Corps géniculé latéral
Le corps géniculé latéral (corpus geniculatum laterale, anciennement corps genouillé externe) du thalamus est une partie du cerveau qui traite l'information visuelle provenant de la rétine.

1 Pulvinar thalami, 2 Colliculi superioris,
4 Colliculi inferiores, 7 Corps géniculé latéral


P : parvocellulaire ; M : magnocellulaire ;
K : koniocellulaire (d'après Casagrande et al. 2007).
Les termes scientifiques « géniculé » et « genouillé » viennent du latin : geniculatum, geniculatus, « courbé à la manière d'un genou plié ».
Description
Afférences et efférences
Le corps géniculé latéral (CGL) reçoit l'information directement de la rétine et envoie des projections dans le cortex visuel primaire, dans la couche 4c. Mais ce cortex visuel V1 exerce une forte rétroaction sur le CGL puisque les 8/10e des connexions excitatrices qui y pénètrent proviennent de V1 et de structures sous-corticales.
Les cellules ganglionnaires de la rétine envoient leurs longs axones au corps géniculé latéral via le nerf optique, le chiasma optique et le tractus optique. Ils sont plus d'un million de fibres à suivre cette voie qui contourne le mésencéphale, chemine sur la face médiale du lobe temporal et aboutit (pour 80 % d'entre elles) dans le corps géniculé latéral.
Le CGL se situe à la partie inférieure du diencéphale, au-dessus du mésencéphale. Il est relié en arrière au tubercule quadrijumeaux antérieur, par le « bras conjonctival » ; et en avant à la bandelette optique, qui circonscrit le mésencéphale jusqu'au chiasmas des nerfs optiques, dans l'étage antérieur de la base du crâne.
Structure
Le CGL est une structure paire et symétrique, de part et d'autre du mésencéphale.
Il est formé de six couches distinctes caractérisées chacune par leurs afférences de fibres du nerf optique et leurs efférences vers le cortex visuel primaire. Elles sont numérotées de 1 à 6, en commençant par la région ventrale, concave. Elles s'empilent les unes sur les autres comme des crêpes, en s'enroulant autour du tractus optique comme l'articulation du genou, d'où vient leur qualificatif de « géniculé » (du latin geniculatum composé de genu, « genou », et de culum, culus, « fesse », « courbé »).
Trois types de neurones, différents par leur taille, sont distinguées dans le CGL[1] : les cellules de grande taille des couches 1 et 2 (dites « magnocellulaires » M), les cellules assez petites des couches 3 à 6 (dites « parvocellulaires » P) et les cellules encore plus petites des 6 sous-couches intercalées (dites « koniocellulaires » K, ou « coniocellulaires »).
Les 2 couches magnocellulaires du CGL reçoivent les axones des cellules ganglionnaires, elles aussi, de grandes tailles, et dites « parasol », car leurs dendrites forment un système élargi en parasol, capable de rassembler les signaux venant de collection aléatoire de cônes R, V, B ou de multiples bâtonnets (via les amacrines). Les 4 couches parvocellulaires reçoivent les axones des cellules ganglionnaires de petites tailles (dites « naines », midget), recevant l'information de cônes R et V. Et les sous-couches koniocellulaires reçoivent les axones des cellules ganglionnaires « bistratifiées » connectées à plusieurs cônes B de la rétine[2],[3] .
Organisation
La transmission nerveuse de la rétine au cortex, se fait suivant des voies parallèles, capables de transporter des aspects différents de la scène visuelle, tout en pouvant conserver en partie l'information sur les relations topographiques au sein du champ visuel[4]. Sur le gros million (1,25 106) de fibres ganglionnaires sortant de la rétine par le nerf optique, 90 % sont de type P, 5 % de type M et le reste de type K (koniocellulaires). La ségrégation entre ces trois types est maintenue au niveau du CGL puis du cortex V1. Ils font partie des multiples voies parallèles et fonctionnellement indépendantes allant de la rétine au cortex.
Entrées : projections rétino-géniculées
La projection des objets du champ visuel se fait de manière très structurée sur le CGL.
Quelques principes régissent l'organisation de la projection sur le CGL[3] :
- 1. Chaque CGL ne reçoit ses informations que du champ visuel controlatéral.
- Ainsi, le CGL droit reçoit les stimuli provenant du champ visuel gauche. Et comme le système optique de l’œil renverse l'image (haut/bas et gauche/droit), ces stimuli provenant de gauche activent les hémirétines droites de chaque œil (à savoir l'hémirétine nasale de l’œil gauche et l'hémirétine temporale de l’œil droit).
- 2. Les couches 1, 4, 6 du CGL reçoivent les axones de l'hémirétine nasale controlatérale, les couches 2, 3, 5 du CGL reçoivent les axones de l'hémirétine temporale homolatérale (fig. 5). Cette organisation est liée au croisement des fibres issues de l'hémirétine nasale au niveau du chiasma et à la poursuite du même côté des fibres issues de l'hémirétine temporale.
| Principe de projection | ||
| Couches du CGL | Hémirétine | Hémichamp visuel |
|---|---|---|
| M1, P4, P6 | nasale controlatérale | controlatéral |
| M2, P3, P5 | temporale homolatérale | |
- 3. Un objet situé dans un quadrant visuel se projette en six représentations rétinotopiques, une par couche.
- Les six représentations rétinotopiques (cartographiques) sont situées à des endroits comparables du CGL.
- 4. Des lieux proches dans un hémichamp visuel correspondent à des lieux proches sur la rétine et se projettent en des lieux proches du CGL. Des stimuli situés dans le quadrant supérieur (resp. inférieur) activent la partie latérale inférieure (resp. médiane supérieure) du CGL[5]. La région centrale de la fovéa se projette sur la partie postérieure du CGL et les parties périphériques sur l'avant.
- 5. Les six couches présentent de nettes différences cytoarchitectoniques. Les neurones des deux couches inférieures, numérotées 1 et 2, ont des axones d'un calibre bien plus gros que ceux des neurones des couches supérieures (3, 4, 5, 6), eux-mêmes plus gros que ceux des sous-couches ventrales[6].
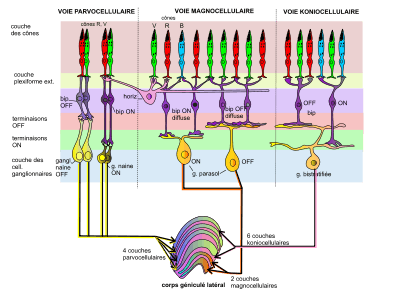
- Les fibres de petits calibres proviennent des cellules ganglionnaires naines de type P (parvocellulaires) de la rétine, dotées de petits champs récepteurs et se trouvant dans la rétine centrale, riche en cônes R et V. Cette voie P transporte des informations sur de fins détails spatiaux d'une scène statique, avec des données sur les couleurs (opposition de couleurs rouge/vert).
- Les fibres de gros calibre proviennent des cellules ganglionnaires parasol, dites par extension de type M (magnocellulaires), pourvues de grands champs récepteurs et se trouvant dans la rétine périphérique, riche en bâtonnets. Cette voie M, spécifiquement activée par la vue d'objets en mouvement, après un relai dans l'aire corticale V1, aboutit sur l'aire V5 (ou MT).
- La voie K (koniocellulaire) fournit la dimension bleu/jaune à la vision des couleurs[7].
- Ainsi, de même que les trois types de cônes correspondent aux composantes de l'espace colorimétrique CIE RGB, on peut rapprocher ces trois voies des composantes du modèle colorimétrique YUV:
- Y', somme pondérée de R, G et B pour constituer la luminance, correspond à la voie M (magnocellulaire) ;
- U, différence entre le bleu et du jaune (somme pondérée de R et G), correspond à la voie K (koniocellulaire) ;
- V, différence entre R et G (avec cependant une légère composante B), correspond à la voie P (parvocellulaire).
Sorties : projections géniculo-striées
Les fibres allant du CGL au cortex strié (ou aire 17 de Brodmann, ou aire V1) forment le faisceau des radiations optiques. Le cortex strié est situé sur les lèvres inférieure et supérieure de la scissure calcarine, sur la face interne des lobes occipitaux gauche et droite. Les fibres qui se projettent sur la lèvre inférieure de la scissure calcarine se dirigent d'abord en avant dans le lobe temporal, en tant que boucle de Meyer, avant de revenir en arrière.
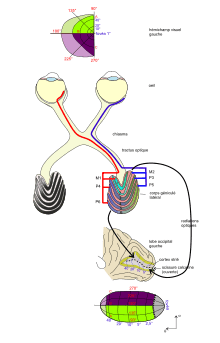
L'organisation très stricte qui régit la projection des fibres de la rétine sur le CGL continue aussi à régir la projection de ce dernier sur le cortex strié[3].
- Les hémichamps visuels se projettent controlatéralement : l'information provenant du champ visuel gauche après un relais dans le CGL droit, aboutit sur le cortex strié de l'hémisphère droit. Lorsqu'on fixe le regard sur un objet, chaque hémisphère ne traite qu'une moitié de l'objet, du fait que les fibres entrant dans le CGL font une projection controlatérale du champ visuel et celles sortant du CGL restent du même côté.
- Le quadrant supérieur du champ visuel se projette sur la berge inférieure de la scissure calcarine et le quadrant inférieur sur la berge supérieure. Par exemple, le quadrant visuel supérieur, gauche (en vert, fig. 5), se projette sur la berge inférieure de l'hémisphère droit. Le méridien horizontal est projeté le long du fond de la scissure calcarine.
- La fovéa, une petite région au centre du champ visuel, se projette sur une région proportionnellement plus grande (en grisé, fig. 5), dans la partie postérieure du cortex strié. La partie binoculaire de chaque hémichamp visuel se projette dans la région intermédiaire du cortex strié. La partie temporale, monoculaire de chaque champ visuel se projette sur la partie antérieure du cortex visuel, près du splénum du corps calleux.
- La ségrégation des informations provenant de la rétine gauche et droite est maintenue du CGL au cortex. On a vu que les couches 1, 4, 6 du CGL recevaient l'information de l’œil controlatéral et les couches 2, 3, 5, celle de l’œil homolatéral. Cette ségrégation gauche/droite se maintient au niveau du cortex strié sous la forme de colonnes de dominance oculaire. La frontière entre ces colonnes est bien distincte dans la couche IV du cortex où la plupart des cellules sont activées par un seul œil. Chez l'homme, ces colonnes de 0,5 mm de diamètre, sont dirigées perpendiculairement à la frontière des aires V1 et V2.
Rétinotopie
L'organisation spatiale des objets du champ visuel est cartographiée dans le cortex visuel primaire V1 (et les aires V2, V3, VP, V3A, V3v) suivant un principe de projection dit « rétinotopique » suivant lequel les lieux proches sur la rétine se projettent en des points proches du cortex.
La continuité de cette application comporte cependant quelques exceptions. Ainsi, bien que l'on puisse passer continûment de l'hémichamp visuel gauche à l'hémichamp droit, il n'en est pas de même dans le cortex où la projection saute d'un hémisphère à l'autre : l'hémisphère gauche ne reçoit l'information que de l'hémichamp visuel droit, et vice-versa.
L'organisation rétinotopique du cortex s'établit donc par un homéomorphisme entre les positions dans le champ visuel et les positions sur la surface corticale au sein des aires rétinotopiques[8]. Mathématiquement, dire que deux objets sont homéomorphes revient à dire que l'on peut passer de l'un à l'autre par une déformation continue.
Notes
Références
- Dale Purves, G-J Augustine, D. Fitzpatrick, W-C Hall, LaManta, McNamara, Williams, Neurosciences, De Boeck, , 811 p.
- (en) Arne Valberg, Light Vision Color, Wiley, , 462 p.
- (en) Rudolf Nieuwenhuys, Jan Voogd, Chr. van Huijzen, The Human Central Nervous System, Springer, 1978, 2008, 967 p.
- (en) Stewart D Shipp, « Parallel Visual Pathways », ACNR, Visual Neuroscience, vol. 6, no 1,
- (en) Kastner S, Schneider KA, Wunderlich K., « Beyond a relay nucleus: neuroimaging views on the human LGN », Prog Brain Res, vol. 155, .
- Gazzaniga, Ivry, Mangun, Neurosciences cognitives La biologie de l'esprit, De Boeck Université, , 585 p..
- (en) Hendry SH, Reid RC., « The koniocellular pathway in primate vision. », Annu Rev Neurosci, vol. 23, , p. 127-53.
- Jan Warnking, Délinéation des aires visuelles rétinotopiques chez l’homme par IRM fonctionnelle, thèse de physique, Université Joseph Fourier - Grenoble,


