Chronobiologie
La chronobiologie est une discipline scientifique étudiant l’organisation temporelle des êtres vivants, des mécanismes qui en assurent la régulation (contrôle, maintien) et de ses altérations. Cette discipline traite essentiellement de l’étude des rythmes biologiques.

Histoire
Premières observations
L’Homme préhistorique acquiert déjà une connaissance sommaire de l’organisation temporelle des êtres vivants (maturité des fruits, migration du gibier, frai des saumons, etc.). L’Homme du néolithique maîtrise l’agriculture et l’élevage par sa connaissance du cycle végétal et du cycle reproducteur des animaux[2]
Les premiers écrits décrivant les rythmes biologiques concernent la biologie végétale. Ils remontent au IVe siècle av. J.-C. : Théophraste rapporte dans son Histoire des plantes qu’Androsthène observe sur l’île de Tylos un arbre « dont les indigènes disent qu’il dort » : ce photopériodisme concerne probablement le tamarinier[3].
Au XVIIe siècle, le médecin italien Santorio Santorio met en évidence le rythme circadien chez l’Homme en mesurant la variation journalière de son poids.
Premières expérimentations et applications
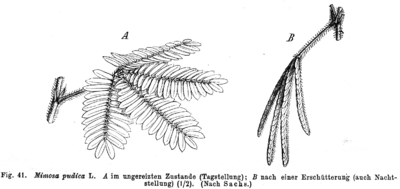
A. Feuilles en « position de veille » le jour.
B. Feuilles en « position de sommeil » la nuit.
En 1729, le savant français Jean-Jacques Dortous de Mairan étudie la nyctinastie chez la sensitive : même placée dans l’obscurité totale et dans un environnement constant (température, humidité), la plante continuait d’ouvrir ses feuilles (comme elle le fait pendant le jour) et les replier la nuit. Il expérimente ainsi pour la première fois les rythmes circadiens et montre ainsi leur nature endogène[4]. En 1751, le naturaliste suédois Carl von Linné applique ce phénomène de nyctinastie pour concevoir une horloge florale. En 1814, le médecin Julien Joseph Virey (1755-1836) publie Éphémérides de la vie humaine, ou Recherches sur la révolution journalière et la périodicité de ses phénomènes dans la santé et les maladies, première thèse de chronopharmacologie dans laquelle il pose la terminologie « horloge du vivant »[5]. En 1832, Augustin Pyrame de Candolle découvre que la nyctinastie de la sensitive s’exerce sur une périodicité de 22 à 23 heures, montrant l’existence d’une période endogène en cours libre. Il réalise aussi la première expérience de resynchronisation biologique en exposant la sensitive à l’obscurité le jour et à un éclairage permanent la nuit[6].
En 1910, l'entomologiste Auguste Forel est le premier à mettre en évidence une horloge interne chez les animaux : observant que les abeilles étaient attirées par la confiture à chaque fois qu'il petit-déjeunait sur la terrasse de son chalet, il nota, par un jour de mauvais temps, qu'elles revenaient à la même heure sur sa terrasse alors qu'il prenait son petit-déjeuner à l'intérieur et qu'elles ne pouvaient la sentir[7],[8]. En 1911, l’éthologiste allemand Karl von Frisch, en étudiant le contrôle photique de la pigmentation cutanée d'un poisson, le Vairon, découvre un mécanisme qu'il nomme « photoréception extra-oculaire », cette photoréception contrôlée par la glande pinéale jouant un rôle important dans la photorégulation physiologique et la synchronisation métabolique[9]. À partir de 1914, il porte toutes ses recherches sur l’abeille et montre avec son étudiante Ingeborg Beling (en) que l'insecte dispose d’une horloge interne, avec trois mécanismes de synchronisation ou de réglage[10]. En 1915 dans son ouvrage Contributions à la connaissance sur l'origine des mouvements de sommeil[11], le botaniste Wilhelm Pfeffer est le premier à émettre l'hypothèse d'une horloge interne autonome [12]. En 1920, les botanistes américains Whigtman Garner et Henry Allard font une étude approfondie sur le photopériodisme et classent un grand nombre de plantes en jours courts et longs[13]. En 1925, le biophysicien russe Alexander Chizhevsky (en) établit une relation entre les tempêtes solaires et les catastrophes sur terre (guerres, épidémies, meurtres). Il fonde l’héliobiologie qui sera plus tard intégrée à la chronobiologie[14].
Recherche contemporaine
Les premiers laboratoires scientifiques étudiant les oscillations biologiques se mettent en place dans les années 1920[15]. En 1935, le biologiste allemand Erwin Bünning montre l’origine génétique du rythme circadien chez des plantes[16].
Les travaux exhaustifs de Jürgen Aschoff, Erwin Bünning et Colin Pittendrigh (en) dans les années 1950 sur les horloges circadiennes des oiseaux et souris, font qu’ils sont considérés comme les fondateurs de la chronobiologie. Franz Halberg, de l’université du Minnesota, qui a étudié l’influence de l’heure d’administration des médicaments et inventé le mot circadien en 1959, est considéré comme le « père de la chronobiologie américaine ». En France, c’est Alain Reinberg qui fait figure de pionnier.
En 1960, le symposium à Cold Spring Harbor Laboratory (en) jette les bases pour le domaine de la chronobiologie. La même année, Patricia DeCoursey invente la Phase response curve (en) (courbe de réponse de phase), un des principaux outils utilisés dans le domaine[17].
Dans les années 1970, le premier gène de l’horloge, nommé per (pour period est mis en évidence dans le règne animal (drosophile[18], en 1971 chez le rongeur), d'autres gènes de ce type sont identifiés dans le règne végétal (algue Chlamydomonas reinhardtii[19]), fongique (Neurospora crassa[20]).
Des expériences « hors du temps » (isolement temporel selon le protocole de libre cours[21]) sont menées par les biologistes allemands Jürgen Aschoff et Rutger Wever (1962) et par Michel Siffre (en 1962 et 1999) : elles montrent que diverses fonctions humaines (physiologiques, cognitives ou comportementales) sont contrôlées par une horloge circadienne de période endogène en cours libre (24h et 9 minutes en moyenne : 24 h 5 min chez les femmes, 24 h 11 min chez les hommes, ce qui explique qu'en moyenne les femmes ont besoin de se coucher plus tôt et sont plus sujettes aux insomnies que les hommes[22]).
En 1992, Michael Rosbash met en évidence des horloges circadiennes au niveau moléculaire (ARN messager de per)[23]. En 1997, une étude révèle que la majorité des cellules possède une horloge moléculaire indépendante[24]. En 2005, une horloge d'une cyanobactérie est reconstituée dans un tube à essai[25].
Si la chronobiologie actuelle s'intéresse à la génétique et aux niveaux moléculaires (par exemple les travaux du docteur James Bendayan qui étudie les différences de rythmicité des génomes différents chez les femmes et les hommes[3]) elle porte également son attention sur les impacts des rythmes biologiques dans un référentiel calqué sur la vie quotidienne des êtres humains et des sociétés, au travers de la chronomédecine, de la variation de la performance humaine (sports, cognition), de la chronobiologie appliquée[26] où plus récemment de la chronoprévention des risques (influences du travail posté et du travail de nuit sur la santé au travail, analyse et couverture des risques, problématiques de santé publique)[27]
Chronobiologie et homéostasie
Bien que l’idée du facteur temps en biologie et en médecine ne soit pas nouvelle (notion que l’on retrouve chez Aristote et Pline qui constatent la rythmicité dans la reproduction, la floraison, l’hibernation ou la migration), les réflexions, recherches et pratiques de ces dernières décennies ont longtemps été influencées par une croyance en l’invariance des êtres vivants sur le « court terme », à l’échelle des 24 heures, tout comme à l’échelle d’une année.
Certains parlent à cet égard de dogme en visant plus ou moins directement le concept d’homéostasie, que l’on retrouve chez Walter Cannon s’inspirant des idées sur la stabilité du milieu intérieur de Claude Bernard.
La contradiction entre le sujet d’étude de la chronobiologie et ce concept n’est qu’apparente et est probablement due à une mauvaise interprétation.
En effet, l’homéostasie traite de la capacité qu’a le milieu intérieur d’un être vivant à se maintenir dans un état apparemment ou globalement stable et ce malgré les fluctuations et changements survenant au sein de son environnement. Or ce dernier n’est jamais constant, ses caractères perceptibles évoluent sans cesse :
- de manière rythmique, facilement prévisibles (la Terre tourne sur elle-même et autour du Soleil, ce qui induit une alternance lumière/obscurité ainsi que la présence de saisons) ;
- de manière aléatoire ce qui est parfois beaucoup plus subtil à percevoir et à prévoir.
L’effet de fluctuations rythmiques (comme l’alternance jour/nuit sur 24 h, ou jours courts / jours longs sur une année) sur un organisme qui se veut homéostatique induit logiquement une compensation du même ordre en vue du maintien de l’organisme observé. Ces rétrocontrôles ou feed-backs réguliers permettent donc l’équilibre d’un état de « non-équilibre ».
La chronobiologie s’inscrit à ce titre dans le cadre de l’étude des processus non linéaires, que l’on retrouve en thermodynamique chez des chercheurs comme Prigogine ou en science des systèmes. Elle traite donc d’oscillations des systèmes ouverts et évolutifs.
Selon Alain Reinberg[28], de nombreux chronobiologistes s’accordent à dire que, globalement, les rythmes biologiques correspondent à une adaptation des êtres vivants aux variations prévisibles de l’environnement. La question du « Pourquoi ? » des rythmes biologiques reste toutefois « embarrassante » : selon l’auteur, tenter d’y répondre correspondrait à introduire la question de la finalité, et plus précisément celle des mécanismes de l’évolution des êtres organisés, de leur adaptation spécifique (relative à l’espèce) et individuelle à l’environnement. Dans cette situation il est donc difficile de fournir des « preuves expérimentales » de ce que l’on avance. Les rythmes biologiques peuvent donc apparaître comme une « condition » de la survie des individus ou d’une espèce dans la périodicité de l’environnement terrestre. Il faut toutefois remarquer qu’il existe certains rythmes qui ne semblent pas correspondre de prime abord à une nécessité environnementale.
Le concept d’homéostasie doit donc impérativement intégrer les notions de dynamique et de biopériodicité. La notion d’équilibre en biologie, lorsque cet équilibre n’est pas dynamique (un déséquilibre perpétuellement rattrapé), est synonyme de mort.
Caractérisation des rythmes biologiques
Un rythme biologique se caractérise par sa période, l’emplacement de l’acrophase (ou pic, ou sommet, ou zénith) de la variation dans l’échelle de temps de la période, l’amplitude et le niveau moyen de la variation (MESOR).
Période
Intervalle de temps mesuré entre deux épisodes qui vont se reproduire identiques à eux-mêmes au cours de la variation. La période du rythme d’une variable biologique peut être obtenue par analyse spectrale, fournissant une estimation de la période prépondérante fondamentale et de ses harmoniques. On peut aussi l’obtenir via la connaissance du rythme des synchroniseurs (conditions expérimentales).
En fonction de la période prépondérante, la chronobiologie distingue trois grands domaines de rythmes :
- les rythmes circadiens, d’une période équivalant théoriquement à un jour (24 heures), mais qui varie en réalité de 20 à 28 heures ;
- les rythmes ultradiens, c’est-à-dire d’une fréquence plus rapide qu’un rythme circadien, donc d’une durée théoriquement inférieure à 24 heures ;
- les rythmes infradiens, c’est-à-dire d’une fréquence plus lente qu’un rythme circadien, donc d’une période supérieure à 24 heures. Parmi ceux-ci :
- les rythmes septénaires (environ une semaine),
- les rythmes circamensuels (environ un mois),
- les rythmes circannuels, ou saisonniers.
Une même variable biologique manifeste sa rythmicité dans plusieurs de ces domaines (exemple du cortisol plasmatique).
Acrophase
L’acrophase (pic, ou zénith), dont l’opposé est la « batyphase » ou « bathyphase », est la position de la plus haute valeur de la variable biologique mesurée dans l’échelle du temps, pour la période considérée en fonction d’une référence temporelle. Lorsque l’on se trouve dans le domaine circadien, le pic peut être donné en heures avec comme référence une heure (par exemple minuit de l’heure locale). Il est possible de donner l’emplacement de l’acrophase par rapport à la température corporelle, mais cela reste beaucoup plus rare.
Lorsqu’on utilise la méthode du Cosinor, le pic sera le point le plus élevé de la fonction sinusoïdale, mais la plupart du temps on parle de pic au regard des valeurs expérimentales.
Amplitude
La caractérisation est la même qu’en sciences physiques ou en mathématiques. Elle représente la variation totale de la valeur biologique mesurée sur la période considérée.
Mesor ou niveau moyen du rythme
MESOR pour Midline Estimating Statistic Of Rythm. Il s’agit de la moyenne arithmétique des mesures de la variable biologique.
Propriétés des rythmes biologiques
Les rythmes biologiques ont une origine à la fois endogène et exogène :
Origine endogène
Leur origine est génétique, ils sont innés et ne résultent pas d’un apprentissage individuel. Ils sont gouvernés par des horloges biologiques (ou garde-temps). Cette caractéristique peut être mise en évidence par une isolation (protocole de libre cours) durant laquelle les rythmes persistent sur une fréquence qui leur est propre.
Ces facteurs endogènes sont entraînés par des facteurs exogènes, les Zeitgebers ou synchroniseurs.
L’origine endogène prend son origine de la constitution génétique de l’espèce et de ses individus. Il est possible qu’interviennent d’une part des gènes programmant directement le rythme considéré et d’autre part la structure d’ensemble de l’individu dépendant à la fois de l’ensemble des autres données génétiques et de facteurs socio-psycho-biologiques exogènes.
On connaît une horloge principale localisée dans l’hypothalamus et des horloges secondaires dont plusieurs sont gérées, elles aussi au niveau cérébral.
Il existe plusieurs gènes codant diverses horloges biologiques : on a, par exemple, décrit une horloge alimentaire qui réglerait la préparation digestive au repas à venir (cf. Étienne Challet et al., Current Biology du 24 octobre 2006).
Rythmes d'origine centrale et rythmes d'origine périphérique
En fait, toutes les cellules de l’organisme, et pas seulement celles qui appartiennent aux structures cérébrales plus spécialisées, sont dotées d’une horloge propre qui est difficile à mettre en évidence in vitro dans les conditions habituelles du laboratoire. Benoît Kornmann et ses collaborateurs ont découvert la possibilité de laisser en activité ou d’annihiler l’horloge de cellules hépatiques ; cela a permis de déterminer que leur rythme circadien est à 90 % d’origine « locale » mais qu’il existe un impact « global » (central et/ou lié directement aux synchroniseurs externes) de 10 % au moins. Cette part est très robuste et persiste lorsqu’on bloque l’horloge propre des cellules périphériques.
Facteurs d'entraînements exogènes, ou synchroniseurs
Le synchroniseur est un facteur environnemental, parfois social, mais toujours périodique, susceptible de modifier la période ou la phase d’un cycle biologique. Les synchroniseurs ne créent pas les rythmes biologiques mais ils en contrôlent la période et la phase.
Les principaux agents d’entraînement des rythmes chez l’homme sont de nature cognitive, ainsi les indicateurs socio-écologiques y jouent un grand rôle.
On peut citer ici l’alternance activité/repos, lumière/obscurité au niveau quotidien, ou encore la photopériode (jours courts / jours longs) et la température au niveau annuel ou saisonnier.
Conclusions et implications
Les rythmes biologiques sont donc entraînables (ajustement de la période des rythmes) mais aussi persistants (mise en évidence par protocoles de free run ou libres cours, dans lesquels on coupe l’individu de tous signaux susceptibles de le resynchroniser).
On peut déplacer leurs phases par induction via la manipulation des synchroniseurs (lumière essentiellement) et ainsi créer des avances ou des retards de ces phases, on peut ainsi en cas de pathologie remettre à l’heure l’horloge biologique et ainsi remettre en phase l’organisation temporelle de l’individu. Les rythmes circadiens, quasiment ubiquitaires, sont peut-être les rythmes biologiques les plus remarquables et les plus facilement observables.
D’autres synchroniseurs – sociaux notamment – s’adressent à notre cortex. Ils sont des signaux et peuvent être appris. Grâce à un travail cérébral spécifique, tout signal perçu comme repère temporel peut devenir un synchroniseur et orienter notre « vécu » circadien, mais aussi, le cas échéant, circannuel, ultradien, etc. Autrement formulé, notre « horlogerie » interne est influencée par le bruit des voisins, le déclenchement de la sonnerie du réveil, l’heure de passage du facteur, le moment quotidien pendant lequel telle personne a pris l’habitude de nous téléphoner — la liste est longue. Chez l’homme, les synchroniseurs sociaux ont un effet plus important que les synchroniseurs naturels, mais on observe des phénomènes semblables chez certains animaux sociaux qui se synchronisent grâce aux informations données par leurs congénères. Un synchroniseur social peut en remplacer un autre par un phénomène d’apprentissage.
Désynchronisation
La désynchronisation correspond à une perte de la relation de phase des rythmes biologiques. Elle peut être d’origine externe (liée aux modifications de l’environnement) ou interne (sans relation directe avec l’environnement).
Désynchronisation externe
Travail posté
Le travail de nuit ou le travail posté peuvent provoquer une désynchronisation de l’organisation temporelle de l’individu (il est difficile de prédire qui est tolérant ou non à ce type de travail).
Décalage horaire ou jet lag
En cas de vol transméridien supérieur à environ cinq heures (phénomène de décalage horaire) on observe une désynchronisation chez les individus.
- Rythme nycthéméral : recadrage en 2 jours ;
- Température du corps : recadrage en une semaine ;
- Sécrétion du cortisol : recadrage en 15 à 20 jours.
Cécité totale
Les aveugles dont la rétine est complètement inopérante (la rétine contient des récepteurs non photiques permettant de stimuler la sécrétion de mélatonine par la glande pinéale) présentent de nombreux troubles de leur organisation temporelle. La lumière ne pouvant pas être traduite en signal hormonal de synchronisation, il s’ensuit des symptômes similaires à ceux pouvant apparaître dans d’autres cas de désynchronisation.
Désynchronisation interne
Cette dernière est mal comprise. Elle est affectée par l’âge, la dépression, ou les cancers hormono-dépendants (sein, ovaires, prostate, etc.).
Mise en évidence d'une désynchronisation
On peut la mettre en valeur via l’étude de rythmes marqueurs (cortisol plasmatique, mélatonine plasmatique, température, etc.). Si la désynchronisation est mise en évidence, ces marqueurs seront dits soit en avance de phase, soit en retard de phase par rapport à l’organisation temporelle de référence (normale) pour l’individu étudié.
Désynchronosation et perte de poids
L’horloge circadienne périphérique des tissus adipeux
L’horloge centrale, soit celle située dans les noyaux suprachiasmatiques (NSC), régule les horloges périphériques par entrée neuronale directe. Par contre, les entrées sympathiques seules ne peuvent être responsable de toutes les activités circadiennes. Il y a présence d’une horloge dans plusieurs organes liés à l’apport alimentaire comme l’estomac, l’intestin, le pancréas ainsi que le foie. Un changement dans l’alimentation peut influencer par entraînement neurohumoral les mécanismes des horloges circadiennes périphériques. De plus, la présence d'une horloge circadienne active dans le tissu adipeux suggère qu'il existe une composante temporelle à la régulation de la fonction des tissus adipeux. L’horloge circadienne dans l’adipocyte modifie la sensibilité de ce dernier à des stimuli spécifiques différents au cours de la journée de 24 heures tels l’insuline ou encore l’adrénaline. L’horloge des adipocytes peut aussi modifier la capacité de stockage des triglycérides comme la périlipine. Une asynchronie entre le sommeil et l’alimentation altère l’horloge circadienne de l’adipocyte et cette altération serait responsable de l’augmentation de l’adiposité. Une modification dans l’horaire d’alimentation peut aussi modifier la phase d’expression de gènes avec un rythme circadien jusqu’à 12h sans affecter la phase de l’expression circadienne dans le NSC. Dans ce cas, il y a donc désaccouplement des horloges périphériques avec l’horloge centrale. Ce réajustement de phase fait par les horloges périphériques suite à un changement dans l’horaire alimentaire se produit rapidement dans le foie, mais est plus lent dans les reins, le cœur et le pancréas.
Impact du moment de la prise alimentaire sur la leptine
Bien que les mécanismes reliant la synchronisation des repas et la prise de poids soient encore méconnus, il semble que les hormones y aient leur rôle à jouer. L'expression rythmique et l'activité des voies métaboliques sont principalement attribuées à la robustesse et l'expression coordonnée des gènes de l’horloge dans les différents organes et tissus. Or, les changements dans le moment de l'apport calorifique peuvent altérer ce bien construit et modifier la rythmicité de nombreuses hormones impliquées dans le métabolisme, telles que la leptine ou encore la ghréline. En fait, les études réalisées en laboratoire ont montré que les moments durant lesquels les souris étaient éveillées et en train de manger au cours de leur nuit biologique, (c’est-à-dire le jour dans le cas des souris, puisque ce sont des animaux nocturnes) a entraîné de multiples changements métaboliques. Cela comprend notamment une modification de la sécrétion de leptine, une hormone anorexigène qui procure le sentiment de satiété à l’organisme en inhibant les neurones NPY / AgRP et en activant les neurones POMC / CART. Plus précisément, il a été montré que les valeurs plasmatiques de leptine nocturne étaient significativement diminuées. Habituellement, c’est-à-dire lorsque l’apport alimentaire est fait durant le jour biologique (la nuit pour les souris), la sécrétion de leptine par les tissus adipeux est faite proportionnellement aux réserves lipidiques. Or, plus les réserves sont élevées, plus la sécrétion de l’hormone est élevée, ce qui signifie une augmentation de la sensation de satiété et une diminution de l’apport calorifique. Par conséquent, les variations quotidiennes de l'apport alimentaire jouent directement sur la sécrétion de leptine puisqu’elle augmente après l'alimentation et diminue pendant le jeûne. Maintenant, le problème avec les souris nourries durant la nuit biologique est que le taux de leptine étant significativement inférieur, le sentiment de satiété est moindre contrairement aux souris nourries la nuit. Ainsi, la faible quantité de l’anorexigène tend à favoriser l’augmentation de l’apport calorifique par jour et ce, bien que les besoins énergétiques restent inchangés. Ce serait ainsi une cause de la prise de poids. C’est d’ailleurs un phénomène qui est aussi présent chez l’homme[réf. nécessaire].
Impact du déphasage de la corticostérone et de l’insuline
Comme mentionné précédemment, le moment auquel l’apport calorifique est effectué a un effet direct sur la sécrétion d’hormones, dont la corticostérone fait partie. Il a été montré expérimentalement qu’une souris habituellement nourrie la nuit et dont le rythme alimentaire est bouleversé par l’apport de ses repas le jour, montre un haut pic de corticostérone lors des repas. Il est donc suggéré que cette hormone serait liée invariablement à la prise inhabituelle de poids. En effectuant une ablation de la glande adrénale, responsable de la production de corticostérone, il est maintenant possible d’observer une perte de poids. Cela serait dû au fait que la corticostérone augmente la lipogénèse et l’accumulation de gras abdominal. La lipogénèse, se faisant normalement lorsque l’animal est en grande période d’activité, limite la création de gras abdominaux. Par contre, si elle se produit durant une phase peu active de la journée, son effet est grandement augmenté.
L’insuline est également responsable de la prise de poids chez l’homme. Une expérience se faisant sur des rats a montré ses effets. Chez des rats ayant six repas par jour répartis sur une période de 24h, le niveau de glucose dans le sang était très régulier durant la période de lumière. Par contre, les deux repas donnés pendant la nuit engendraient un niveau d’insuline beaucoup plus haut. Ainsi, le glucose sanguin s’est vu largement diminué par l’effet de l’insuline. Les rats normalement actifs la nuit et mangeant à cette période voient leur métabolisme complètement bouleversé lorsqu’ils mangent en journée. Le glucose sanguin devient alors plus élevé, ce qui amène à des risque d’augmenter la masse adipeuse de l’individu.
Impact du déphasage sur le métabolisme des souris
La désynchronisation des divers éléments humoraux du métabolisme présentés plus haut est impliquée dans le découplement de deux paramètres importants du métabolisme : le ratio d’échange respiratoire (RER) et la dépense énergétique des cellules. En situation de restriction alimentaire à la nuit subjective plutôt que durant le jour subjectif, le RER subit un déphasage d’environ 10 heures dans les cellules hépatiques. Cela signifie que la prise de nourriture a un impact plus important dans les cellules du foie que l’horloge centrale. En situation de nutrition sans restriction, le RER fluctue de façon à synchroniser les moments où l’énergie provient des réserves de gras ou des intrants provenant de l’alimentation. Ici, la restriction de nourriture lors de la nuit subjective augmente la valeur moyenne de ce paramètre sur toute la période de 24 heures, indiquant que moins de réserves de gras sont sollicitées par la dépense énergétique. Le jour, la dépense énergétique vient en grande partie de l’activité musculaire, qui repose sur l’utilisation des carbohydrates (fournis directement par l’alimentation) imposée par une valeur élevée du RER (au-dessus de 1). Celui-ci étant moins sensible au zeitgeber alimentaire, son déphasage se situe donc entre 5 et 7 heures, ce qui le désynchronise d’avec le déphasage du RER du foie (10h). De plus, la dépense énergétique est environ 9% plus faible durant toute la période de 24 heures, signifiant qu’en plus de ne pas être en phase avec les pics d’utilisation des carbohydrates, moins d’énergie est dépensée par l’organisme . Ainsi, garder plus de gras stocké chez les souris nourries durant leur nuit subjective, couplé à une diminution des dépenses énergétiques, entraîne une augmentation du nombre de cellules adipeuses des individus. La désynchronisation des rythmes du foie, qui fournit une grande partie de l’énergie durant la nuit, entraîne une utilisation moins efficace du glycogène et un fort débalancement de l’homéostasie énergétique du corps, pouvant avoir des impacts sur la prise de poids.
Pour conclure, plusieurs hypothèses sont émis selon lesquelles le moment auquel l’apport alimentaire est effectué aurait des impacts sur la prise de poids. En effet, le changement dans le taux de sécrétion d’hormone telles que la leptine, la ghréline, la corticostérone et l’insuline, ainsi que la diminution de l’activité physique de l’individu semble être les causes principales de ce gain. Par contre, le mécanisme précis qui explique le lien direct entre ces composantes et la prise de poids reste encore inconnu et ce, bien que le lien entre ceux-ci et l'embonpoint soit prouvé.
Autres facteurs pouvant affecter les rythmes biologiques
L’âge est un facteur dont il faut tenir compte :
- le fœtus est cosynchronisé avec les rythmes de sa mère[réf. nécessaire]
- le nourrisson a ses rythmes qui seront plutôt portés sur l’ultradien (cycle activité/repos de 50 min à une heure en rapport avec la maturité du système nerveux ?)[29]
- l’enfant de 4 ans est totalement circadien[réf. nécessaire]
- le stade pubertaire change les rythmes biologiques[réf. nécessaire]
- la personne âgée aura des rythmes de moins en moins bien synchronisés et « marqués »[réf. nécessaire]
Le sexe : la notion de rythme chez la femme est moins facile à étudier que chez l’homme (cycles menstruels).[réf. nécessaire]
La surface corporelle joue également.[réf. nécessaire]
Exemples d'applications
En France, Michel Siffre, spéléologue, a réalisé l'une des premières expérience d’isolement hors temps dans le gouffre du Scarasson[30], du 18 juillet au à 2 000 m d’altitude dans les Alpes italiennes (entre Limone et Tende).
Les conditions de cette expérimentation peuvent se rapprocher des conditions de free run, situation dans laquelle les individus étudiés sont privés de tous synchroniseurs. Le free run permet de mettre en valeur les périodes des rythmes endogènes de chaque individu.
Le Pr Christian Poirel (Canada) a étudié les rythmes circadiens de la souris et sur les phénomènes psychopathologiques humains. [réf. souhaitée]
Chronopsychologie
En 1967, dans Psychologie du temps, Paul Fraisse crée et développe la notion de chronopsychologie.
François Testu (université de Tours), a étudié les rythmes d’apprentissage chez l’enfant, en leur faisant faire des exercices simples et en regardant les taux de réussite selon les heures[31]. Il a observé la présence de deux acrophases, vers 11 h et 17 h 30 (acrophase qui n’existe pas chez les petits enfants), et de deux batyphases, la première vers 13 h 30 (elle n’est pas directement et uniquement liée à la digestion du déjeuner, sinon il y aurait également une batyphase durant toute période post-prandiale, après toute prise d’aliments). Elle dure environ 2 heures, (entre 13 h et 15 h). Cette baisse est très liée à la baisse physiologique de la vigilance correspondant au creux méridien[32]. La deuxième a lieu vers 3 h 30 du matin.[réf. nécessaire] Claire Leconte s’étonne de voir un tel résultat sur les rythmes d’apprentissage chez l’enfant : est-il réveillé la nuit pour faire une épreuve d’attention[33] ? Cette dernière est sans doute liée à la chute de la température, qui est au plus bas entre 3 et 5 heures du matin[3].
Outre ce cycle circadien d’attention, on note aussi un cycle ultradien d’environ 90 minutes, ce que Kleitman appelle le BRAC (Basic Rest-Activity Cycle, Cycle fondamental activité-repos)[29]. Par exemple après le début d’un cours, l’attention est à son maximum après environ 25 minutes, puis décroît et la batyphase se situe vers 75 minutes[réf. nécessaire]. Aucune recherche n’a permis de confirmer un tel résultat, la variation de l’attention lors d’un cours d’une heure est très dépendante du contenu de ce cours, de la compétence de l’enfant par rapport à l’activité à réaliser, de la motivation que cet enfant éprouve pour ce cours, du contexte pédagogique dans lequel il est fait. Dans les expériences menées, on relève de grandes différences inter-individuelles[32].
Une étude américaine a révélé un cycle d’attention correspondant à l’intervalle entre les publicités qui coupent les émissions télévisées[réf. nécessaire].
Rôle dans l'accidentologie
Alain Reinberg[34], en citant Folkard[35], insiste sur la place de la chronobiologie en accidentologie et donne quelques raisons :
- L’accident a une rythmicité à l’échelle d’une population. « Il est unique et peut être mortel pour l’individu, mais le regroupement de son incidence en fonction du temps montre qu’il existe des heures noires »[36]. Selon l’auteur, l’intervalle des heures noires se situe entre minuit et quatre heures du matin (mais les frontières sont légèrement floues, dues à la variabilité biologique et aux synchronisations respectives des individus concernés).
- Le caractère nocturne de l’accident de l’adulte est une expression des rythmes circadiens affectant directement la vigilance et la performance des activités des individus[37],[38],[39],[40],[41].
Ces variations de vigilance sont très étudiées dans le cas de surveillance du pilotage des navires (organisation en quarts) ou de salles de contrôles d'installation industrielles (usines chimiques, centrales nucléaires) ou de trafic (tour de contrôle, Cross). Des catastrophes industrielles de l'époque moderne se sont produites au cœur de la nuit, à un moment de vigilance moindre ; on peut citer l'exemple célèbre du naufrage du Titanic qui s'est produit durant la période critique aux alentours de 23 h et 1 h du matin.
Justice et management
En 2011, une étude sur l’impartialité de la justice a montré que les libérations sur parole accordées par les tribunaux varient de 65 % (après une restauration) à pratiquement zéro relaxe obtenue avant la pause déjeuner[42].
Recherche sur l'horloge biologique
L'horloge biologique est déjà largement pressentie au XVIIIe siècle. En 1729 l'astronome français Jean-Jacques Dortous de Mairan cite ainsi comme exemple la feuilles du Mimosa se fermant au crépuscule et s'ouvrant à l'aube (même quand elle est conservée [vivante] dans l'obscurité)[43].
Parallèlement à la découverte de l'importance de la mélatonine, ce n'est qu'au XXème siècle que le mécanisme génétique et moléculaire commence à être expliqué. Seymour Benzer et Ronald Konopka au California Institute of Technology de Pasadena créent dans les années 1970 des drosophiles mutantes présentant une horloge biologique anormale et montrent que ces mutations et anomalies proviennent d'un même gène muté qu'ils dénommeront avec d'autres le gène « Period »[43] (qui sera séquencé en 1984).
En 2017 le travail sur les mécanismes de l'horloge biologique (identification des gènes impliqués dans le rythme circadien, chez la drosophile) réalisé par trois chercheurs américains : Jeffrey Hall, Michael Rosbash et Michael Young a été récompensé par le prix Nobel de médecine[44]. En 1984 M. Rosbash avait isolé un gène dit "period" contrôlant le rythme biologique circadien. Avec Jeffrey Hall il a montré que la protéine PER (codée par le gène period) est accumulée dans les cellules avec un pic la nuit puis dégradée le jour. Puis en 1994 Michael Young montre qu'un autre gène dit « timeless » code une protéine dite « TIM » indispensable au déroulement du rythme circadien, TIM se liant à PER pour entrer dans le noyau de la cellule et bloquer l'activité du gène period (rétrocontrôle négatif)[45]. Ce principe a été détecté chez la drosophile, mais ensuite retrouvé dans les cellules de nombreuses autres espèces, dont Homo sapiens[43].
Notes et références
- (en) Michael Smolensky, Lynne Lamberg, The Body Clock Guide to Better Health: How to Use Your Body's Natural Clock to Fight Illness and Achieve Maximum Health, Henry Holt and Company,
- Ladislas Robert, Les Temps de la vie, Flammarion, , p. 129.
- Claire Leconte, professeur émérite de psychologie, La chronobiologie, émission La Tête au carré sur France Inter, 12 avril 2011.
- Jean-Jacques Dortous de Mairan, Observation botanique, Histoire de l’Académie royale des sciences, , p. 35-36.
- Alain Reinberg, L’Art et les secrets du temps : une approche biologique, du Rocher, , p. 33-34.
- Augustin Pyrame de Candolle, Physiologie végétale, Bechet Jeune, .
- Françoise Macar, Le temps : perspectives psychophysiologiques, éditions Mardaga, , p. 29
- (en) Jeremy Rifkin, Time Wars : The Primary Conflict in Human History, Simon & Schuster, , p. 39
- Jean Boissin et BernardCanguilhem, Les rythmes du vivant : origines et contrôle des rythmes biologiques, Éditions du CNRS, , p. 212-213)
- (en) Adrian Horridge, What Does the Honeybee See?, ANU E Press, , p. 21
- (de) Wilhelm Pfeffer, Beiträge zur Kenntnis der Entstehung der Schlafbewegungen, B. G. Teubner, , 154 p.
- (en) Fritz-Albert Popp, L.V. Beloussov, Integrative Biophysics : Biophotonics, Springer, , p. 45
- (en) Garner & Allard, « Effect of the relative length of day and night and other factors of the environment on growth and reproduction in plants », Journal of Agriculture Research, no 18, , p. 553-606.
- (en) Edward S. Ayensu, Philip Whitfield, The Rhythms of life, Marshall Publishing Limited, .
- [PDF] La synchronisation et la cadence.
- (de) Erwin Bünning, « Zur Kenntnis der erblichen Tagesperioizitat bei den Primarblatter von Phaseolus multiflorus », Jahrbücher für wissenschaftliche Botanik, no 81, , p. 411-418.
- (en) Leon Kreitzman et coll, Rhythms of life : the biological clocks that control the daily lives of every living thing, Yale University Press,
- (en) Konopka, R.J., Benzer, S., « Clock mutants of Drosophila melanogaster », Proceedings of the National Academy of Sciences, vol. 68, , p. 2112-2116
- (en) Bruce, V.G., « Mutants of the biological clock in Chlamydomonas reinhardi », Genetics, vol. 70, , p. 537-548
- (en) Feldman, J.F., Hoyle, M., « Isolation of circadian clock mutants of Neurospora crassa », Genetics, vol. 755, , p. 605-613
- Par opposition au protocole de désynchronisation forcée
- (en) Jeanne F. Duffy et coll, « Quantification of Behavior Sackler Colloquium: Sex difference in the near-24-hour intrinsic period of the human circadian timing system », Proceedings of the National Academy of Sciences, vol. 108, , p. 15602-8 (DOI 10.1073/pnas.1010666108)
- (en) M. Rosbash et coll, « Circadian oscillations in period gene mRNA levels are transcriptionally regulated », Proceedings of the National Academy of Sciences, no 89, , p. 89:11711-11715.
- (en) Liu C., Weaver D.R., Strogatz S.H., Reppert S.M., « Cellular construction of a circadian clock: period determination in the suprachiasmatic nuclei », Cell, no 91, , p. 855-860
- (en) M. Nakajima et col, « Reconstitution of circadian oscillation of cyanobacterial KaiC phosphorylation in vitro », Science, no 308, , p. 414-415
- Voir les colloques thématiques de l'International Society for Chronobiology: http://www.chronoint.org/55431/ICACC-MEETING
- Cette approche est celle de certains sapeurs-pompiers français. Voir Brousse E, Forget C, Riedel M, Marlot M, Mechkouri M, Smolensky MH, Touitou Y, Reinberg A., 24-hour pattern in lag time of response by firemen to calls for urgent medical aid, in Chronobiology International, 2011 Apr;28(3):275-81, https://www.ncbi.nlm.nih.gov/pubmed/21452923
- REINBERG, A., (1991), Dimension temporelle de la médecine, in Chronobiologie médicale, chronothérapeutique, Flammarion, coll. Médecine Sciences, 2e édition (2003), Paris, p. 8-9.
- Nathaniel Kleitman, Sleep and wakefulness, University of Chicago Press, 1963.
- http://www.cms06.com/pagesperso-orange.fr/c.m.speleo-nice/albums/index5.htm Photos du site de l’expérience de Michel Siffre dans le gouffre du Scarasson.
- François Testu, Rythmes de vie et rythmes scolaires : aspects chronobiologiques et chronopsychologiques, Masson, , 175 p. (lire en ligne).
- Claire Leconte-Lambert et Pierre Leconte, La Chronopsychologie, PUF, coll. « Que sais-je ? », , 2e éd., 128 p. (ISBN 978-2130433316)
- Claire Leconte, Des rythmes de vie aux rythmes scolaires : une histoire sans fin, Septentrion, coll. « Que sais-je ? », , 2e éd., 270 p. (ISBN 978-2-7574-0583-3)
- REINBERG A. (2003), Heures noires, Rythmes du risque des accidents, in Chronobiologie médicale, chronothérapeutique, Flammarion, coll. Médecine Sciences, 2e édition, Paris, p. 263-273.
- FOLKARD S., Black times: temporal determinents of transport safety, Accid. Anal and Prev, 1997, 29: 417-430.
- REINBERG A. (2003), op. cit., p. 263.
- LAVIE P., The search for cycles in mental performance from Lombard to Kleitmann, in Chronobiologia, 1980,7:247-258.
- COLQUHOUN W.P., Biological rythms and human performance, Londres, Academic Press, 1971:39-107.
- GILLOOLY P, SMOLENSKY M.H., ALBRIGHT D., et al., Circadian variations in human performance evaluated by the Walter Reed Assessement Battery, Chronobiology Int, 1990, 7: 143-153.
- CARRIER J, MONK T, Effects of sleep and circadian rythms on performance. In FW Turek, PC Zee, Regulation of sleep and circadian rythms. New York, Marcel Dekker Inc. 1999 527:556.
- MONK T., Shift worker performance, In: AJ Scott. Shiftwork. Occupationnal medecine. Philadelphie, Hanley & Belfus Inc. 1990:183-198.
- « Comment notre cerveau décide », Shai Danziger, chercheur au sein de l’université Ben Gourion (Israël), La Recherche no 473, mars 2013, p. 46
- Vogel G & Stokstad E (2017) Timing is everything: U.S. trio earns Nobel for work on the body’s biological clock ; Science News ; 2 octobre 2017
- vidéo de l'annonce du Nobel de médecine 2017, sur le site du prix Nobel
- LB Vosshall, JL Price, A Sehgal, L Saez, MW Young (1004) Block in nuclear localization of period protein by a second clock mutation, timeless] 18 mars 1994 | Vol. 263, Issue 5153, pp. 1606-1609 | DOI: 10.1126/science.8128247|résumé
- autres publications
- Buijs ,R. M., van Eden, C. G., Goncharuk, V. D., & Kalsbeek, A. (2003). The biological clock tunes the organs of the body: timing by hormones and the autonomic nervous system. J Endocrinol, 177, 17 –26.
- Kobayashi, H., Oishi, K., Hanai, S., & Ishida, N. (2004). Effect of feeding on peripheral circadian rhythms and behaviour in mammals. Genes Cells, 9, 857–864.
- Zvonic, S., Ptitsyn, A.A., Conrad, S. A., Scott, L. K., Floyd, Z. E., Kilroy, G., Wu, X., Goh, B. C., Mynatt, R. L., & Gimble, J. M. (2006). Characterization of Peripheral Circadian Clocks in Adipose Tissues. Diabetes, 55(4), 962-970.
- Bray, M. S., & Young, M. E. (2007). Circadian rhythms in the development of obesity: potential role for the circadian clock within the adipocyte. Obesity Reviews, 8(2), 169-181.
- Damiola F., Le Minh, N., Preitner, N., Kornmann, B., Fleury-Olela, F., & Schibler, U. (2000). Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev, 14, 2950–2961.
- Arble DM, Bass J, Laposky AD, Vitaterna MH, Turek FW. (2009). Circadian timing of food intake contributes to weight gain. Obesity (Silver Spring); 17: 2100–2.
- Scheer FA, Hilton MF, Mantzoros CS, Shea SA. (2009). Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc Natl Acad Sci U S A ;106: 4453–8.
- Gomez Abellan P, Gomez Santos C, Madrid JA, Milagro FI, Campion J, Martinez JA, et al. (2011). Site-specific circadian expression of leptin and its receptor in human adipose tissue. Nutr Hosp; 26:1394–401.
- Garaulet, M., & Ordovás, J. M. (2013). Genetics in chronobiology and obesity. In Chronobiology and obesity (pp. 37-40; 66-68; 75-80). Springer New York.
- Gorman MR (2003). Differential effects of multiple short day lengths on body weights of gonadectomized siberian hamsters. Physiol Biochem Zool 76:398–405
- Salgado-Delgado, R., Angeles-Castellanis, M., Saderi, N., Buijs, R. M., Escobar, C. (2010) Food Intake during the Normal Activity Phase Prevents Obesity and Circadian Desynchrony in a Rat Model of Night-club Work. Endocrinologue, Volume 37, Issue 3.
- Kalsbeek, A., Strubbe, J. H. (1998) Circadian Control of Insulin Sécrétion Is Independent of the Temporal Distribution of Feeding. Physiologie & Béhaviorisme, Volume 63, Issue 4, p.553-560.
- Bray, M. S., Ratcliffe, W. F., Grenett, M. H., Brewer, R. A., Gamble, K. L., & Young, M. E. (2013). Quantitative analysis of light-phase restricted feeding reveals metabolic dyssynchrony in mice. International journal of obesity, 37(6), 843-852.
- Reznick, J., Preston, E., Wilks, D. L., Beale, S. M., Turner, N., & Cooney, G. J. (2013). Altered feeding differentially regulates circadian rhythms and energy metabolism in liver and muscle of rats. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease, 1832(1), 228-238.
Voir aussi
Bibliographie
- (en) Dunlap J.C., Loros J.J., DeCoursey P.J. dir., Chronobiology, biological time keeping, Sinauer Associates Inc., 2004, Sunderland,
- Albert Goldbeter, La Vie oscillatoire. Au cœur des rythmes du vivant, éditions Odile Jacob, 2010. Présentation en ligne
- André Klarsfeld, Les Horloges du vivant. Comment elles rythment nos jours et nos nuits, éditions Odile Jacob, 2009. Présentation en ligne
- Leconte, C. (1995) La chronopsychologie à l'école, In Manuel de psychologie pour l'enseignement, coord. D. Gaonac’h et C. Golder, éd. Hachette éducation, 456-489.
- Christian Poirel, Les Rythmes circadiens en psychopathologie (Perspectives neurobiologiques sur les structures de rythmes temporalité), Masson éd., Paris, 1975.
- Alain Reinberg, (1991), Chronobiologie médicale, chronothérapeutique, Flammarion, coll. Médecine Sciences, 2e édition (2003), Paris
- Alain Reinberg, F. Levi et M. Smolensky, « Chronobiologie et pathologie infectieuse »/« Chronobiology and infectious diseases », Médecine et Maladies infectieuses, vol. 17, supplément 2, mai 1987, pages 348-350 ; doi:10.1016/S0399-077X(87)80286-X
- Sechter, D. et Poirel, C., Chronobiologie et psychiatrie, Masson Publ., Paris et New York, 1985.
Articles connexes
- Chronobiology International
- Horloge circadienne
- Horloge moléculaire
- Alain Reinberg
- Yvan Touitou
Liens externes
- International Society for Chronobiology
- Société francophone de chronobiologie (SFC)
- Synchroniseurs cellulaires périphériques
- (histoire des sciences) Aux origines de la chronobiologie, texte d'A. Klarsfeld commentant une « observation botanique » (Mairan 1729), en ligne sur BibNum.




