cette composante est mieux représentée chez les primates ayant une plus grande ancienneté phylogénétique
- une composante latérale, dite parvocellulaire, en raison de la présence de petites cellules, et mieux représentée chez les espèces plus récentes.
Ces deux composantes ont des projections différentes sur le cortex :
- la partie magnocellulaire se projette sur le cortex préfrontal orbitaire et médial
- la partie parvocellulaire se projette principalement sur le (nouveau) cortex préfrontal latéral.
En outre :
- le pars paralamellaris, un segment latéral du noyau se projette sur l'aire 8 de Brodmann, l'aire de coordination oculo-motrice.
- les parties caudale et dorsale du noyau se projettent sur les aires médiales
Ces projections médiodorsales sur le cortex préfrontal représentent 80 % des projections thalamo-préfrontales, le reste provient du noyau ventral antérieur VA et du noyau intralaminaire rostral. Il existe aussi des afférences qui ne passent pas par le thalamus. On connait ainsi des afférences directes du tegmentum du tronc cérébral, du pont, de l'hypothalamus et de l'amygdale.
En résumé[2], le cortex préfrontal reçoit directement ou indirectement, des informations venant de l'hypothalamus, du sous-thalamus, du mésencéphale, du système limbique et du cervelet.
Le cortex préfrontal est aussi interconnecté avec d'autres régions corticales. Dans la région latérale du préfrontal, convergent des informations de nature visuelles, auditives et somatiques alors que la région orbitomédiale reçoit des informations olfactives, gustatives en plus des informations auditives (et accessoirement visuelles et somatiques). En gros, la collecte d'informations sensorielles servent au cortex préfrontal latéral à organiser et exécuter des actions dirigées vers un but, celles récoltées dans le préfrontal orbitomédial servent aux réactions émotionnelles.
Efférences
Suivant un principe général de connectivité, la plupart des afférences sont réciproques : le cortex préfrontal envoie donc des informations aux structures qui lui en envoient.
Il y a cependant une exception constituée par les ganglions de la base vers lesquels ils envoient des efférences non réciproques.
Fonctions
- Le cortex préfrontal latéral et en particulier les aires de Brodmann 9 et 46 (dorsolatéral) est associé à l’élaboration de processus cognitifs complexes. Il joue un rôle dans le contrôle exécutif tel que la planification et le raisonnement déductif (changement de l'ensemble des règles en cours set-shifting, résolution de problèmes complexes, récupération de souvenirs en mémoire à long terme, stratégies d’organisation et mémoire de travail[12],[13]). On a observé par imagerie médicale l’activation de cette région au cours de tâches de planification comme dans le jeu des tours de Londres.
- Le cortex orbitofrontal (aire 12 et 13) est impliqué dans les processus affectifs et motivationnels à savoir le contrôle du système limbique : inhibition, codage de la valeur motivationnelle d'un stimulus, prise de décisions et contrôle de l'action basée sur la récompense, contrôle de l'humeur, comportement social[14].
Notes et références
- (en) Finger, Stanley, Origins of neuroscience: a history of explorations into brain function, Oxford Oxfordshire, Oxford University Press, , poche (ISBN 978-0-19-514694-3, LCCN 92048265).
- Joaquin M. Fuster (dir.), The Prefrontal Cortex, Academic Press Inc, 4e édit., , 424 p..
- (en) Preuss TM, « Do rats have prefrontal cortex? The Rose-Woolsey-Akert program reconsidered. », Journal of Cognitive Neuroscience (en), vol. 7, no 1, , p. 1–24 (DOI 10.1162/jocn.1995.7.1.1).
- (en) Rose JE, Woolsey CN, « The orbitofrontal cortex and its connections with the mediodorsal nucleus in rabbit, sheep and cat », Research Publications - Association for Research in Nervous and Mental Disease, vol. 27 (1 vol.), , p. 210–32 (PMID 18106857).
- (en) Preuss TM, Goldman-Rakic PS, « Myelo- and cytoarchitecture of the granular frontal cortex and surrounding regions in the strepsirhine primate Galago and the anthropoid primate Macaca », The Journal of Comparative Neurology, vol. 310, no 4, , p. 429–74 (PMID 1939732, DOI 10.1002/cne.903100402).
- (en) Uylings HB, Groenewegen HJ, Kolb B, « Do rats have a prefrontal cortex? », Behavioural Brain Research, vol. 146, nos 1-2, , p. 3–17 (PMID 14643455, DOI 10.1016/j.bbr.2003.09.028, lire en ligne).
- Roger Gil, Neuropsychologie, Masson, 1996, 2003.
- Rat Genome Sequencing Project Consortium (2004) Genome sequence of the Brown Norway rat yields insights into mammalian evolution. Nature, 428(6982):493-521.
- Uylings HBM, Gronewegen HJ, Kolb B (2003) Do rats have a prefrontal cortex ? Behavioural Brain Research, 146:3-17.
- Preuss TM, Kaas JH (1999) Human brain evolution. In: Zigmond MJ, Bloom FE, Landis SC, Roberts JL, Squire LR, editors. Fundamental of neuroscience. San Diego, CA: Academic Press, p. 1283-311.
- Rajkowska G, Kosmal, A (1987) Intrinsic connections and cytoarchitectonic data of the frontal association cortex in the dog. Acta Neurobiol Exp 48:169-92.
- Claire Landmann, Le cortex préfrontal et la dopamine striatale dans l'apprentissage guidé par la récompense, thèse, Université de Paris 6, Neurosciences cognitives, , 213 p.
- Fuster J.M., « The prefrontal cortex - an update : time is of essence », Neuron, vol. 30, .
- Bechara, A., Damasio, H. et Damasio, A. R., « Emotion, decision making and the orbitofrontal cortex. », Cereb cortex, vol. 10, no 3, , p. 295-307.
- Sylvain Pirot, « L'anatomie fonctionnelle du cortex préfrontal : du singe à l'homme », Neuropsychiatrie Tendances et Débats, vol. 20, , p. 27-31.