Développement dentaire
Le développement dentaire ou odontogenèse est le processus complexe par lequel se forment, grandissent et apparaissent les dents dans la bouche. Afin que les dents humaines se développent dans un environnement buccal sain, l'émail, la dentine (ou l'ivoire), le cément et le parodonte doivent tous se développer aux stades adéquats du développement fœtal. Les dents de lait (ou déciduales, ou temporaires) commencent à se développer entre six et huit semaines de vie in utero et les dents définitives vers la vingtième semaine de grossesse[1]. Si les dents n'ont pas commencé à se développer à ces stades, elles ne pourront plus se développer par la suite.
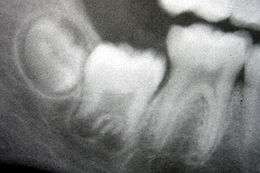
Les processus responsables du déclenchement du développement des dents ont fait l'objet d'un nombre important de recherches. Il est largement admis qu'un certain nombre de molécules (facteurs de croissance, de régulation…) sont indispensables dans les tissus du premier arc branchial pour permettre ce développement[1].
De nombreuses espèces animales possèdent des dents et le développement de ces dernières est pratiquement identique à celui des humains. Chez les vertébrés, plusieurs structures spécialisées du tissu épithélial génèrent après épaississement des structures spécifiques : structures kératinisées (cheveux, ongles) ou structures exosquelettiques (écailles, dents). Les écailles placoïdes et les dents de requin sont considérées comme étant des organes homologues aux dents humaines.
Vue d'ensemble
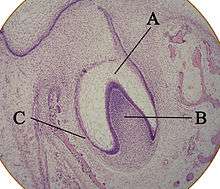
A: organe de l'émail avec la lame dentaire
B: papille dentaire
C: follicule dentaire.
Il se forme chez l'humain, au cours du deuxième mois de grossesse, une lame labiogingivale par prolifération des cellules ectodermiques locales, au niveau du pourtour de l'orifice buccal primitif et se prolongeant en fer à cheval dans la cavité buccale à l'emplacement de ce qui sera les futures mâchoires. Cette lame est formée de l'épithélium épidermique qui repose sur un mésenchyme sous-jacent, va se modifier en se creusant dans sa portion externe pour former l'ébauche du vestibule et en formant une crête interne.
Des cellules épithéliales du versant labial de cette crête vont ensuite proliférer en seize puis vingt endroits et s'enfoncer dans le tissu sous-jacent pour former la lame dentaire, puis les bourgeons dentaires, les ébauches des dents déciduales (ou dents de lait). Par la suite, se formeront à partir de bourgeons issus sur le versant lingual des bourgeons déciduaux les trente-deux ébauches des dents définitives.
La formation d'une dent commence donc par un agrégat cellulaire, le germe dentaire[2], dérivant de l'ectoderme issu du premier arc branchial de la crête neurale[1],[3],[4]. Ce tissu d'origine ectoblastique va se transformer en un tissu mésenchymateux appelé ectomésenchyme ou mésenchyme ectodermique. Le germe dentaire est divisé en trois parties : le bourgeon dentaire (ou organe de l'émail ou encore organe adamantin), issu de l'ectoderme, la papille dentaire et le follicule dentaire, issus du mésenchyme.
L'agrégat de cellules ectodermiques va s'enfoncer dans le mésenchyme sous-jacent et former une lame dentaire dont l'extrémité va s'élargir pour une sorte de coupe renversée avasculaire dans laquelle vont se différencier trois tissus : l'épithélium adamantin externe (EAE) sur la face supéro-externe, l'épithélium adamantin interne (EAI) sur la face inféro-interne et une zone de grandes cellules étoilées, la pulpe de l'émail (ou réticulum étoilé de l'émail, REI), située entre les deux premiers[2]. Les cellules épithéliales continuent ensuite à s'enfoncer dans le tissu sous-jacent en formant une coupe plus profonde, une sorte de cloche, dont la paroi intérieure est formée de cellules de l'EAI, la paroi extérieure de cellules de l'EAE séparées par un tissu gélatineux : la gelée de l'émail. La zone de jonction entre les épithéliums adamantins externe et interne est appelée l'anse cervicale[1]. Les cellules de cette anse qui ne sont pas séparées par le réticulum étoilé vont s'enfoncer de plus en plus dans les tissus profonds, formant la gaine épithéliale de Hertwig qui va donner la racine de la dent. Une partie de l'EAI va voir ses cellules cylindriques se différencier en proaméloblastes puis en améloblastes (ou anciennement adamontoblastes) qui vont sécréter l'émail qui recouvrira la dent.
Le mésenchyme ectodermique situé à l'intérieur de la cloche est appelé papille dentaire. Il va former dans sa partie supérieure, à partir de ses fibroblastes au contact de l'EAI, son propre épithélium, dont les cellules seront appelées odontoblastes et serviront à former l'ivoire qui se déposera entre les amèloblastes et les odontoblastes. Ce dépôt d'ivoire va provoquer en réaction la production d'émail par les améloblastes.
Le mésenchyme situé pour sa part à l'extérieur de la cloche dentaire va se condenser à une certaine distance de l'organe adamantin en un tissu fibreux appelé follicule dentaire, qui deviendra par la suite le ligament alvéolo-dentaire (ou ligament alvéolaire).
Chronologie du développement des dents chez l'homme
Le calendrier de développement des dents chez l'homme est donné dans les tableaux ci-dessous[5]. Les dates de début de calcification des dents de lait sont des semaines in utero (I.U.).
| Dents de lait | |||||
|---|---|---|---|---|---|
| Dents du maxillaire | |||||
| Incisive centrale | Incisive latérale | Canine | Première molaire | Seconde molaire | |
| Début de calcification | 14 sem I.U. | 16 sem I.U. | 17 sem I.U. | 15.5 sem I.U. | 19 sem I.U. |
| Couronne complète | 1,5 mois | 2,5 mois | 9 mois | 6 mois | 11 mois |
| Racine complète | 1.5 ans | 2 ans | 3,25 ans | 2,5 ans | 3 ans |
| Dents de la mandibule | |||||
| Début de calcification | 14 sem I.U. | 16 sem I.U. | 17 sem I.U. | 15.5 sem I.U. | 18 sem I.U. |
| Couronne complète | 2,5 mois | 3 mois | 9 mois | 5,5 mois | 10 mois |
| Racine complète | 1,5 ans | 1,5 ans | 3,25 ans | 2,5 ans | 3 ans |
| Dents définives | ||||||||
|---|---|---|---|---|---|---|---|---|
| Dents du maxillaire | ||||||||
| Incisive centrale |
Incisive latérale |
Canine | Première prémolaire |
Seconde prémolaire |
Première molaire |
Seconde molaire |
Troisième molaire | |
| Début de calcification | 3–4 mois | 10–12 mois | 4–5 mois | 1,5–1.75 ans | 2–2.25 ans | à la naissance | 2,5–3 ans | 7–9 ans |
| Couronne complète | 4–5 ans | 4–5 ans | 6–7 ans | 5–6 ans | 6–7 ans | 2,5–3 ans | 7–8 ans | 12–16 ans |
| Racine complète | 10 ans | 11 ans | 13–15 ans | 12–13 ans | 12–14 ans | 9–10 ans | 14–16 ans | 18–25 ans |
| Dents de la mandibule | ||||||||
| Début de calcification | 3–4 mois | 3–4 mois | 4–5 mois | 1,5–2 an | 2,25–2,5 ans | à la naissance | 2,5–3 ans | 8–10 ans |
| Couronne complète | 4–5 ans | 4–5 ans | 6–7 ans | 5–6 ans | 6–7 ans | 2,5–3 ans | 7–8 ans | 12–16 ans |
| Racine complète | 9 ans | 10 ans | 12–14 ans | 12–13 ans | 13–14 ans | 9–10 ans | 14–15 ans | 18–25 ans |
Le développement des bourgeons dentaires
Une des premières étapes dans la formation d'une dent visible au microscope est la séparation entre le sillon vestibulaire et la lame dentaire.
Le développement dentaire est généralement divisé, par convention, en quatre stades : le stade du bourgeon dentaire, le stade de la cupule, le stade de la cloche et, enfin, le stade de la maturation. Cette division n'est que virtuelle car les changements ont lieu dans les faits de façon continue ; il est souvent difficile de décider quel stade particulier attribuer à une dent en développement[1]. Cette détermination se trouve encore compliquée par l'aspect variable des divers tissus retrouvés sur différentes sections de la même dent en développement, qui peuvent apparaître être situés à des stades différents.
Stade du bourgeon
Le stade du bourgeon est caractérisé par l'apparition, sur le pourtour de l'orifice buccal primitif et se prolongeant en fer à cheval dans la cavité buccale à l'emplacement de ce qui sera les futures mâchoires, d'un tas de cellules épithéliales qui se multiplient et s'enfoncent dans le mésenchyme sous-jacent qui formera la future mâchoire[1]. Ce stade a lieu en général lorsque le fœtus a environ 6 semaines[6]. Le bourgeon dentaire se présente donc comme un amas de cellules au sommet de la lame dentaire.
Stade de la cupule
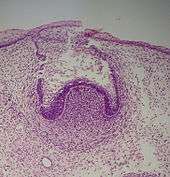
Le début de l'agencement des cellules dans le bourgeon dentaire se produit au stade de la cupule. Dans le mésenchyme, tissu conjonctif, un petit groupe de cellules cesse de sécréter le substrat extracellulaire qui les entoure, ce qui va se traduire par un rapprochement des cellules entre elles et former la papille dentaire. À ce stade, le bourgeon dentaire se développe autour de cet agrégat, prenant la forme d'une cupule et devient l'organe de l'émail. Des cellules du mésenchyme vont s'agglomérer autour de l'organe de l'émail et de la papille dentaire pour former le follicule dentaire. La papille dentaire va alors produire la dentine et la pulpe, l'organe de l'émail va produire de l'émail et le follicule dentaire produire le parodonte, les structures de soutien de la dent[1].
Stade de la cloche
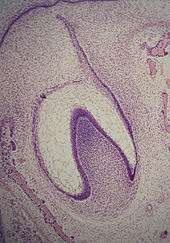
Le stade de la cloche est celui des différenciations histologiques et morphologiques. L'organe de l'émail y prend une forme de cloche et la majorité de ses cellules forment le réticulum étoilé, nom dû à leur forme en étoile[1]. Les cellules de la périphérie de l'organe de l'émail se répartissent en trois couches importantes : les cellules cubiques situées à la périphérie de l'organe de l'émail forment l'épithélium adamantin externe ou latéral[2] ; Les cellules cylindriques de l'organe de l'émail situées du côté de la papille dentaire forment l'épithélium adamantin interne ou médial et les cellules entre l'épithélium adamantin interne et le réticulum étoilé forment une couche connue sous le nom de strate intermédiaire. La zone où les épithéliums externe et interne de l'organe de l'émail se rejoignent est appelée l'anse cervicale[7].
D'autres événements se produisent au cours du stade de la cloche. La lame dentaire qui relie la dent à l’épithélium buccal se désintègre, séparant complètement les dents en développement de l'épithélium de la cavité buccale. Les deux ne seront à nouveau en contact qu'à l'éruption de la dent dans la bouche[1].
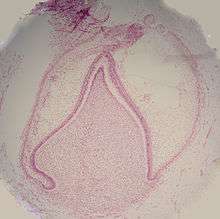
C'est également lors de cette étape que la couronne de la dent prend sa forme, sous l'influence de la forme de l'épithélium adamantin interne. Dans toute la bouche, les dents suivent le même processus, mais on ne connait pas encore avec certitude les raisons qui font que les dents prennent des formes différentes, pourquoi par exemple, les incisives sont différentes des canines. Il y a deux hypothèses dominantes à ce sujet. Le « modèle champ » suppose qu'il y a des éléments pour chaque type de forme de la dent présents dans le mésenchyme pendant le développement dentaire. Ces composants, différents pour les différents types de dents, sont localisés dans une région et disparaissent rapidement dans les autres parties de la bouche. Ainsi, par exemple, le « champ incisive » contient des facteurs qui font que les dents de ce champ se développent sous forme d'incisives et ce facteur concentré dans la région de l'incisive centrale décroît rapidement dans le « domaine canin ». L'autre hypothèse dominante, le « modèle clone », propose comme explication que des cellules épithéliales transmettraient à un groupe de cellules mésenchymateuses des commandes pour générer la forme des différentes dents. Ce groupe particulier de cellules, appelé clone, active la lame dentaire pour qu'elle développe une dent, provoquant la formation du bourgeon dentaire. La lame dentaire continue de croître dans une zone appelée « zone de progression ». Une fois que la zone de progression se trouve à une certaine distance du premier bourgeon dentaire, un deuxième bourgeon dentaire va commencer à se développer. Ces deux modèles ne sont pas nécessairement mutuellement exclusifs, ni largement acceptés par les spécialistes en science dentaire. On suppose seulement que ces deux modèles influencent le développement des dents à des moments différents[1].
D'autres structures peuvent apparaître dans une dent en développement à ce stade : ce sont les nodules de l'émail ou nodules adamantins, les cordons de l'émail et les niches de l'émail.
Stade de la couronne

Les tissus minéralisés, comme l'émail et la dentine, vont se développer au cours de l'étape suivante du développement dentaire. Cette étape actuelle est appelée stade de la couronne ou étape de maturation par certains chercheurs. D'importants changements cellulaires se produisent à ce moment-là. Dans les étapes antérieures, toutes les cellules de l'épithélium adamantin interne se sont multipliées pour augmenter la taille globale du bourgeon dentaire mais cette multiplication par mitose s'arrête pendant le stade de la couronne à l'endroit où les cuspides des dents vont se former. Les premiers tissus minéralisés apparaissent à cet endroit. Au même moment, les cellules de l'EAI changent de forme, passant de cubiques à prismatiques. Les noyaux de ces cellules se rapprochent du stratum intermedium et s'éloignent de la papille dentaire[1].
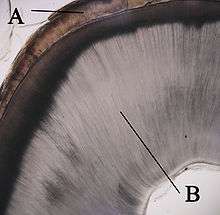
A: émail
B: dentine
Les cellules superficielles de la papille dentaire augmentent brusquement de taille et se différencient en odontoblastes qui vont former la dentine[8]. Les chercheurs croient que l'apparition des odontoblastes ne se ferait pas s'il n'y avait pas eu les changements intervenus dans l'épithélium adamantin interne. En même temps que les cellules de l'épithélium adamantin interne se modifient et que se forment les odontoblastes à partir de la pointe des cuspides, les odontoblastes se mettent à sécréter une substance, une matrice organique, la prédentine, dans leur environnement immédiat. Cette matrice organique contient le matériel nécessaire pour la formation de la dentine. En formant la matrice organique, les odontoblastes reculent et migrent vers le centre de la papille dentaire. Ainsi, contrairement à l'émail, la dentine se forme de l'extérieur vers l'intérieur de la dent. Les odontoblastes, en se déplaçant vers l'intérieur de la papille, laissent des filaments cytoplasmiques dans la matrice, les fibres de Tomes qui en disparaissant laisseront place à de fins canaux, les canalicules de la dentine ou canalicules de Tomes. L'aspect tubulaire de la dentine que l'on observe au microscope est le résultat de la formation de la dentine autour de ces filaments[1].
Après le début de la formation de la dentine, les cellules de l'épithélium adamantin interne sécrètent une matrice organique contre la dentine. Cette matrice se minéralise et devient immédiatement l'émail des dents. En dehors de la dentine, on retrouve les améloblastes, qui sont des cellules qui continuent le processus de formation de l'émail, par conséquent, l'émail se forme de l'intérieur vers l'extérieur par ajout de nouveaux éléments à la surface externe de la dent en développement.
La formation des tissus minéralisés
Émail
La formation de l'émail est appelée amélogénèse et se produit au stade de la couronne. Une induction réciproque régit la relation entre la formation de la dentine et celle de l'émail ; la formation de dentine doit toujours avoir lieu avant la formation de l'émail. En règle générale, la formation de l'émail se fait en deux étapes : la phase de sécrétion et la phase de maturation[9]. Au cours de la phase de sécrétion, se forment un dépôt de protéines et d'une matrice organique donnant un émail partiellement minéralisé. Dans la phase de maturation, le minéralisation de l'émail est achevée.
Pendant la phase de sécrétion, les améloblastes produisent et libèrent des protéines qui contribuent à former la matrice de l'émail, qui est ensuite partiellement minéralisée avec la formation de cristaux d'hydroxyapatite dans cette matrice sursaturée en calcium et en phosphate [10]. L'apparition de ce tissu minéralisé, qui survient généralement vers la troisième ou quatrième mois de grossesse, constitue la première apparition d'émail dans le corps. Les améloblastes commencent leur depôt à l'emplacement de ce qui deviendra les cuspides des dents face à la dentine. La formation de l'émail se poursuit ensuite vers l'extérieur et le bas de la dent, de façon centrifuge.
À la phase de maturation, les améloblastes enlèvent de l'émail certaines des substances utilisées dans sa formation. Ainsi, le rôle des améloblastes change : au lieu de sécréter de l'émail comme c'était le cas dans la phase de sécrétion, ils transportent des substances organiques. La plupart des matières transportées par les améloblastes à ce stade sont des protéines utilisées pour la minéralisation complète. Les principales protéines impliquées dans cette phase sont les amélogénines, les améloblastines, les énamélines et les tuftélines[11]. À la fin de cette étape, l'émail a terminé sa minéralisation.
Dentine
La formation de la dentine, connue sous le nom de dentinogénèse, est la première étape identifiable du stade de la couronne. La formation de la dentine doit toujours avoir lieu avant la formation de l'émail. Les différentes étapes de la formation de la dentine sont reprises des différents types de dentine : la dentine palléale, la dentine primaire, la dentine secondaire et la dentine tertiaire.
Les odontoblastes, les cellules qui vont former la dentine, se différencient des autres cellules de la papille dentaire. Elles commencent par sécréter une matrice organique autour de la zone directement adjacente à l'épithélium adamantin interne en débutant dans la zone qui deviendra la cuspide. Cette matrice organique contient des fibres de collagène de grand diamètre (0,1 à 0,2 um de diamètre)[12]. Les odontoblastes vont ensuite se déplacer vers le centre de la dent, laissant derrière eux une extension appelée fibre de Tomes[1]. Puis, la dentine va se déposer progressivement vers l'intérieur de la dent. Les fibres de Tomes provoquent l'apparition de cristaux d'hydroxyapatite et donc la minéralisation de la matrice. Cette zone de minéralisation est connue comme la dentine palléale et mesure environ 150 µm d'épaisseur[12].
Pendant que la dentine palléale se forme à partir de la substance fondamentale préexistante de la papille dentaire, la dentine primaire se forme par le biais d'un processus différent. Les odontoblastes augmentent de taille, empêchant toutes les réserves extracellulaire à contribuer à la formation d'une matrice organique pour servir de support à la minéralisation. De plus, les plus grands odontoblastes sécrètent du collagène en petite quantité, qui se dépose en rangs serré, servant à une nucléation hétérogène qui servira de base à la minéralisation. D'autres matériaux (comme les lipides, les phosphoprotéines, et les phospholipides) sont également sécrétés[12].
La dentine secondaire se forme après que la formation des racines est terminée et se produit à un rythme beaucoup plus lent. Elle ne se forme pas à vitesse uniforme le long de la dent, mais plus vite le long des sections proches de la couronne d'une dent[13]. Ce développement se poursuit tout au long de la vie et explique le peu de pulpe dentaire retrouvée chez les personnes âgées[12]. La dentine tertiaire, aussi connue comme dentine réparatrice, se forme en réaction à des stimuli, comme l'attrition ou la carie dentaire[14].
Cément
La formation du cément est appelée cémentogénèse et survient tardivement dans le développement des dents. Les cémentoblastes sont les cellules responsables de la cémentogenèse. Il existe deux types de cément : cellulaire et acellulaire[15].
Le cément acellulaire se forme en premier. Les cémentoblastes se différencient à partir des cellules folliculaires qui ne peuvent atteindre la surface de la racine de la dent que lorsque la gaine épithéliale de Hertwig a commencé à se détériorer. Les cémentoblastes sécrètent de fines fibrilles de collagène le long de la surface de la racine disposées perpendiculairement à la surface avant de migrer loin de la dent. Lorsque les cémentoblastes reculent, du nouveau collagène est déposé pour allonger et épaissir les précédents faisceaux de fibres. Des protéines non collagéniques, telles que la sialoprotéine osseuse et l'ostéocalcine, sont également sécrétées[16]. Le cément acellulaire contient une matrice sécrétée par les cémentoblastes et formé de protéines et de fibres. Lorsque la minéralisation commence, les cémentoblastes s'éloignent du cément, et les fibres laissées le long de la surface finissent par rejoindre le ligament parodontal en formation.
Le cément cellulaire se développe lorsque la plus grande partie de la dent est terminée et après entrée en contact avec une dent sur l'arcade opposée[16]. Ce type de cément se forme autour des faisceaux de fibres du ligament parodontal. Les cémentoblastes formant le cément cellulaire sont piégées dans le cément qu'elles produisent.
On estime que l'origine des cémentoblastes des céments cellulaire et acellulaire est différente. L'une des principales hypothèses est que les cellules produisant le cément cellulaire migrent de la zone adjacente de l'os, tandis que les cellules produisant le cément acellulaire proviennent du follicule dentaire[16]. Néanmoins, on sait que le cément cellulaire n'est généralement pas retrouvé dans les dents avec une seule racine[16]. Dans les prémolaires et les molaires, le cément cellulaire se trouve uniquement dans la partie la plus proche de la pointe de la racine et dans les zones interradiculaires pour les racines multiples.
Formation du parodonte
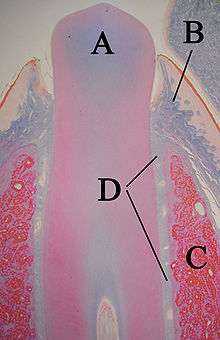
A: dent
B: gencive
C: os alvéolaire
D: ligament parodontal ou desmodonte
Le parodonte, qui est la structure de soutien de la dent, se compose du cément, du ligament parodontal ou desmodonte, de la gencive et de l'os alvéolaire[17]. Le cément est le seul d'entre eux qui fasse partie de la dent. L'os alvéolaire entourant la racine d'une dent lui fournit un appui et forme une cavité adaptée à la forme de la racine qu'on appelle alvéole dentaire. Le ligament parodontal relie l'os alvéolaire au cément et la gencive est le tissu environnant visible dans la bouche [18].
Ligament parodontal
Les cellules du follicule dentaire donnent naissance au ligament parodontal. Les événements spécifiques menant à la formation du ligament parodontal ne sont pas les mêmes pour les dents de lait et les dents permanentes et entre différentes espèces d'animaux[16]. Néanmoins, la formation du ligament parodontal commence avec des fibroblastes du follicule dentaire. Ces fibroblastes sécrètent du collagène, qui interagit avec les fibres situées sur la surface de l'os adjacent et sur le cément[19]. Cette interaction conduit à un amarrage de la racine de la dent lorsque la dent sort de la gencive. L'occlusion dentaire, qui est la manière dont les dents supérieures (dents maxillaires) s'engrènent avec les inférieures, affecte en permanence la formation du ligament parodontal. Cette adaptation perpétuelle du ligament parodontal conduit à la formation de groupes de fibres dans des orientations différentes, telles que les fibres horizontales et obliques[16].
Os alvéolaire
En même temps que la racine de la dent et le cément commencent à se former, de l'os se crée dans les régions adjacentes. Toutes les cellules qui forment des os sont appelées ostéoblastes. Dans le cas de l'os alvéolaire, les ostéoblastes proviennent du follicule dentaire[16]. De la même façon que pour la formation du cément primaire, les fibres de collagène apparaissent sur la surface de l'os la plus proche de la dent et elles y restent jusqu'à la formation du ligament parodontal.
Comme tous les autres os du humain, l'os alvéolaire est modifié tout au long de la vie. Les ostéoblastes et les ostéoclastes créent puis détruisent l'os, surtout si une force est exercée sur la dent[20]. Comme c'est le cas lorsqu'on veut déplacer des dents en orthodontie, une zone osseuse soumise à un effort de compression important fourni par une dent voulant s'en rapprocher, se voit soumise a une forte activité ostéoclastique à ce niveau, entrainant une résorption osseuse. Au contraire, la partie du parodonte mise en tension par une dent qui s'éloigne va voir un grand nombre d'ostéoblastes devenir actifs, entraînant la formation osseuse.
Gencive
La zone de jonction entre la gencive et la dent est appelée jonction dento-gingivale. La gencive possède un revêtement épihélial variable selon les endroits et qu'on peut partager en trois types de tissus épithéliaux : buccal qui tapisse la plus grande partie de la gencive en dehors de l'étroite zone entourant les dents, sulculaire sur le versant externe du sillon dentaire et jonctionnel au contact du collet de la dent. Ces trois types de tissus forment une masse de cellules épithéliales connue sous le nom de manchon épithélial entre les dents et la bouche[16].
Une grande partie de la formation de la gencive n'est pas encore entièrement comprise, mais on sait que des hémidesmosomes se forment entre l'épithélium gingival et la dent et sont responsables de la fixation épithéliale primaire[16]. Les hémidesmosomes fournissent un ancrage entre les cellules par le biais de petites structures filamenteuses fabriquées par les améloblastes restants. Une fois que cela se produit, se forme et se divise rapidement l'épithélium jonctionnel à partir des restes de l'épithélium adamantin. Il en résulte une augmentation continuelle de la taille de l'épithélium jonctionnel et l'isolement des améloblastes de toute source de nourriture. Lorsque les améloblastes vont dégénérer, un sillon gingival va être créé.
Apparition des nerfs et des vaisseaux
Souvent, les nerfs et les vaisseaux sanguins se forment parallèlement dans le corps et la formation des deux a généralement lieu simultanément et de façon similaire. Cependant, ce n'est pas le cas pour les nerfs et les vaisseaux sanguins autour de la dent, à cause de leurs vitesses différentes de développement[1].
Apparition des nerfs
Les fibres nerveuses commencent à apparaître près de la dent au stade de la cupule et croissent en direction du follicule dentaire. Une fois là, les nerfs se développent autour du bourgeon dentaire et entrent dans la papille dentaire lorsque la formation de la dentine a commencé. Les nerfs ne pénètrent pas dans l'organe adamantin[1].
Apparition des vaisseaux
Les vaisseaux sanguins apparaissent dans le follicule dentaire et entrent dans la papille dentaire au stade de la cupule[1]. Des groupes de vaisseaux sanguins se forment à l'entrée de la papille dentaire. Le nombre de vaisseaux sanguins atteint un maximum au début de l'étape de la couronne, et la papille dentaire se transforme finalement en pulpe dentaire. Tout au long de la vie, la quantité de pulpe diminue dans une dent, ce qui signifie que l'approvisionnement en sang de la dent diminue avec l'âge[20]. L'organe de l'émail est dépourvu de vaisseaux sanguins en raison de son origine épithéliale et les tissus minéralisés que sont l'émail et la dentine n'ont pas besoin de nutriments sanguins.
Éruption dentaire
L'éruption des dents se produit lorsque les dents pénètrent dans la cavité buccale et deviennent visibles. Même si les chercheurs conviennent que l'éruption des dents est un processus complexe, il y a peu d'accord sur l'origine du mécanisme qui contrôle cette éruption[21]. Parmi les théories communément admises et qui ont été réfutées au fil du temps, on peut citer :
- La dent est poussée dans la bouche par la croissance de la racine de la dent,
- la dent est poussée à sortir par la croissance de l'os qui l'entoure,
- la dent est poussée par la pression vasculaire, et
- la dent est poussée par un ligament, le ligament en hamac[22].
La théorie du ligament en hamac, d'abord proposée par Harry Sicher, a été largement enseignée des années 1930 aux années 1950. Cette théorie postule qu'un ligament passant en dessous de la dent, que Sicher avait observé au microscope sur une coupe histologique, serait responsable de l'éruption. Plus tard, on s'apercevra que le « ligament » observé par Sicher était simplement un artefact créé par le processus de préparation de la lame[23].
La théorie la plus répandue actuellement est que, si plusieurs forces semblent impliquées dans l'éruption, le ligament parodontal donnerait l'impulsion principale au processus. Les théoriciens émettent l'hypothèse que le ligament parodontal provoquerait l'éruption par le raccourcissement et la réticulation de ses fibres de collagène et la contraction de ses fibroblastes[24].
Bien que l'éruption des dents se produise à différents moments selon les personnes, une chronologie générale des éruptions dentaires existe. En général, les humains ont 20 dents de lait et 32 dents permanentes[25]. L'éruption comporte trois étapes. La première, connue sous le nom stade de la dentition primaire, se produit lorsque les dents de lait sont visibles. Une fois la première dent permanente sortie dans la bouche, les deux types de dents sont présentes dans la bouche : c'est la dentition mixte. Après que la dernière dent de lait est tombée par un processus connu sous le nom d'exfoliation, on est au stade de la dentition permanente.
La dentition primaire commence à l'arrivée des incisives centrales de la mandibule, habituellement à huit mois, et dure jusqu'à ce que les premières molaires permanentes apparaissent dans la bouche, généralement avant six ans[26]. Les dents primaires sortent dans l'ordre suivant: (1). incisive centrale, (2) incisive latérale, (3) la première molaire, (4) canine et (5) deuxième molaire[27]. En règle générale, il y a quatre éruptions de dents tous les six mois ; les dents mandibulaires sortent avant les dents maxillaires et l'éruption des dents se produit plus tôt chez les filles que les garçons[28]. Au cours de la dentition primaire, les bourgeons dentaires des dents permanentes se développent sous les dents primaires, sur le versant lingual ou palatin.
La dentition mixte commence lorsque la première molaire permanente apparaît dans la bouche, généralement à six ans, et dure jusqu'à ce que la dernière dent de lait soit perdue, le plus souvent à onze ou douze ans[29]. Les dents permanentes du maxillaire sortent dans un ordre différent des dents permanentes de la mandibule. Les dents du maxillaire sortent dans l'ordre suivant : (1) première molaire (2) incisive centrale, (3) incisive latérale, (4) première prémolaire, (5) deuxième prémolaire, (6) canine, (7) deuxième molaire et (8) troisième molaire. Les dents de la mandibule sortent dans l'ordre suivant : (1) première molaire (2) incisive centrale, (3) incisive latérale, (4) canine, (5) première prémolaire, (6) deuxième prémolaire, (7) deuxième molaire, et (8) troisième molaire. Comme il n'y a pas de prémolaires dans la dentition primaire, les molaires primaires sont remplacées par les prémolaires permanentes[30]. Si les dents primaires sont perdues avant que les dents permanentes ne soient prêtes à les remplacer, les dents postérieures peuvent se déplacer vers l'avant et réduire l'espace disponible pour la future dent[31]. Cela peut provoquer des chevauchements et/ou des malpositions des dents permanentes, ce qu'on appelle généralement « malocclusion ». Une orthodontie peut être alors nécessaire dans de telles circonstances pour réaligner les dents.
La dentition permanente commence lorsque la dernière dent de lait est tombée, habituellement à 11 à 12 ans, et dure jusqu'à la fin de la vie d'une personne ou jusqu'à ce que toutes les dents soient perdues (édentation). Durant cette phase, les troisièmes molaires (aussi appelées dents de sagesse) sont souvent extraites en raison d'infections locales, de douleurs à l'éruption ou d'impactions dentaires. Les principales raisons de la perte des dents sont la carie et la maladie parodontale[32].
| Âge d'éruption des dents de lait et des dents définitives[33]. | ||||||||
|---|---|---|---|---|---|---|---|---|
| Dents de lait | ||||||||
| Incisive centrale |
Incisive latérale |
Canine |
Première prémolaire |
Seconde prémolaire |
Première molaire |
Seconde molaire |
Troisième molaire | |
| Maxillaire | 10 mois | 11 mois | 19 mois | 16 mois | 29 mois | |||
| Mandibule | 8 mois | 13 mois | 20 mois | 16 mois | 27 mois | |||
| Dents définitives | ||||||||
| Maxillaire | 7–8 ans | 8–9 ans | 11–12 ans | 10–11 ans | 10–12 ans | 6–7 ans | 12–13 ans | 17–21 ans |
| Mandibule | 6–7 ans | 7–8 ans | 9–10 ans | 10–12 ans | 11–12 ans | 6–7 ans | 11–13 ans | 17–21 ans |
Immédiatement après éruption de la dent, l'émail est recouvert par une très fine couche transparente et amorphe spécifique : la membrane de Nasmyth ou cuticule de l'émail, qui est formée de kératine et qui provient de l'organe de l'émail[34],[35].
Nutrition et développement dentaire
Comme pour d'autres parties de l'organisme humain, la nutrition influence le développement des dents. Les nutriments essentiels pour une dentition saine comprennent le calcium, le phosphore et les vitamines A, C et D[36]. Le calcium et le phosphore sont indispensables à la forme des cristaux d'hydroxyapatite, et leurs concentratations dans le sang sont maintenues constantes par la vitamine D. La vitamine A est nécessaires à la formation de la kératine alors que la vitamine C l'est pour le collagène. Le fluor incorporé en faible quantité dans les cristaux d'hydroxyapatite dentaire en formation rend les dents plus résistantes à la déminéralisation et la désintégration[19].
Les carences en ces éléments nutritifs peuvent avoir un large éventail d'effets sur le développement dentaire[37]. Dans les situations où le calcium, le phosphore et la vitamine D sont insuffisants, les structures dures d'une dent peuvent être moins minéralisées. Un manque de vitamine A peut entraîner une réduction de la quantité d'émail formé. Une carence en fluor provoque une augmentation de la déminéralisation dentaire lorsque la dent est exposée à un environnement acide et la reminéralisation est retardée. En outre, un excès d'apport de fluorure à une dent en développement peut conduire à une anomalie connue sous le nom de fluorose.
Anomalies dentaires
Il y a un certain nombre d'anomalies dentaires liées au développement des dents.
L'anodontie est une absence totale de développement dentaire et l'hypodontie l'absence de développement de certaines dents. L'anodontie est rare, survenant le plus souvent chez les porteurs d'une anomalie génétique en rapport avec un trouble du développement ectodermique tandis que l'hypodontie est l'une des anomalies les plus fréquentes du développement, touchant 3,5 à 8,0 % de la population (non compris les troisièmes molaires). L'absence des troisièmes molaires est très fréquente, survenant chez 20-23 % de la population, suivie, en prévalence, par celle de la deuxième prémolaire et d'une incisive latérale. L'hypodontie est souvent associée à l'absence de formation de la lame dentaire, qui est vulnérable au milieu environnemental, tel que certains antibiotiques et la chimiothérapie, et est également associée à de nombreux syndromes, comme la trisomie 21 et le syndrome de Crouzon[38].
L'hyperdontie est le développement des dents surnuméraires. Elle se rencontre chez 1 à 3 % des Caucasiens et est encore plus fréquente chez les Asiatiques[39]. Environ 86 % de ces cas impliquent une seule dent supplémentaire dans la bouche, le plus souvent dans le maxillaire supérieur, au niveau des incisives[40]. L'hyperdontie semble être associée à un excès de lame dentaire.
La courbure ou angulation prononcée anormale d'une racine dentaire, appelée 'dilacération, est presque toujours associée à un traumatisme qui provoque un déplacement de la dent en développement. En effet, lorsqu’une dent se forme, une force peut déplacer une partie de la dent en laissant le reste en place et former un angle anormal. Les kystes ou tumeurs situés à côté d'un bourgeon dentaire peuvent provoquer de telles forces de dilacération, de même que les traumatismes des dents de lait peuvent faire se déplacer le bourgeon dentaire de la dent permanente associée[41].
L’odontodysplasie régionale ou odontodysplasie localisée est une anomalie rare du développement des dents, généralement localisée à la partie antérieure du maxillaire et non héréditaire. Il n'existe aucune prédilection de race, mais les femmes sont plus exposées que les hommes. Les tissus d'origine ectodermique et mésenchymateux sont affectés. L'émail, la dentine et la pulpe de dents sont touchées, les dents sont pâles, avec une racine courte et apparaissent presque transparentes sur les radiographies, décrites comme des « dents fantômes »[42]. La plupart des cas sont considérés comme idiopathiques et non liés aux conditions de grossesse mais certains cas sont associés à des syndromes, des anomalies de croissance, des troubles du système nerveux et des malformations vasculaires. Les dents permanentes ne présentent généralement une odontodysplasie régionale que si les dents de lait avaient été affectées, les deux types de dents étant pratiquement toujours touchés. La plupart du temps, ces dents ne sont pas exposées à un risque accru de carie ou d'inflammation périapicale.
Biologie moléculaire
Chez les poissons, l'expression des gènes HOX intervient pour réguler les mécanismes de l'initiation des dents[43],[44].
Chez la souris la signalisation Wnt est nécessaire à l'initiation du développement des dents[45],[46].
Le facteur NGF-R est présent dans les cellules de la papille dentaire dès le début du stade de la coupe[47] et joue des rôles multiples au cours des événements morphogénétiques et la cytodifférenciation dans la dent[48],[49],[50]. Il existe une relation entre une agénésie dentaire et l'absence du nerf trijumeau (voir Hypodontie).
Tous les stades (bourgeon, coupe, cloche, couronne) de croissance et de morphogenèse des dents sont régis par une protéine : la Sonic hedgehog[45],[51],[52],[53].
Au cours du développement dentaire, il y a de fortes similitudes entre la kératinisation et l'amélogenèse[54],[55]. La kératine est également présente dans les cellules épithéliales du germe dentaire[56] et une mince couche de kératine est présente sur la dent récemment sortie (membrane de Nasmyth ou cuticule de l'émail)[57].
L'émail se comporte comme un centre de signalisation dans la morphogenèse et la différenciation des odontoblastes[58],[59],[60],[61].
Différents éléments phénotypiques modulent la taille des dents[62].
La forme des dents chez l'homme préhistorique était différente de celle de l'homme moderne[62],[63].
Dans certains tératomes (en particulier de l'ovaire, du poumon, du pancréas, des testicules), des dents complètes peuvent se développer[64],[65],[66],[67].
L'éruption des dents nécessite l'hormone parathyroïdienne[68].
Développement des dents chez les animaux
Les animaux dentés ayant le génome le plus simple sont sans doute les vers de la famille des Ancylostomatidae (Ancylostoma duodenale, Necator americanus[69]...).
Les dents sont une structure atavique et leur développement est semblable chez de nombreux vertébrés[52],[70],[71],[72].
Les poissons ont de nombreuses structures osseuses spécialisées[73], il existe des poissons avec dents (Archosargus probatocephalus ordre des Perciformes, famille des Sparidae) et sans dents (les Caristiidae dans l'ordre des Perciformes aussi ont seulement des ébauches de dents présentes chez les juvéniles)[74].
Contrairement à la plupart des animaux, les requins produisent de nouvelles dents en continu tout au long de leur vie[75],[76],[77] via un mécanisme radicalement différent. Comme les dents de requins n'ont pas de racines, les requins peuvent facilement les perdre quand ils se nourrissent (des zoologistes estiment qu'un seul requin peut perdre jusqu'à 2 400 dents en un an[78]) et il doit donc continuellement les remplacer. Les dents de requin sont en fait des sortes d'écailles modifiées qui se forment à proximité de la langue, migrent vers l'extérieur de la mâchoire jusqu'à ce qu'elles soient complètement développées, soient utilisées et finissent par tomber[79].
Les serpents ont généralement des dents, à quelques exceptions près comme le serpent mangeur d'œufs (Dasypeltis scabra).
Aujourd'hui, les oiseaux n'ont pas de dents, mais il semble que les oiseaux préhistoriques, comme l'archéoptéryx, en avaient[80].
Chez les Tubulidentata (Classe des Mammalia) les dents sont sans émail. Ces animaux n'ont ni incisive ni canine, et les molaires ont une croissance continue de la racine [81].
En règle générale, le développement des dents chez les mammifères non-humains est similaire à la formation des dents humaines. Les variations se situent dans la morphologie, le nombre, le calendrier de développement, et les types de dents, mais habituellement pas dans l'évolution réelle des dents.
La formation de l'émail chez les mammifères non humains est presque identique à celui de l'homme. Les améloblastes et organes de l'émail, y compris la papille dentaire, fonctionnent de manière similaire[82]. Néanmoins, si les améloblastes meurent chez l'homme et la plupart des autres animaux, les rendant incapables de continuer à produire de l'émail, les rongeurs en produisent continuellement, ce qui les force à user leurs dents en rongeant divers matériaux[83]. S'ils ne peuvent pas le faire, leurs dents finissent par percer le toit de leur bouche. En outre, les incisives de rongeurs se composent de deux moitiés, connues sous le nom d'analogues de la couronne et de la racine. La moitié vestibulaire est recouverte d'émail et ressemble à une couronne, tandis que la moitié linguale est recouverte de dentine et ressemble à une racine. La racine et la couronne se développent simultanément dans les incisives des rongeurs et continuent de croître pendant toute la vie de l'animal.
La distribution des minéraux dans l'émail des rongeurs est différente de celle des singes, des chiens, des porcs et des humains[84]. Chez le cheval, les couches d'émail et de dentine sont étroitement liées, ce qui augmente la résistance et diminue le taux d'usure des dents[85],[86].
On ne trouve les structures qui créent le ligament parodontal que chez les Mammifères et les Crocodylia[16]. Chez les lamantins, les molaires mandibulaires se développent indépendamment de la mâchoire et sont entourées d'une coque osseuse séparée de l'os par des tissus mous. On retrouve cela également chez les éléphants.
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Tooth development » (voir la liste des auteurs).
- A. R. Ten Cate, Oral Histology: Development, Structure, and Function, 5th ed. (Saint Louis: Mosby-Year Book, 1998), p. 81-102.
- University of Texas Medical Branch.
- Thesleff I, Vaahtokari A, Partanen AM, « Regulation of organogenesis. Common molecular mechanisms regulating the development of teeth and other organs », The International Journal of Developmental Biology, vol. 39, no 1, , p. 35–50 (PMID 7626420, lire en ligne)
- Thesleff I, Vaahtokari A, Kettunen P, Aberg T, « Epithelial-mesenchymal signaling during tooth development », Connective Tissue Research, vol. 32, nos 1-4, , p. 9–15 (PMID 7554939, DOI 10.3109/03008209509013700)
- (en) Major M. Ash et Stanley J. Nelson (trad. du grec ancien), Wheeler's dental anatomy, physiology, and occlusion, Philadelphia, W.B. Saunders, , 8e éd. (ISBN 978-0-7216-9382-8, LCCN 2002030988), p. 32, 45, and 53.
- (en) Barbara Young, Paul R. Wheater (trad. du grec ancien), Wheaters Functional Histology, Elsevier Health Sciences, , 5e éd., poche (ISBN 978-0-443-06850-8, LCCN 2006280114, lire en ligne), p. 255
- University of Southern California School of Dentistry, The Bell Stage: Image 30 found here.
- Ross, Kaye, and Pawlina, Histology: a text and atlas, p. 444.
- Cate, Oral Histology, p. 197.
- Ross, Kaye, and Pawlina, Histology: Text and Atlas, p. 445.
- Ross, Kaye, and Pawlina, Histology: Text and Atlas, p. 447.
- Cate, Oral Histology, p. 128-139.
- Summitt, Fundamentals of Operative Dentistry, p. 13.
- Summitt, Fundamentals of Operative Dentistry, p. 183.
- Johnson, Biology of the Human Dentition, p. 183.
- Cate, Oral Histology, p. 236-248.
- http://www.caradent.net/parodontologie-f36/paro-2-la-gencive-t227.htm
- Luan X, Ito Y, Diekwisch TG, « Evolution and development of Hertwig's epithelial root sheath », Developmental Dynamics, vol. 235, no 5, , p. 1167–80 (PMID 16450392, PMCID 2734338, DOI 10.1002/dvdy.20674)
- Ross, Kaye, and Pawlina, Histology: Text and Atlas, p. 453.
- Ross, Kaye, and Pawlina, Histology: Text and Atlas, p. 452.
- Riolo and Avery, Essentials for Orthodontic Practice, p. 142.
- Harris, Craniofacial Growth and Development, p. 1–3.
- Harris, Craniofacial Growth and Development, p. 3.
- Harris, Craniofacial Growth and Development, p. 5.
- The American Dental Association, Tooth Eruption Charts found here.
- (en) Major M. Ash et Stanley J. Nelson (trad. du grec ancien), Wheeler's dental anatomy, physiology, and occlusion, Philadelphia, W.B. Saunders, , 8e éd. (ISBN 978-0-7216-9382-8, LCCN 2002030988), p. 38 and 41
- (en) Major M. Ash et Stanley J. Nelson (trad. du grec ancien), Wheeler's dental anatomy, physiology, and occlusion, Philadelphia, W.B. Saunders, , 8e éd. (ISBN 978-0-7216-9382-8, LCCN 2002030988), p. 38
- WebMd, Dental Health: Your Child's Teeth found here.
- (en) Major M. Ash et Stanley J. Nelson (trad. du grec ancien), Wheeler's dental anatomy, physiology, and occlusion, Philadelphia, W.B. Saunders, , 8e éd. (ISBN 978-0-7216-9382-8, LCCN 2002030988), p. 41
- Monthly Microscopy Explorations, Exploration of the Month: January 1998 .
- Health Hawaii, Primary Teeth: Importance and Care found here.
- American Academy of Periodontology, Oral Health Information for the Public found here.
- (en) Major M. Ash et Stanley J. Nelson (trad. du grec ancien), Wheeler's dental anatomy, physiology, and occlusion, Philadelphia, W.B. Saunders, , 8e éd. (ISBN 978-0-7216-9382-8, LCCN 2002030988), p. 53
- Armstrong WG, « Origin and nature of the acquired pellicle », Proceedings of the Royal Society of Medicine, vol. 61, no 9, , p. 923–30 (PMID 5679017, PMCID 1902619)
- Darling AI, « The Distribution of the Enamel Cuticle and Its Significance », Proceedings of the Royal Society of Medicine, vol. 36, no 9, , p. 499–502 (PMID 19992694, PMCID 1998608)
- The American Dental Hygiene Association, Nutritional Factors in Tooth Development found here.
- The American Dental Hygiene Association, Table II. Effects of nutrient deficiencies on tooth development found here.
- (en) Declan T. Millett, Richard Welbury (trad. du grec ancien), Orthodontics and Paediatric Dentistry, Édimbourg, Elsevier Health Sciences, (ISBN 978-0-443-06287-2, LCCN 99053892)
- Neville, Damm, Allen, and Bouquot, Oral & Maxillofacial Pathology, p. 70.
- Kahn, Basic Oral & Maxillofacial Pathology, p. 49.
- Neville, Damm, Allen, and Bouquot, Oral & Maxillofacial Pathology, p. 86.
- Kahn, Michael A. Basic Oral and Maxillofacial Pathology. Volume 1. 2001.
- Fraser GJ, Hulsey CD, Bloomquist RF, Uyesugi K, Manley NR, Streelman JT, « An ancient gene network is co-opted for teeth on old and new jaws », PloS Biology, vol. 7, no 2, , e31 (PMID 19215146, PMCID 2637924, DOI 10.1371/journal.pbio.1000031)
- Fraser GJ, Bloomquist RF, Streelman JT, « A periodic pattern generator for dental diversity », BMC Biology, vol. 6, , p. 32 (PMID 18625062, PMCID 2496899, DOI 10.1186/1741-7007-6-32)
- Dassule HR, Lewis P, Bei M, Maas R, McMahon AP, « Sonic hedgehog regulates growth and morphogenesis of the tooth », Development, vol. 127, no 22, , p. 4775–85 (PMID 11044393, lire en ligne)
- Järvinen E, Salazar-Ciudad I, Birchmeier W, Taketo MM, Jernvall J, Thesleff I, « Continuous tooth generation in mouse is induced by activated epithelial Wnt/beta-catenin signaling », Proceedings of the National Academy of Sciences of the United States of America, vol. 103, no 49, , p. 18627–32 (PMID 17121988, PMCID 1693713, DOI 10.1073/pnas.0607289103)
- Christensen L R, Møllgård K, Kjær I, Janas M S., « Immunocytochemical demonstration of nerve growth factor receptor (NGF.-R) in developing human fetal teeth. », Anat Embryol, vol. 188, no 3, , p. 247–55 (PMID 8250280, DOI 10.1007/BF00188216, lire en ligne)
- Mitsiadis TA, Dicou E, Joffre A, Magloire H., « Immunohistochemical localization of nerve growth factor (NGF) and NGF receptor (NGF-R) in the developing first molar tooth of the rat. », Differentiation, vol. 49, no 1, , p. 47–61 (PMID 1320577, DOI 10.1007/BF00495427, lire en ligne)
- Mitsiadis TA, Dicou E, Joffre A, Magloire H., « NGF Signals Supporting the Tooth Development are Mediated through p75. (Japanese) », Journal of the Kyushu Dental Society, vol. 55, no 6, , p. 347–355 (ISSN 0368-6833, DOI 10.2504/kds.55.347, lire en ligne)
- Amano O, Bringas P, Takahashi I, Takahashi K, Yamane A, Chai Y, Nuckolls GH, Shum L, Slavkin HC., « Nerve growth factor (NGF) supports tooth morphogenesis in mouse first branchial arch explants », Dev Dyn, vol. 216, no 3, , p. 299–310 (ISSN 0368-6833, PMID 10590481, DOI 10.1002/(SICI)1097-0177(199911), lire en ligne)
- Cobourne MT, Hardcastle Z, Sharpe PT, « Sonic hedgehog regulates epithelial proliferation and cell survival in the developing tooth germ », Journal of Dental Research, vol. 80, no 11, , p. 1974–9 (PMID 11759005, DOI 10.1177/00220345010800110501)
- Nakatomi M, Morita I, Eto K, Ota MS, « Sonic hedgehog signaling is important in tooth root development », Journal of Dental Research, vol. 85, no 5, , p. 427–31 (PMID 16632755, DOI 10.1177/154405910608500506)
- « Expression of Sonic hedgehog in mouse tooth », Gene expression in tooth by Pekka Nieminen (consulté le 17 octobre 2009)
- Toto PD, O'Malley JJ, Grandel ER, « Similarities of keratinization and amelogenesis », Journal of Dental Research, vol. 46, no 3, , p. 602–7 (PMID 4165207, DOI 10.1177/00220345670460032401, lire en ligne)
- Gustafson G, Sundström B, « Enamel: morphological considerations », Journal of Dental Research, vol. 54 Spec No B, no 2 suppl, , B114–20 (PMID 1094042, DOI 10.1177/00220345750540020301, lire en ligne)
- Domingues MG, Jaeger MM, Araújo VC, Araújo NS, « Expression of cytokeratins in human enamel organ », European Journal of Oral Sciences, vol. 108, no 1, , p. 43–7 (PMID 10706476, DOI 10.1034/j.1600-0722.2000.00717.x, lire en ligne)
- Theodor Rosebury, « Presence of Iron in Enamel Keratin », Journal of Dental Research, vol. 14, , p. 269–72 (DOI 10.1177/00220345340140040301, lire en ligne)
- Vaahtokari A, Aberg T, Jernvall J, Keränen S, Thesleff I, « The enamel knot as a signaling center in the developing mouse tooth », Mechanisms of Development, vol. 54, no 1, , p. 39–43 (PMID 8808404, DOI 10.1016/0925-4773(95)00459-9)
- Tucker AS, Headon DJ, Schneider P, et al., « Edar/Eda interactions regulate enamel knot formation in tooth morphogenesis », Development, vol. 127, no 21, , p. 4691–700 (PMID 11023871, lire en ligne)
- Thesleff I, Keränen S, Jernvall J, « Enamel knots as signaling centers linking tooth morphogenesis and odontoblast differentiation », Advances in Dental Research, vol. 15, , p. 14–8 (PMID 12640732, DOI 10.1177/08959374010150010401)
- Lesot H, Lisi S, Peterkova R, Peterka M, Mitolo V, Ruch JV, « Epigenetic signals during odontoblast differentiation », Advances in Dental Research, vol. 15, , p. 8–13 (PMID 12640731, DOI 10.1177/08959374010150012001)
- Townsend G, Richards L, Hughes T, « Molar intercuspal dimensions: genetic input to phenotypic variation », Journal of Dental Research, vol. 82, no 5, , p. 350–5 (PMID 12709500, DOI 10.1177/154405910308200505)
- Keith A, « Problems relating to the Teeth of the Earlier Forms of Prehistoric Man », Proceedings of the Royal Society of Medicine, vol. 6, no Odontol Sect, , p. 103–124 (PMID 19977113, PMCID 2005996)
- « Ovarian teratoma (dermoid) with teeth », Doctor T's BrokenDown Palace (consulté le 7 novembre 2009)
- Lee R, « On the Nature of Ovarian Cysts which contain Teeth, Hair, and Fatty Matter. », Medico-Chirurgical Transactions, vol. 43, no 2, , p. 93–114 (PMCID 2147752)
- Eccles WM, Hopewell-Smith A, « 'Dermoid Teeth,' or Teeth developed in Teratomata », Proceedings of the Royal Society of Medicine, vol. 5, no Odontol Sect, , p. 123–139 (PMID 19976169, PMCID 2005364)
- Smith CJ, « A teratoma of the lung containing teeth », Annals of the Royal College of Surgeons of England, vol. 41, no 5, , p. 413–22 (PMID 6061946, PMCID 2312017)
- Philbrick WM, Dreyer BE, Nakchbandi IA, Karaplis AC, « Parathyroid hormone-related protein is required for tooth eruption », Proceedings of the National Academy of Sciences of the United States of America, vol. 95, no 20, , p. 11846–51 (PMID 9751753, PMCID 21728, DOI 10.1073/pnas.95.20.11846)
- « Ancylostoma duodenale »(Archive • Wikiwix • Archive.is • Google • Que faire ?), Nematode.net Genome Sequencing Center (consulté le 27 octobre 2009)
- James WW, Wellings AW, « The Dental Epithelium and its Significance in Tooth Development », Proceedings of the Royal Society of Medicine, vol. 37, no 1, , p. 1–6.12 (PMID 19992735, PMCID 2180846)
- Koussoulakou DS, Margaritis LH, Koussoulakos SL, « A curriculum vitae of teeth: evolution, generation, regeneration », International Journal of Biological Sciences, vol. 5, no 3, , p. 226–43 (PMID 19266065, PMCID 2651620, lire en ligne)
- Salazar-Ciudad I, Jernvall J, « A gene network model accounting for development and evolution of mammalian teeth », Proceedings of the National Academy of Sciences of the United States of America, vol. 99, no 12, , p. 8116–20 (PMID 12048258, PMCID 123030, DOI 10.1073/pnas.132069499)
- Sander Kranenbarg, « Skeletal tissue differentiation in fish », Wageninger University (consulté le 24 octobre 2009)
- (en) Richard Owen, The principal forms of the skeleton and the teeth as the basis for a system of natural history and comparativa anatomy, Houlston and Wright, (lire en ligne)
- Dave Abbott, Sharks, found here.
- Boyne PJ, « Study of the chronologic development and eruption of teeth in elasmobranchs », Journal of Dental Research, vol. 49, no 3, , p. 556–60 (PMID 5269110, DOI 10.1177/00220345700490031501, lire en ligne)
- Sasagawa I, « The fine structure of initial mineralisation during tooth development in the gummy shark, Mustelus manazo, Elasmobranchia », Journal of Anatomy, vol. 164, , p. 175–87 (PMID 2606790, PMCID 1256608)
- Jason Buchheim, A Quick Course in Ichthyology, found here.
- Michael E. Williams, Jaws: The early years, found here.
- Sire JY, Delgado SC, Girondot M, « Hen's teeth with enamel cap: from dream to impossibility », BMC Evolutionary Biology, vol. 8, , p. 246 (PMID 18775069, PMCID 2542379, DOI 10.1186/1471-2148-8-246)
- "Class Mammalia, Order Tubulidentata, Family Orycteropodidae, Species Orycteropus afer," University of Michigan Museum of Zoology. Page accessed November 16, 2009.
- Frandson and Spurgeon, Anatomy and Physiology of Farm Animals., p. 305.
- Caceci. Veterinary Histology with subtitle "Digestive System: Oral Cavity" found here.
- Fejerskov O, « Human dentition and experimental animals », Journal of Dental Research, vol. 58, no Spec Issue B, , p. 725–34 (PMID 105027, DOI 10.1177/002203457905800224011, lire en ligne)
- Randall-Bowman's April 2004 "Gummed Out: Young Horses Lose Many Teeth, Vet Says." See reference here.
- Encarta, whose link can be found « here »(Archive • Wikiwix • Archive.is • Google • Que faire ?) (consulté le 8 avril 2013)
{kind=link}
Voir aussi
- Dentition
- Denture
