Appareil respiratoire (anatomie des insectes)
L'appareil respiratoire des insectes est l'ensemble d'organes qui permettent les échanges de gaz respiratoires entre ces animaux et leur environnement. Les insectes sont des arthropodes essentiellement terrestres, dotés d'un système respiratoire spécialisé, adapté à la respiration aérienne. À l'opposé du système circulatoire des animaux couplé avec leur système respiratoire, ce dernier est distinct chez les insectes. Il est constitué d'un réseau ouvert très ramifié de voies respiratoires (trachées et trachéoles de calibres très divers[4], s'anastomosant plus ou moins)[5] qui assurent le transfert des gaz métaboliques et débouchent extérieurement au niveau des stigmates à ouverture variable, appelés spiracles.


Certains insectes minuscules et vivant en milieu humide ont une respiration uniquement tégumentaire. 3 % des insectes sont devenus secondairement aquatiques (en), la majorité, restés très engagés dans leurs voies évolutives respectives, ayant conservé leur système trachéen.
L'internalisation du système respiratoire et la présence d'un exosquelette chitineux protégeant efficacement de la dessiccation, sont des facteurs qui ont permis la réussite de la conquête du milieu terrestre par les insectes et favorisé la colonisation de nombreuses niches écologiques, à l'origine de leur succès évolutif[6], mais sont aussi responsables de la petite taille de ces arthropodes[7].
Insectes terrestres
Description
L'insecte, à la différence d'autres animaux (y compris certains arthropodes), n'est pas muni de poumons ni de branchies. Il respire par un système de trachées se divisant en trachéoles, et apportant directement l'air aux organes. Les trachées sont des invaginations cuticulaires tubuleuses[8] qui s'enfoncent dans le corps sous forme de gros troncs trachéens qui se ramifient en diminuant de diamètre, devenant des trachéoles qui transportent l'air sous forme gazeuse jusqu'aux cellules ou des ensembles de cellules consommatrices d'oxygène (tissus). Ces trachéoles se terminent, au contact de ces tissus, dans des cellules étoilées (cellules trachéolaires) sous forme de canalicules intracytoplasmiques contenant un liquide trachéolaire dans lequel se dissolvent les gaz respiratoires, ce qui est nécessaire pour leur diffusion transmembranaire. La fine paroi des trachées est rigidifiée par des épaississements spiralés ou annelés (ténidies ou taenidies) contribuant à éviter l'affaissement de ces tubes[9]. Les gènes régulateurs qui induisent la formation de ces trachées chez les insectes, « sont homologues des gènes qui induisent le développement des branchies chez les crustacés » : les trachées peuvent être ainsi considérées comme des replis branchiaux invaginés évitant la dessication lors de la terrestrialisation des arthropodes[3].
Les trachées prennent naissance à partir d'un spiracle qui est un orifice percé dans les sclérites (pleurites thoraciques et abdominaux) de l'exosquelette[9]. Ces orifices ont des structures filtrantes (soies hydrophobes) et des valves de fermeture, évitant les pertes en eau pour ces Arthropodes dont la conquête des terres a nécessité la protection des organes respiratoires contre le desséchement[10],[11].
Certains insectes bons voiliers (Hyménoptères, Diptères, Orthoptères) possèdent des sacs aériens trachéens, dilatations des gros troncs trachéens qui stockent une réserve d'air et permettent une diffusion en masse de l'oxygène[9].
Ventilation
Chez les insectes de petite taille au métabolisme peu élevé, la ventilation est passive, la circulation de l'air dans les trachées résultant d'un simple phénomène de diffusion. Chez ceux de taille importante ou au métabolisme élevé, elle est active, impliquant des cycles de contraction et de relâchement des muscles abdominaux, dorso-ventraux et longitudinaux, ainsi que des cycles d'ouverture et d'obturation des stigmates[9]. L'abdomen se rétracte grâce à l'action des muscles internes (myofibrilles) afin de faire sortir l'air vicié du système de trachées. L'expiration est donc active, mais l'inspiration est un processus passif, car la chitine de la cuticule est élastique, et reprend sa forme une fois l'action des muscles finie[3]. « Beaucoup d'insectes peuvent pratiquer une ventilation dite discontinue. Dans ce système, les spiracles restent fermés pendant un certain temps, pouvant varier de quelques minutes à plusieurs heures suivant l'espèce et le niveau d'activité de l'animal. Pendant cette phase, le CO2 s'accumule dans les trachées alors que la teneur en oxygène diminue largement. Cette phase est suivie d'une phase de "flutter[12]", en général courte, pendant laquelle les spiracles s'ouvrent et se ferment très rapidement. Il y a ensuite une phase d'ouverture pendant laquelle l'essentiel des échanges gazeux s'effectue[13] ».
Avec l'augmentation de taille, le rapport entre la surface intérieure des tubes et le volume du corps rend ce système respiratoire inefficace, ce qui serait un des facteurs expliquant la petitesse des insectes. En effet, plus l'insecte est grand, plus il a besoin d'oxygène[14]. Or ce système trouve ses limites. Il n'a pas de pompe efficace, comme c'est le cas dans la respiration pulmonaire, actionnée par les muscles thoraciques et le diaphragme. Cependant, l'hypothèse qui a longtemps prévalu selon laquelle le transport des gaz se faisait par diffusion (mécanisme peu efficace) est désormais nuancée : de nombreux insectes ventilent le système trachéal assurant un transport des gaz par convection. Cette ventilation résulte de la compression de certaines parties du système trachéal par différents mouvements musculaires, par déplacement de l'hémolymphe entre les parties du corps provoquée par le renversement des battements cardiaques, et par des cycles rythmiques rapides des trachées en lien avec l'ouverture et la fermeture des spiracles[15].
Insectes aquatiques
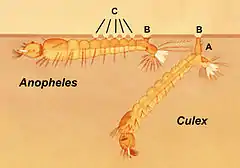
3 % des insectes sont devenus secondairement aquatiques mais ils ont généralement conservé leur système de trachées[17].
Quelques petits ordres (éphéméroptères, odonates, plécoptères, trichoptères) restés très engagés dans leurs voies évolutives respectives, ont des larves devenues dulçaquicoles, grâce à la mise en œuvre d'organes temporaires (trachéobranchies qui permettent une totale et durable immersion dans la pleine réalisation d'une écophase aquatique) adaptés à ce milieu de colonisation. Chez des petits groupes d'hétéroptères (Hydrocorises : Naucores, Nèpes, Notonectes…) ou de coléoptères (certains Hydrophilidés et Dytiscidés), les espèces sont devenues, à l'état adulte comme à l'état larvaire, « essentiellement ou complètement dulçaquicoles, bien que conservant une respiration aérienne typique, mais contraignante — à l'instar, globalement, des grands Mammifères devenus marins —, et avec des accessoires dédiés ou des modalités particulières[18] ». Ces insectes présentent typiquement cinq adaptations : respiration aérienne (par exemple grâce à un siphon respiratoire de surface), respiration « aérienne » sous l'eau (par capture de bulles d'air au contact vers de la cuticule, siphonnage d'oxygène dans les lacunes aérifères des plantes aquatiques, comme chez les Donacia et Coquillettidia), respiration transtégumentaire (l'oxygène dissout diffuse à travers la cuticule et rejoint l'hémolymphe ou les trachées qui ne s'ouvrent pas sur l'extérieur, par exemple larve de chironome), branchies sanguines qui fonctionnent selon le même principe que les branchies des autres animaux (par exemple larve de chironome dotée de branchies au niveau de papilles ventrales, remplies d'hémolymphe qui contient de l'hémoglobine dissoute) ou trachéobranchies (l'O2 ne diffuse pas dans l'hémolymphe, mais dans des trachées)[19].
Chez ces espèces qui possèdent des trachéo-branchies (appelées aussi branchies trachéennes), l'appareil respiratoire terrestre s'est adapté à la vie aquatique, grâce à la constitution d'expansions tégumentaires très fines au niveau des stigmates, dans lesquelles se trouvent des ramifications de trachées (les trachéoles dont la paroi est très fine et perméable aux gaz) diffusant l'air. Ces expansions latérales, dont le but est d’augmenter la surface d'échange avec le milieu extérieur, fonctionnent comme des branchies, d'où leur nom de trachéo-branchies. Elles sont le plus souvent lamelleuses, foliacées, pectinées, digitées ou filiformes[20]. Les branchies trachéennes peuvent être externes (branchies abdominales chez les larves d'Odonates zygoptères et d'Éphéméroptères ; trachéobranchies prosternales — sous le cou —, coxales — au niveau de l’insertion des pattes sur le thorax — ou anales — à l'extrémité de l'abdomen — chez les Plécoptères ; certains Névroptères, Trichoptères,Culicidés) ; ou internes (trachéobranchies rectales chez les larves d'Odonates anisoptères)[21].
| Larves aquatiques | ||||||||||||||||
| ||||||||||||||||



.jpg.webp)

.jpg.webp)


| Insectes aquatiques | |||||||||
| |||||||||


Notes et références
- Une réduction du nombre de spiracles s'observe, en relation avec la modification du système trachéen, chez certains insectes immatures, insectes aquatiques, ou avec la spécialisation des segments terminaux chez les adultes.
- Une au-dessus de chaque paire de pattes
- Lauralee Sherwood, Hillar Klandorf et Paul Yancey, Physiologie animale, De Boeck Superieur, , p. 504
- Calibres qui vont du millimètre au micromètre.
- Ce réseau est dense chez les insectes ancestraux. Les trachées s’anastomosent chez les insectes plus évolués, formant plusieurs réseaux (réseau dorsal : muscles des ailes et cœur ; réseau médian : viscères ; réseau ventral : muscles et chaîne nerveuse) qui améliorent l'efficacité du système respiratoire.
- Jeanne Raccaud-Schoeller, Les insectes : physiologie, développement, Masson, , p. 3.
- Peter H Raven, Susan R Singer, Georges B Johnson, Kenneth A Mason, Jonathan B Losos, Biologie, De Boeck Superieur, (lire en ligne), p. 965.
- Lorsque l'insecte mue, la cuticule chitineuse tapissant les parois internes du réseau de trachées est renouvelée elle aussi
- Sandrine Heusser et Henri-Gabriel Dupuy, Atlas de biologie animale, Dunod, , p. 87
- R. F. Chapman, Stephen J. Simpson, Angela E. Douglas, The Insects. Structure and Function, Cambridge University Press, , p. 501
- (en) P D Srivastava et R. P. Singh, An introduction to entomology, Concept Publishing Company, , p. 96
- Par analogie avec le flutter atrial.
- Raymond Gilles et Michel Anctil, Physiologie animale, De Boeck Supérieur, , p. 211
- Pour un facteur d'agrandissement K, la surface d'échange (la consommation d'oxygène possible, pour un type de système respiratoire donné) est multipliée par K2 et le volume (donc la masse et la consommation) par K3.
- (en) M.W. Westneat, O. Betz, R.W. Blob, K. Fezzaa, W.J. Cooper, W.-K. Lee, « Tracheal respiration in insects visualized with synchrotron X-ray imaging », Science, 299 (2003), p. 558-560.
- Helena da Cunha Ramos, Jacques Brunhes, Insecta Diptera Culicidae Uranotaenia, Quae, , p. 408-409
- Alain Ramel, « La respiration aquatique des insectes », Insectes, no 146, , p. 3
- Michel Boulard, Métamorphoses et transformations animales. Oblitérations évolutives, Société nouvelle des éditions Boubée, , p. 254
- Denis Poinsot, Maxime Hervé, Bernard Le Garff, Mael Ceillier, Diversité animale. Histoire, évolution et biologie des Métazoaires, De Boeck Superieur, , p. 336
- Roger Dajoz, Dictionnaire d'entomologie, Lavoisier, , p. 163
- Pierre-Paul Grassé, Traité de zoologie, Masson, , p. 146
- (de) Gerhard von Frankenberg, « Die Schwimmblasen von Corethra. Biologische und morphologische Studien über ihre Funktion und Beiträge zur Physiologie des geschlossenen Tracheensystems », Zool. Jahrb. Allg. Zool., vol. 35, , p. 505-592
- (en) S. Teraguchi, « Correction of negative buoyancy in the phantom larva, Chaoborus americanus », Journal of Insect Physiology, vol. 21, no 10, , p. 1659 (DOI 10.1016/0022-1910(75)90176-6)
- (de) Gerhard von Frankenberg, « Die Schwimmblasen von Corethra. Biologische und morphologische Studien über ihre Funktion und Beiträge zur Physiologie des geschlossenen Tracheensystems », Zool. Jahrb. Allg. Zool., vol. 35, , p. 505-592
- (en) S. Teraguchi, « Correction of negative buoyancy in the phantom larva, Chaoborus americanus », Journal of Insect Physiology, vol. 21, no 10, , p. 1659 (DOI 10.1016/0022-1910(75)90176-6)
- Trachéobranchies filamenteuses à la base des pattes, formées de nombreuses houppes ramifiées.
- Trachéobranchies filamenteuses à la base des pattes, formées de nombreuses houppes ramifiées.
- Le CO2 est éliminé par les stigmates abdominaux, formant des bulles sous les élytres, finalement expulsées en arrière par des mouvements abdominaux. Henri Bertrand, Les insectes aquatiques d'Europe, P. Lechevalier, , p. 504.
- Le CO2 est éliminé par les stigmates abdominaux, formant des bulles sous les élytres, finalement expulsées en arrière par des mouvements abdominaux. Henri Bertrand, Les insectes aquatiques d'Europe, P. Lechevalier, , p. 504.
- Alain Ramel, op. cit., p.4
- Alain Ramel, op. cit., p.4
Voir aussi
Bibliographie
- (en) J. Buck, « Some physical aspects of insect respiration », Annu. Rev. Entomol., 7 (1962), p. 27-59
Articles connexes
Liens externes
- Respiration des insectes – simulation, animation interactive…
- Méthodes pour respirer dans l'eau chez les insectes aquatiques
 Portail de l’entomologie
Portail de l’entomologie