Stratégie de reproduction
Une stratégie de reproduction, est un ensemble structuré de comportements ou de règles qui s'applique aux événements reproductifs, le but étant de maximiser la fitness (ou valeur sélective) d'un individu ou d'une espèce. Elle porte notamment sur le choix du partenaire sexuel (offrande et parade nuptiale), les stratégies et systèmes d'accouplement, le type de fécondation et l'investissement parental.
L'évolution de ces stratégies est régie par la sélection naturelle. Au cours du temps, les stratégies de reproduction évoluent parallèlement à celle du milieu. Ces modifications du milieu vont s'assimiler à des poussées évolutives[1].
Plusieurs stratégies peuvent être observées selon un environnement précis. Elles sont fonction de l'espèce et de l'environnement[2]. Le biologiste Robert Trivers a cherché à les modéliser sous le concept d'effort reproductif[3], stratégie idéale qui repose sur deux points principaux, l'effort parental (temps et énergie consacrés à l'investissement parental) et l'effort sexuel (production de gamètes, temps et énergie consacrés à la recherche et la conquête de partenaire et à l'éloignement des rivaux)[4].
Historique
Charles Darwin (1809 – 1882) fut le premier à apporter l'idée que les espèces évoluent au cours des temps grâce à un processus particulier : la sélection naturelle. Grâce à ses expéditions sur le Beagle il fut témoin d'une biodiversité importante résultant de processus de tri aboutissant à une diversité interspecifique et intraspécifique.
Après avoir étudié le travail de l'économiste Thomas Robert Malthus (1766 – 1834), Darwin conçut la théorie de la lutte pour l'existence. Cette théorie démontre que les espèces évoluent au sein de leur milieu afin d'optimiser l'accès aux ressources naturelles. Darwin a également introduit le concept de sélection sexuelle dans les années 1870. Pour lui l'apparition de caractères sexuels secondaires serait due uniquement à la préférence des femelles pour ces traits. Cependant, il n'a pas abordé la question de l'origine de cette préférence. Alfred Russel Wallace amena l'idée qu'un trait était présent du fait qu'il ait été favorisé par la sélection naturelle. Et toujours selon lui, la sélection sexuelle n'était qu'un processus d'amplification de la sélection naturelle.
Le débat entre Darwin et Wallace resta figé jusqu'à la contribution de Ronald A. Fischer. Sa théorie de l'emballement permit d'associer sélection sexuelle et sélection naturelle comme processus d'évolution des espèces (Runaway de Fisher)[1].
Stratégies de reproduction chez les plantes et les animaux
Les stratégies de reproduction chez les plantes sont multiples : apomixie, fécondation croisée ou allogamie, autofécondation ou autogamie. Le régime de reproduction peut être décrit par un modèle mathématique appelé mixed mating model (en).
Les principaux systèmes d’accouplement chez les animaux sont la monogamie et la polygamie (polygynie, polyandrie et polygynandrie).
Il arrive que des animaux utilisent des stratégies alternatives d'accouplement (en), notamment dans les espèces polygynes lorsque le mâle assure le monopole sur les femelles.
Évolution des stratégies de reproduction
Pression via la sélection naturelle
La sélection naturelle est un mécanisme permettant d'accroître la fréquence de traits conférant un avantage sélectif pour les individus qui les portent. Ces traits, pour un milieu donné, apportent une meilleure survie à l'individu qui va pouvoir transmettre ce gène à sa descendance et améliorer ainsi sa fitness. On aura alors une héritabilité du trait considéré comme avantageux. La sélection naturelle va permettre l'adoption ou le rejet de différentes stratégies.
L'apparition de la reproduction sexuée
La dichotomie entre reproduction asexuée et sexuée est la division la plus globale au sein des stratégies de reproduction. Certains taxons de l'arbre du vivant (notamment les unicellulaires, les cnidaires et certains embryophytes) se reproduisent de façon asexuée, par mitose, bourgeonnement, scissiparité ou strobilation. Elle fut le mode de reproduction des premiers êtres vivants. Elle est rapide, simple, peu coûteuse en énergie et ne requiert la présence d'aucun partenaire (pas de brassage génétique).
La reproduction sexuée est apparue et s'est fixée chez la plupart des espèces. Elle a un coût : temps passé à la recherche d'un partenaire, à le garder, à protéger les femelles ; diminution des chances d'avoir d'autres petits (moins de temps pour se nourrir, chercher d'autres partenaires …) ; augmentation du risque de prédation lors de la copulation, coût énergique plus élevé. Mais elle apporte aussi des gains : elle permet le brassage génétique, la variabilité du génome, et se trouve donc à la base de toute l'évolution ; elle permet de lutter contre les maladies et les parasites ; elle augmente les chances de survie.
Sans la diversité génétique permise par la reproduction sexuée, le génome reste une entité relativement fixe qu'il est difficile de faire évoluer.
Dans un environnement stable et à court terme, la reproduction asexuée est donc une excellente stratégie de reproduction, permettant par exemple de coloniser rapidement un milieu. Mais dans un environnement plus variable, et sur le long terme, la reproduction sexuée est avantageuse, car elle permet une meilleure adaptabilité via la sélection naturelle des meilleurs génomes. C'est la raison pour laquelle elle est si répandue.
Une autre stratégie adoptée notamment par certaines plantes consiste à alterner ces deux modes de reproduction selon les conditions du milieu. Par exemple, certains taxons sont partiellement revenus à la reproduction asexuée afin de coloniser rapidement de grands espaces. La reproduction peut ainsi être asexuée en cas d'environnement stable, et devenir sexuée lorsque l'environnement devient variable et requiert des adaptations. Il s'agit probablement d'une stratégie à grand potentiel évolutif, mais qui reste moins adaptable que la reproduction sexuée.
L'équilibre entre auto-fécondation et allo-fécondation
Chez les organismes sexués possédant plusieurs organes reproducteurs (principalement les plantes à fleurs), deux stratégies de reproduction sont possibles : l'auto-fécondation ou l'allo-fécondation. Soit il y a croisement des gamètes de l'individu entre elles, soit entre individus différents. Ces deux stratégies existent dans la nature, ne s'excluent pas l'une l'autre, et sont souvent utilisées toutes les deux au sein d'une même espèce.
Près de 50 % des plantes à fleurs sont capables de faire de l'auto-incompatibilité, un mécanisme complexe d'exclusion pollinique, permettant d'éviter toute auto-fécondation. À l'inverse, la cléistogamie est un mécanisme peu répandu permettant d'éviter toute allo-fécondation (les fleurs ne s'ouvrent jamais, si bien qu'aucun grain de pollen étranger ne peut atteindre le pistil). De très nombreuses plantes se situent entre ces deux extrêmes que constituent l'auto-incompatibilité et la cléistogamie.
Un compromis évolutif existe chez chaque espèce de plante à fleurs entre le taux d'auto-fécondation et d'allo-fécondation, afin de conserver une grande diversité génétique et de limiter la consanguinité (par l'allo-fécondation), tout en étant le plus compétitif possible dans un contexte de colonisation d'un habitat (par l'auto-fécondation). Chaque taxon, au cours de l'évolution, a répondu à ce compromis d'une façon différente, avec un taux d'auto-fécondation approprié à son environnement.
Compromis entre traits d'histoire de vie
Chaque organisme aura des caractéristiques qui lui sont propres et qui influenceront sa valeur sélective. Ces caractéristiques sont appelées traits d'histoire de vie, ce sont les caractères associés au taux de reproduction, à l'âge de la première reproduction et à l’espérance de vie. Ils sont sexe-dépendants et intraspécifiques.
L'évolution des histoires de vie implique des compromis entre des demandes contradictoires ou en compétition pour des variables comme le temps, l'énergie ou les ressources. Parmi les compromis les plus importants, nous pouvons citer la taille et le nombre des jeunes à la naissance ou la croissance et la reproduction.
Les histoires de vie varient selon le taux d'accroissement de la population. Certains ont proposé que dans les environnements imprévisibles, les caractères sélectionnés sont ceux augmentant le taux d'accroissement (r) alors que dans les environnements plus stables, ce sont ceux augmentant la compétitivité et l'efficacité d'utilisation des ressources qui sont sélectionnés. David Lack montra que les traits d'histoire de vie variaient en fonction des paramètres environnementaux, ce qui suggère qu'ils puissent être soumis à la sélection naturelle[5].
Ce phénomène peut être observé chez les morues (Gadus morhua) femelles du Nord-Est de l'Arctique. Elles répartissent leurs ressources de différentes manières entre la croissance, la longueur et la reproduction. Les poissons atteignant la maturité sexuelle plus précoce sont de plus grande taille à trois ans que les autres poissons immatures mais le début de la reproduction ralentit leur croissance. Les chercheurs ont constaté que les femelles qui ne frayent pas une seconde fois à six et sept ans ont une croissance significativement plus rapide cette année-là que les femelles qui frayent une deuxième année consécutive. Ainsi, sauter une année de frai peut constituer une stratégie d'adaptation étant donnée la plus grande fécondité future que procure une plus grande taille[6].
Afin d'éviter un trade-offs trop important, certaines espèces sont capables de mettre en place une hiérarchie entre les individus appelée « Cooperative breeders » (reproduction communautaire). Ceci correspond à un système social où des individus auxiliaires remettent à plus tard ou oublient leur reproduction afin d'aider des individus reproducteurs. De cette façon, les auxiliaires exploitent les reproducteurs en augmentant leur fécondité. Les reproducteurs ayant ainsi une haute mortalité, les contraintes liées à ce statut permet de minimiser les conflits existants entre reproducteurs et auxiliaires dans une population[7].
Pression via la sélection sexuelle
La sélection sexuelle ne peut exister que chez les espèces à reproduction sexuée. Elle va être influencée par la variation du sex-ratio. C’est un processus de choix parmi les traits des partenaires. Cette notion est intraspécifique et sexe-dépendant. La sélection sexuelle permet d’expliquer l’apparition des caractères sexuels secondaires[8].
La sélection sexuelle agit via deux mécanismes principaux : Sélection du partenaire par le sexe investissant le plus dans la reproduction (femelle) favorisant l'apparition d'ornementations chez le sexe qui investit le moins dans la reproduction (mâle). Compétition entre individus favorisant l'apparition d'armes chez les individus du sexe investissant le moins dans la reproduction (mâle).
On parle de « vivre vite mourir jeune » des mâles car ils sont soumis à un taux de mortalité plus élevé que les femelles. Cela est dû à leur investissement pour la production des caractères sexuels secondaires[9]. Selon Darwin, la sélection sexuelle se traduit par une lutte entre les mâles pour la possession des femelles. Le perdant du conflit a un risque d’avoir une descendance limitée ou pas de descendance du tout. Il y a aussi une compétition entre femelles selon leur choix (Sarah Blaffer Hrdy 1984) mettant en avant leur importance dans les processus évolutifs.
Compétition spermatique
Dans certains cas la femelle peut s’accoupler avec plusieurs mâles et pourra alors sélectionner le sperme le plus performant (au niveau du tractus génital). Le mâle va alors être doté de plusieurs mécanismes afin de favoriser son don génétique[1].
Les caractères sexuels secondaires
La sélection sexuelle favorise le dimorphisme sexuel comme avoir de plus grand bois pour un cerf, de posséder des ornements, d’être le plus grand… afin d’être le meilleur compétiteur pour la reproduction. La manifestation de caractères sexuels secondaires tels que les armes ou l’ornementation contribue à orienter le choix des femelles et constitue donc une stratégie à part entière[10].

L’hypothèse de l’apparition de ces attributs serait comme des « signaux honnêtes » envoyés aux femelles sur la qualité génétique des mâles. Les femelles préférant ces traits engendrent une progéniture avec une meilleure survie. Cette hypothèse de signaux rejoint celle du handicap (Amotz Zahavi 1971). L’apparition d’un ornement telle que la couleur vive des rayures chez les guppies favorise la prédation car la proie semble plus attractive. Donc, seuls les mâles les plus forts peuvent porter ce handicap.
- La théorie de l'emballement fisherien
Ce modèle de sélection sexuelle explique l'apparition de caractères sexuels secondaires. La préférence P des femelles entraîne l’apparition du trait T chez les mâles et inversement : Au cours du temps la préférence devient plus forte pour des caractères plus prononcés.
Il peut y avoir une corrélation génétique positive entre le trait T et la préférence P : Les mâles porteurs de traits exagérés T sont préférentiellement choisis par les femelles ayant une forte préférence P pour le trait. Ainsi cela suffit à créer cette corrélation. Du point de vue évolutif, T et P peuvent se fixer au niveau du génome en tant que « bon gène ». La sélection positive sur T va entrainer une sélection positive de P et peut favoriser l'apparition d'un emballement évolutif (ou Runaway de Fisher) dans lequel les deux caractères se renforcent mutuellement. Ce processus se produit dans le cas où la sélection est dite disruptive (quand la sélection est stabilisante l'emballement fisherien ne peut pas se produire).
Cependant, la sélection sexuelle s'oppose à la sélection de viabilité (agit pour améliorer la probabilité de survie d'un individu). Donc ce que l'on peut prédire : quand les avantages liés à la sélection sexuelle sont compensés par la sélection de la viabilité : l'évolution du trait « s'arrête ». Ainsi, il se peut que le trait T disparaisse (la préférence peut également disparaitre).
Maturité sexuelle
L’âge à maturité sexuelle est un paramètre influençant la reproduction. Des pressions telles que la densité vont le faire varier. En effet, si la population est faible, l’âge à maturité sexuelle aura tendance à être précoce et inversement. Par exemple, dans une zone de pêche fortement exploitée (la densité de la population diminue) l'âge à maturité sexuelle sera plus précoce afin de maintenir la stabilité de la population.
L’élément clé pour la reproduction chez les mâles nécessite l’excrétion d’hormones spécifiques comme la testostérone qui permet le développement de caractères sexuels secondaires[9].
Pression via l'environnement
L'hétérogénéité des environnements et des ressources impose une adaptation aux milieux de vie. Des pressions vont s'exercer sur les individus.
Levins a déterminé une classification séparant les habitats selon leur coût reproductif.
On distingue :
- Les habitats « à coût de reproduction faible » :
Ce type d'habitat est plutôt instable (changements climatiques réguliers, imprévisibles...). Il y a une forte mortalité indépendante de la taille des individus. Les individus vivant dans cet environnements ont un investissement reproductif très fort et une croissance amoindrie. La reproduction prime sur la taille des individus (compromis)
- Les habitats « à coût de reproduction élevé » :
Ce type d'habitat, lui, est plutôt stable. La valeur sélective est très sensible à la taille des individus. L'investissement reproductif y est faible. La croissance prime sur la reproduction (compromis). L'âge à maturité sexuelle des occupants de cet habitat est souvent tardif. La compétition (intraspécifique ou interspécifique) y est très forte. En général, les mâles doivent être les plus compétitifs pour gagner ou protéger un harem de femelles. Ce type de milieu inclut souvent un investissement parental important.
Qui dit environnements différents dit stratégies d'adaptation différentes. Le modèle évolutif r/K proposé par Robert MacArthur et E. O. Wilson en 1967, et auquel d'autres, comme Eric Pianka, ont participé, est devenu le plus classique de ces modèles dépendant de l'environnement qui permettent d'expliquer ces adaptations propres à chaque habitat[11].
- Stratégie r : Les « espèces r » vivent plutôt dans les environnements à faible coût reproducteur et donc instables. Dans ce cas, les ressources sont considérées comme non limitantes et donc il n'y a presque pas de compétition. Ces « espèces r » vont avoir tendance à se reproduire tôt (âge à maturité précoce dû à l'instabilité de l'environnement) et tout investir dans leur reproduction en une seule fois (le nombre de descendants est relativement important). La valeur sélective (notamment la survie) étant fortement affectée, les individus meurent souvent après leur reproduction. : c'est ce qu'on appelle la sémelparité (se reproduire une seule fois au cours de sa vie).
Exemple : le bambou, le saumon, les éphémères...
- Stratégie k : Les « espèces K » vivent plutôt dans les environnements à fort coût reproducteur et donc stables. Les ressources du milieu sont alors limitantes et la compétition y est forte. Les individus vont favoriser leur croissance avant leur reproduction pour pouvoir lutter face à la compétition (la défense d'un territoire, l'accès à la reproduction, la prédation...). L'âge à maturité sexuelle est tardif (l'environnement étant stable il n'est pas nécessaire de se reproduire tôt). Il y a plusieurs épisodes reproductifs au cours de la vie d'un individu. C'est ce qu'on appelle l'itéroparité. Des soins parentaux vont être donnés aux descendants.
Exemple : l'homme, les plantes vivaces...
Cependant, ces stratégies sont considérées comme des « extrêmes » ; il existe un continuum entre elles. Les individus peuvent s'adapter sans besoin d'évoluer grâce à leur plasticité phénotypique. On constate alors que l'environnement exerce une influence sur différents paramètres et est fortement lié à l'évolution des espèces[12],[13].
Cas particuliers
Mimétisme lié à la reproduction
.jpg.webp)
Afin de maximiser leurs chances de reproduction certaines espèces vont développer des morphologies particulières. On peut citer comme exemple l’Orchidée de Darwin qui va imiter l'apparence et l'odeur d'une abeille femelle. Le mâle va vouloir aller à la rencontre de la « femelle » et va donc se poser sur la fleur et la polliniser.
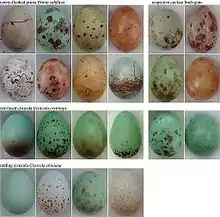
Une autre stratégie bien connue est celle du Coucou ou de l'Anomalospize parasite. La femelle pond ses œufs dans le nid d'une autre afin de ne dépenser aucun coût dans les soins parentaux tout en maximisant sa fitness. Par exemple, l’œuf du coucou éclos en général avant les autres et le petit peut se débarrasser des autres œufs afin d'être le seul à être nourri.
Mais pourquoi la femelle ne reconnait-elle pas l’œuf intrus et ne s'en débarrasse-t-elle pas ? Car ces oiseaux ont mis au point une stratégie mimant les œufs des espèces qu'ils décident de parasiter. Ainsi malgré le doute, l'espèce parasitée préférera garder tous les œufs plutôt que de risquer la perte de l'un des siens. Mais il arrive que l'intrus soit reconnu et éjecté du nid.
On pourrait se poser la question suivante : pourquoi cette stratégie ne s'est-elle pas fixée chez bon nombre d'autres espèces ?
Altruisme lié à la reproduction
L'altruisme lié à la reproduction est un des paradoxes les plus importants de l'histoire de l’Évolution. Cette stratégie est observée, par exemple, chez les hyménoptères ou chez certains oiseaux. Certains individus vont « sacrifier » leur propre reproduction en faveur d'autres individus. Cependant, la sélection naturelle favorise le succès reproducteur individuel et non le « bien de l'espèce » comme on pourrait le penser.
Alors, comment expliquer l'apparition et la fixation de ces comportements chez certaines espèces ? Ce problème n'avait pas échappé à C. Darwin qui remettait en cause sa propre théorie : ces comportements altruistes diminuent l’aptitude de celui qui les manifeste et augmentent celle d’un ou de plusieurs congénères. En effet, le coût des « égoïstes » est moins élevé que celui des altruistes. Donc, ces comportements devraient tendre à disparaître, cependant ils se sont stabilisés et fixés[1].
La sélection de parentèle serait la clé permettant d'expliquer ce type de sélection. Les liens de parenté engendreraient un comportement altruiste : les individus apparentés favorisent la propagation de leur gènes (Loi d'Hamilton)[14].
Elle est indispensable pour qu'il y ait de l'altruisme.

Contraintes à la reproduction
Depuis l’avancement des études sur l’eusocialité chez les fourmis , beaucoup d’études s’étaient concentrées sur le contrôle génétique ou comportemental de la capacité reproductrice des ouvrières, qui se doit d’être maintenue pour assurer la cohésion sociale de la colonie et le maintien des castes reproductrices, notamment en se penchant sur la relation génétique entre les ouvrières ainsi que sur les comportements de police. Cependant peu de chercheurs s’étaient concentrés sur cette question : comment, d’un point de vue développemental, les sociétés de fourmis empêchent-elles leurs ouvrières de se reproduire aux dépens de la reine ?
Khila et Abouheif[15],[16] se sont penchés sur cette question dans deux études parues en 2008 et en 2010.
Selon ces auteurs, les sociétés de fourmis utilisent plusieurs stratégies développementales affectant le système reproducteur de leurs ouvrières, appelées contraintes à la reproduction, en les empêchant ou en limitant leur production d’embryons.
Dans le but d’observer le développement des œufs d’ouvrières de leur formation jusqu’à leur éclosion ou dégénérescence, les auteurs ont utilisé des techniques d’immonufluorescence leur permettant d’observer la polarisation de certaines protéines dans les oocytes de diverses espèces de fourmis. En comparant ainsi les oocytes de la mouche à fruit Drosophila Megalonaster à différentes espèces de fourmis aux diverses structures sociales, les auteurs ont pu observer un changement dans la polarisation de la protéine Vasa et des ARN messager nanos dans l’oocyte chez certaines colonies.
En comparant les structures reproductrices au microscope de différentes espèces de fourmis sous différentes conditions, les auteurs ont remarqué que selon un changement environnemental, la longueur des ovarioles de certaines colonies pouvait varier, probablement à la suite d'un changement dans un mécanisme moléculaire. En raison d'une contrainte à la reproduction, les structures reproductrices de différentes espèces peuvent également être modifiées. Ainsi, la perte de la spermathèque ou d’un certain nombre d’ovarioles peut être observée chez différentes espèces.
La perte complète des structures reproductrices lors du développement a également été observée à la suite d'une technique d’immunofluorescence effectuée sur des cellules embryonnaires d’ouvrières à en devenir.
Ces différentes stratégies de contrainte à la reproduction peuvent complètement bloquer les capacités reproductives des ouvrières ou les forcer à faire plus d’œufs trophiques servant à la colonie. Les cinq stratégies principales développementales de contrainte à la reproduction découvertes par Khila et Abouheif[15],[16] peuvent être résumées ainsi :
1- La localisation des protéines Vasa et des ARN messagers nanos, des déterminants maternels, hors de leur pôle d’expression habituel dans les oocytes de certaines ouvrières amènent une dégénérescence des embryons à la suite d'un dérèglement dans le patron d’expression de ceux-ci et la formation d’œufs trophiques. Ce mécanisme prive donc les ouvrières de leur capacité à produire des mâles à l’aide d’œufs non fécondés.[15]
2- La régulation de certains mécanismes moléculaires, possiblement dans ceux régulant l’insuline, permet de varier la longueur des ovarioles et donc la quantité de division des cellules souches germinales et la production de kystes reproducteurs chez certaines ouvrières. Cette régulation se fait à la suite d’un signal environnemental, puisqu’elle change en l’absence ou la présence d’une reine.Le changement dans le mécanisme moléculaire n’a pas été observé directement, mais est probablement lié à l’insuline, puisque cette voie est impliquée dans le développement de structures reproductrices chez Drosophila Megalonaster.
3- Une perte de la spermathèque chez certaines ouvrières empêche celles-ci de féconder leurs œufs et donc de produire des filles.
4- Une baisse de la quantité d’ovarioles chez certaines ouvrières par la dégénération de certaines cellules polaires lors de leur embryogenèse contrôle leurs capacités reproductrices.
5- Une perte totale du système reproducteur chez les certaines ouvrières les empêchent de produire des œufs. Lors de leurs développements embryonnaires, les cellules germinales des ouvrières sont donc entièrement éliminées.
La contrainte à la reproduction, lorsque développementale, s’articule donc soit par un changement dans le patron de développement des œufs des ouvrières à la suite d'un signal génétique, comme dans la stratégie 1, soit par un signal environnemental qui change le patron de développement des structures reproductrices via une modification d’une voie moléculaire, comme dans la stratégie 2 ou soit par une modification génétique du patron de développement lors du développement des structures reproductrices des ouvrières avant leur sortie de l’œuf comme dans les stratégies 3,4 et 5.
La contrainte à la reproduction, combinée aux contrôles comportementaux et à la relation génétique entre les ouvrières, est essentielle au maintien de l’eusocialité complète des fourmis. Les fourmilières ayant un meilleur système de maintien strict de l’eusocialité sont plus compétitives que celle ayant un système plus permissif puisqu’elles sont sujettes à moins de compétition interne entre les ouvrières et la reine. La stratégie 1 donne aussi un avantage compétitif considérable en changeant le patron de développement des œufs pour qu’ils deviennent des œufs trophiques, puisque ceux-ci permettent de nourrir la colonie de manière plus efficace tout en empêchant la reproduction des ouvrières.
Notes et références
- Écologie comportementale - Cours et questions de réflexion. Étienne Danchin, Luc-Alain Giraldeau, Frank Cézilly - Dunod, 11 mars 2005 - p. 3-20
- Environnement. Linda R. Berg, Peter H. Raven, Dr David .M. Hassenzahl - De Boeck Supérieur, 9 novembre 2009 - p. 186-187
- (en) Robert Trivers, « Parental investment and sexual selection », in B. Campbell (Ed.) Sexual selection and the descent of man, Aldine, p.139
- Georges B. Johnson, Peter H. Raven, Kenneth A.. Mason, Jonathan B. Losos, Susan R. Singer, Biologie, De Boeck Supérieur, , p. 1184
- Écologie, par Robert E. Ricklefs,Gary L. Miller - De Boeck Supérieur, 15 avril 2005 - p. 662-675
- Arild Folkvord, Christian Jørgensen, Knut Korsbrekke, Richard D.M. Nash, Trygve Nilsen and Jon Egil Skjæraasen. Trade-offs between growth and reproduction in wild Atlantic cod. published at www.nrcresearchpress.com/cjfas on 8 April 2014
- David V. McLeod, Geoff Wild. The relationship between ecology and the optimal helping strategy in cooperative breeders. Journal of Theoretical Biology 354 (2014) 25-34
- Jean-François Lemaître, Jean-Michel Gaillard, Josephine Pemberton, Tim H. Clutton-Brock and Daniel H. Nussey, Early life expenditure in sexual competition is associated with increased reproductive senescence in male red deer - Proceedings of The Royal Society, Biological sciences 13 août 2014
- R. Bonduriansky, A. Maklakov, F. Zajitschek and R. Brooks, Sexual selection, sexual conflict and the evolution of ageing and life span - Functional Ecology, Volume 22 No 3 pp 443-453 ; British Ecological Society ; June 2008
- Les stratégies de reproduction des animaux : l’aventure évolutive de la sexualité, Thierry Lodé - Dunod, 2001.
- Écologie approche scientifique et pratique - Claude Faurie, Christiane Ferra, Paul Médori, Jean Dévaux et Jean-Louis Hemptinne - Tec & Doc Lavoisier 6e édition
- David Reznick, Michael J.Bryant et Farrah Bashey; r- and K-Selection revisited : the rôle of population regulation in life-history evolution - Ecology, 83(6), 2002, p. 1509-1520 © 2002 by the Ecological Society of America
- D. Pont et al., Stratégies démographique des poissons des rivières françaises : premiers résultats - An international journal on freshwater ecosystem (2013)
- Kevin R. Foster, Tom Wenseleers et Francis L.W. Ratnieks, Kin selection is the key to altruism - TRENDS in ecology and evolution Vol.21 No.2 février 2006.
- (en) A. Khila et E. Abouheif, « Reproductive constraint is a developmental mechanism that maintains social harmony in advanced ant societies », Proceedings of the National Academy of Sciences, vol. 105, no 46, , p. 17884–17889 (ISSN 0027-8424 et 1091-6490, PMID 19004767, PMCID PMC2584687, DOI 10.1073/pnas.0807351105, lire en ligne, consulté le )
- (en) Abderrahman Khila et Ehab Abouheif, « Evaluating the role of reproductive constraints in ant social evolution », Philosophical Transactions of the Royal Society B: Biological Sciences, vol. 365, no 1540, , p. 617–630 (ISSN 0962-8436 et 1471-2970, PMID 20083637, PMCID PMC2817144, DOI 10.1098/rstb.2009.0257, lire en ligne, consulté le )
Voir aussi
 Portail origine et évolution du vivant
Portail origine et évolution du vivant