Forme transitionnelle
Une forme transitionnelle est une espèce fossile présentée comme à la fois descendante d'un groupe d'espèces connues et ancêtre d'un autre groupe d'espèces connues. Elle présente une mosaïque de caractères plésiomorphes et apomorphes des deux autres groupes d'espèces. Les trois groupes formeraient ensemble un segment d'une lignée évolutive. Les formes transitionnelles ne doivent pas présenter d'autapomorphies.
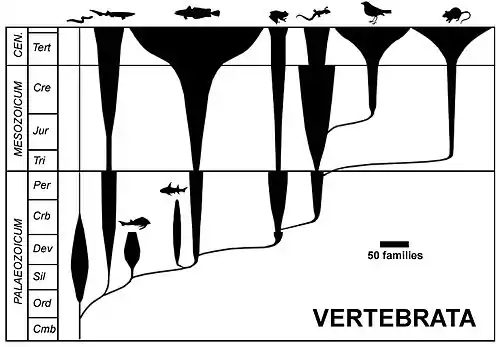
Selon les systématiciens cladistes[3], cette expression est à bannir comme le sont ses équivalents de « chainon manquant » ou de « maillon ». Pour les cladistes, ces termes sont des métaphores qui évoquent la Scala naturæ[4], « grande chaine de la vie » marquée par sa dimension téléologique et anthropocentrique[5]. Cet avis n'est pas partagé par les systématiciens évolutionnistes qui utilisent des techniques d'analyse des données permettant de placer certains fossiles directement sur une lignée évolutive[6].
Une notion débattue
Les cladistes soutiennent généralement l'idée qu'il n'est pas possible d'identifier une forme transitionnelle comme telle. Ils s'appuient pour cela sur les principes méthodologiques de la cladistique qui ne permettent pas de placer une espèce directement sur une branche de l'arbre du vivant mais seulement à ses extrémités : « La phylogénie n'exprime pas de notion d'ancêtre à descendant (qui descend de qui) mais une notion de parenté (qui est plus proche de qui)[7] ». Cela implique qu'un fossile jugé transitionnel sera placé dans une position de groupe frère d'une autre espèce, et non comme ancêtre de celle-ci[8]. Les principes du cladisme n'admettent pas de filiation entre taxons mais seulement des parentés plus ou moins proches, et n'admettent pas la validité taxonomique des groupes paraphylétiques. Seuls sont admis les groupes incluant obligatoirement tous les descendants d'un ancêtre commun. Les cladistes soutiennent qu'il est très improbable qu'une forme fossile identifiée soit réellement la forme transitionnelle recherchée.
Les évolutionnistes contestent ces affirmations et y voient de simples axiomes, posés a priori et sans fondement empirique[9],[10]. Au contraire, des statisticiens ont estimé que 63 % des espèces actuelles avaient au moins une espèce ancestrale encore vivante[11]. Cela signifie que de très nombreuses espèces actuelles descendent l'une de l'autre et que nous ne devrions pas les considérer comme étant toujours des espèces-sœurs, mais bien souvent comme des espèces-mères et des espèces-filles. Une espèce-mère vivante formerait donc de facto à la fois un groupe naturel observable et un groupe paraphylétique, alors que les cladistes considèrent que seuls les groupes monophylétiques sont valides d'un point de vue taxonomique. Les biologistes évolutionnistes critiquent la vision cladistique en ce qu'elle ne prend pas en compte la flèche du temps, ni d'éventuels sauts évolutifs (à cause d'une évolution tachytélique) qui seraient suffisants pour justifier l'exclusion taxonomique d'un groupe du clade auquel il appartient cladistiquement. Ils admettent donc certains groupes paraphylétiques dès lors qu'ils représentent un grade évolutif. Certains paléontologues ont de plus développé de leur côté des méthodes, notamment bayésiennes, permettant de placer des espèces fossiles directement sur les branches d'un cladogramme et non plus seulement à leurs sommets[6].
L'image du chainon manquant reste très populaire et est couramment utilisée dans les médias, notamment lors de découvertes de nouvelles espèces fossiles. Il convient de remarquer que si l'on trouve une forme intermédiaire X soupçonnée d'avoir existé entre A et B, on ne se retrouve plus avec une absence de chainon manquant, mais dorénavant avec deux : une entre A et X et une seconde entre X et B. La quête peut de ce fait continuer presque indéfiniment, rendue d'autant plus difficile par le fait que les organismes fossiles présentent souvent une mosaïque de caractères ancestraux et dérivés et que les caractères apomorphes peuvent masquer le « substratum » de caractères plésiomorphes[3]. Pour éviter toute ambiguïté, des cladistes comme Guillaume Lecointre préfèrent l'emploi du terme d'intermédiaire structural à celui de forme transitionnelle[3].
Exemples emblématiques
Évolution humaine
Dans le langage courant, les mots « singe » et « homme » désignent des catégories très différentes, et il est habituel de parler de genre humain en l'opposant au règne animal, englobant l'ordre des Primates.
À partir du deuxième ouvrage majeur de Charles Darwin en 1871, La Filiation de l'homme et la sélection liée au sexe, il a été avancé par les savants de l'époque que « l'homme descendait du singe ». On a recherché alors les fossiles d'êtres intermédiaires qui illustreraient cette vision. La quête du « chainon manquant » a poussé à de nombreuses recherches et continue de le faire aujourd'hui. C'est ainsi que le médecin anatomiste néerlandais Eugène Dubois a découvert en 1891 à Java des fossiles qui semblaient répondre à cette quête, associant bipédie moderne et crâne archaïque, et qu'il a nommé en 1894 Pithecanthropus erectus, devenu par la suite Homo erectus.
On sait depuis 1967 que les chimpanzés sont génétiquement plus proches des humains que des gorilles. L'Homme et le Chimpanzé descendent d'un ancêtre commun, remontant à au moins 7 millions d'années et qui reste inconnu à ce jour. De la même façon, les gorilles sont plus proches des humains et des chimpanzés que des orang-outans. De ce point de vue, l'Homme fait partie des singes plutôt qu'il n'en descend.
Certains anthropologues argüent toutefois que cette vision déforme notre conception de l'évolution[12]. De plus, les chercheurs savent que la nature ne se plie qu'approximativement aux catégories schématiques de la science[13], et que par exemple « les frontières des espèces sont plus floues » qu'on l'imagine souvent.
Les formes fossiles ayant précédé l’Homme moderne commencent aujourd'hui à être mieux connues. Une des difficultés de la paléoanthropologie est de comprendre comment se sont enchainées ces multiples espèces, sachant qu'on ne peut présenter aujourd'hui aucune filiation consensuelle qui puisse relier deux quelconques de ces formes.
Évolution des oiseaux
.jpg.webp)
Une des espèces fossiles d'archéoptéryx
L'un des exemples les plus célèbres de forme transitionnelle est celui de l'Archaeopteryx, la première forme découverte jugée transitionnelle entre dinosaures et oiseaux. Cette espèce présente des caractères dérivés propres aux oiseaux (vol ailé à plumes) et des caractères ancestraux propres aux dinosaures (dents)[14]. Cette mosaïque de caractères a conduit à considérer l'archéoptéryx comme un chainon manquant entre dinosaures et oiseaux. Comme tous les organismes vivants présentent des mosaïques de caractères ancestraux et de caractères dérivés, les cladistes considèrent que l'Archaeopteryx est une espèce fossile située à l'extrémité d'une branche éteinte. Les évolutionnistes mettent généralement en avant l'absence d'autapomorphie pour défendre sa nature d'espèce ancestrale.
Pour les cladistes les oiseaux ne sont donc qu'un groupe de dinosaures parmi d'autres, ce qui signifie que ces derniers n'ont pas tous disparu lors de l'extinction Crétacé-Paléogène d'il y a 66 millions d'années, mais ont en partie survécu jusqu'à l'époque actuelle. L'analyse phylogénétique montre par exemple que les tyrannosaures sont bien plus proches parents de l'archéoptéryx et des oiseaux actuels que des tricératops[14]. Ainsi, pour eux, de la même manière que l'homme ne descend pas du singe mais est un singe, les oiseaux ne descendent pas des dinosaures mais sont des dinosaures[15]. Les évolutionnistes ne contestent pas ces résultats mais insistent sur les divergences morphologiques marquées entre ces deux taxons et sur la rapidité de cette transition évolutive pour exclure taxonomiquement les oiseaux des dinosaures[10]. Des paléontologues spécialistes des dinosaures pointent également que ce détournement volontaire des noms des taxons paraphylétiques et la confusion entre les verbes être et descendre de génèrent un obscurcissement sémantique[16].
Dans les œuvres de fiction
Le roman Les Animaux dénaturés de Vercors, dont fut tirée la pièce Zoo ou l'assassin philanthrope, est fondé sur l'hypothèse suivante : le chaînon manquant n'est pas fossile mais encore vivant. Se pose alors une question que les paléontologues considèrent comme sans objet quand il s'agit d'espèces éteintes : s'agit-il d'un être protégé par la Déclaration universelle des droits de l'homme, puisqu'il se trouve, par nature, à mi-chemin entre l'humain et le singe[17] ? L'homme adulte utilise le plus souvent de 15 000 à 30 000 mots, les chimpanzés peuvent en maîtriser 200 en langue des signes. Que faire si le vocabulaire de ces êtres est de 1 000 mots ? Vercors décide de prendre comme critère d'hominisation l'interrogation métaphysique qui fut commune à Homo sapiens et à l'Homme de Néandertal. À cette aune, une partie de la tribu découverte est humaine et une autre, qui semble ne se poser aucune question de ce genre, ne l'est pas.
Le roman Le Père de nos pères, de Bernard Werber, propose plusieurs théories sur l'évolution plus ou moins fictives. Le chaînon manquant se révèle être l'union d'un chimpanzé mâle et d'un phacochère femelle.
Dans le film Man to Man, des paléontologues soutiennent que les pygmées sont le chaînon manquant.
Notes et références
- Avec les cinq principaux clades représentés : Agnathes (lamproies), Chondrichthyens (requins, raies), Placodermes (fossiles), Acanthodiens (fossiles), Osteichthyens (poissons osseux).
- « Systématique : ordonner la diversité du vivant », Rapport sur la Science et la technologie N°11, Académie des sciences, Lavoisier, 2010, p. 65
- Guillaume Lecointre et Hervé Le Guyader, 2006, Classification phylogénétique du vivant, Éd. Belin, troisième édition, augmentée et corrigée ; (ISBN 2-7011-4273-3)
- Jean Piveteau, Des premiers vertébrés à l'homme, Albin Michel, , p. 152.
- Guillaume Lecointre et Hervé Le Guyader, Classification phylogénétique du vivant, Belin, , p. 17
- Gavryushkina, A., Welch, D., Stadler, T. et Drummond, A. J., « Bayesian Inference of Sampled Ancestor Trees for Epidemiology and Fossil Calibration », PLoS computational biology, vol. 12, no 10, , e1003919
- Guillaume Lecointre et Hervé Le Guyader, Classification phylogénétique du vivant, Belin, , p. 46
- (en) Bapst David W., « When Can Clades Be Potentially Resolved with Morphology? », PLoS ONE, vol. 8, no 4, , e62312
- (en) Richard H. Zander, A Framework for Post-Phylogenetic Systematics, Saint Louis (Missouri), Zetetic Publications,
- Damien Aubert, Classer le vivant : Les perspectives de la systématique évolutionniste moderne, Ellipses, (ISBN 9782340017733)
- Aldous, D. J., Krikun, M. A. et Popovic, L., « Five statistical questions about the tree of life », Systematic biology, vol. 60, no 3, , p. 318-328
- Marks, J., Tales of the Ex-Apes : How We Think about Human Evolution, University of California Press, , 240 p.
- Sommes-nous humains ?, Léo Grasset
- R.O. Prum et A.H. Brush : « Les plumes de dinosaures ». Pour la Science (juillet-aout 2005), dossier Le Monde des Dinosaures.
- G. Lecointre et H. Le Guyader 2006 : Classification phylogénétique du vivant. Éd. Belin (3e éd), (ISBN 2-7011-4273-3)
- Dodson, P., « Origin of Birds: The Final Solution? », Integrative and Comparative Biology, vol. 40, no 4, , p. 504–512
- Voir Paradoxe sorite
Voir aussi
 Portail origine et évolution du vivant
Portail origine et évolution du vivant  Portail de la paléontologie
Portail de la paléontologie  Portail de la Préhistoire
Portail de la Préhistoire