Système immunitaire inné
Le système immunitaire inné comprend les cellules et les mécanismes permettant la défense de l'organisme contre les agents infectieux de façon immédiate[1] car elle ne nécessite pas de division cellulaire, à l'inverse du système immunitaire adaptatif qui confère une protection plus tardive mais plus durable[2] et qui nécessite une division cellulaire (lymphocyte B et T). Alors que le système immunitaire adaptatif existe uniquement chez les gnathostomes, le système immunitaire inné existe dans tous les organismes du règne végétal et animal.
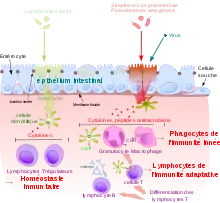
Bien qu'elle ne soit pas spécifique d'une espèce particulière d'agent pathogène, elle peut reconnaître des types de pathogènes comme les bactéries Gram négatif qui sont reconnues par les récepteurs[3] appelés pattern recognition receptor (PRR) ou en français récepteur de reconnaissance de motifs moléculaires.
Les principales fonctions du système immunitaire inné des chordés sont :
- la constitution d'une barrière physique et chimique contre les agents infectieux ;
- la détection des agents infectieux et le recrutement de cellules immunitaires sur le site de l'infection ;
- l'activation de la cascade du complément permettant l'activation des cellules et l'élimination de cellules mortes ou de complexes immuns ;
- l'identification et l'élimination de corps étrangers présents dans l'organisme, les tissus, le sang et la lymphe, par les globules blancs ;
- l'activation de l'immunité adaptative à travers la présentation de motif moléculaire associé aux pathogènes fixés aux récepteur de reconnaissance de motifs moléculaires.
Barrière anatomique
L'épithélium est une barrière physique constituant l'une des premières défenses contre les agents infectieux[4]. La desquamation de l'épiderme permet ainsi d'éliminer des bactéries ou autres agents infectieux ayant adhéré à la surface de l'épithélium. Dans le tube digestif et les voies respiratoires, le mouvement opéré par le péristaltisme ou les cils contribuent à l'élimination des agents infectieux. La flore intestinale prévient la colonisation par des bactéries pathogènes en agissant notamment par compétition pour les nutriments. Les larmes et la salive contribuent également à prévenir l'infection des yeux et de la bouche, respectivement.
| Barrière anatomique | Mécanismes correspondants |
|---|---|
| Peau | Transpiration, desquamation, acides organiques |
| Tube digestif | Péristaltisme, mucus, acide gastrique, acide biliaire, enzymes digestives, thiocyanate, défensines, flore intestinale |
| Voies respiratoires et poumons | Mucus, surfactant pulmonaire, défensines |
| Rhinopharynx | Mucus, salive, lysozyme |
| Yeux | Larmes |
Cellules du système immunitaire inné
Les différents types de cellules sanguines se distinguent par leurs protéines de surface : ces différents types de cellules, en immunologie, sont regroupés en clusters de différenciation (CD). Ces clusters sont reconnus par des anticorps monoclonaux et les clusters sont nommés CD1, CD2, CD3, etc. selon qu'on utilise un anticorps monoclonal de type 1, 2 ou 3[5].
Les leucocytes, ou globules blancs, se distinguent des autres cellules de l'organisme en cela qu'elles ne sont pas associées à un organe ou un tissu en particulier. Les leucocytes peuvent se déplacer librement dans l'organisme et capturer des débris cellulaires, des corps étrangers, ou des microorganismes invasifs.
Les cellules immunitaires innées comprennent les cellules NK, les granulocytes (mastocytes, polynucléaires neutrophiles, éosinophiles et basophiles) et les phagocytes (macrophages, polynucléaires neutrophiles et cellules dendritiques), et agissent en identifiant et en éliminant les agents infectieux[6].
Mastocytes
Les mastocytes sont un type de cellules immunitaires innées résidant dans les tissus conjonctifs et les muqueuses. Ces cellules contribuent à la défense contre les agents infectieux et à la cicatrisation, mais sont également associées à l'allergie et au phénomène d'anaphylaxie[7]. À la suite de leur activation, les mastocytes relarguent rapidement des granules caractéristiques enrichies en histamine et en héparine, ainsi que différents médiateurs hormonaux et des chimiokines. L'histamine induit la vasodilatation, causant ainsi les différents signes caractéristiques de l'inflammation, et recrute des neutrophiles et des macrophages[7].
Macrophages
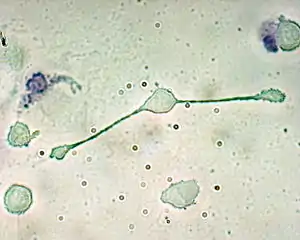
Les macrophages (du grec macro-, gros et -phagein, manger), sont de grosses cellules qui sont différenciées dans les tissus à partir des monocytes circulant dans les vaisseaux sanguins. La reconnaissance de motifs microbiens par les récepteurs situés à la surface des macrophages conduit à la phagocytose et à la destruction des agents infectieux par le processus d'explosion oxydative (oxidative burst en anglais) et la production de radicaux libres de l'oxygène. Les macrophages produisent également des chimiokines, permettant le recrutement d'autres cellules sur le site de l'infection[6].
Cellules dendritiques
Les cellules dendritiques sont des cellules présentes au niveau des muqueuses et qui sont donc parmi les premières cellules exposées à l'environnement extérieur. Elles sont présentes dans l'épiderme (où elles sont appelées cellules de Langerhans), dans les poumons, et dans l'intestin[2]. Les cellules dendritiques jouent un rôle essentiel dans la présentation antigénique et l'activation de l'immunité adaptative. Ces cellules tirent leur nom des dendrites neuronales.
Neutrophiles
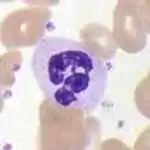
Les neutrophiles font partie, avec les éosinophiles et les basophiles, des cellules granulocytes ou cellules polynucléaires (voir ci-dessous). Les granules présents dans le cytoplasme des neutrophiles contiennent des substances toxiques permettant l'élimination des microorganismes extracellulaires comme les bactéries ou les champignons. Comme pour les macrophages, l'activation de l'explosion oxydative (oxidative burst) par les neutrophiles conduit à la production de radicaux libres de l'oxygène et la sécrétion de peroxyde d'hydrogène et d'hypochlorite. Les neutrophiles sont les phagocytes les plus nombreux, constituant 50 % à 60 % de l'ensemble des leucocytes circulants, et sont parmi les premières cellules recrutées au site d'infection[7]. La moelle osseuse d'un individu sain adulte produit environ 100 milliards de neutrophiles par jour, et environ 10 fois plus au cours d'une infection aigüe[7].
Basophiles et éosinophiles
Les basophiles et les éosinophiles sont apparentés aux neutrophiles (voir ci-dessus). La sécrétion d'histamine par les basophiles joue un rôle important dans la réponse immunitaire contre les parasites, et contribue aux réactions allergiques comme l'asthme[6]. Les éosinophiles sécrètent de nombreux composés toxiques et des radicaux libres permettant également l'élimination des bactéries et des parasites, mais qui peuvent occasionner des lésions tissulaires[7].
Monocytes
Les monocytes sont des leucocytes circulant dans le sang, ce sont des précurseurs de plusieurs populations de cellules dendritiques et de macrophages tissulaires, et ils occupent donc à ce titre une place prépondérante dans la mise en place des réponses immunitaires.
Cellules tueuses naturelles
Les cellules tueuses naturelles (ou cellules NK, pour Natural Killer), sont des cellules du système immunitaire inné cytotoxique qui n'éliminent pas directement les agents infectieux. Découvertes dans les années 1960, ces cellules tirent leur nom du fait qu'elles n'ont pas besoin de l'activation d'un lymphocyte CD4+ pour avoir une action cytotoxique[8],[9].
Ce sont des cellules provenant de la lignée lymphocytaire proche des cellules des lymphocytes T mais ne possédant pas de récepteur membranaire aux antigènes comme les lymphocytes. Les cellules tueuses naturelles éliminent les cellules dont la fonction est altérée, comme les cellules tumorales ou les cellules infectées par un virus. Par exemple, les cellules tueuses naturelles peuvent reconnaître les cellules n'exprimant plus le CMH de classe I. Cette situation est observée durant une infection virale, certains virus pouvant induire la diminution du CMH de classe I pour éviter la reconnaissance par d'autres cellules immunitaires comme les lymphocytes T CD8[10].
Lymphocytes T γδ
Comme d'autres cellules, les lymphocytes T NK ou les cellules lymphoïdes innées, les lymphocytes T γδ ont des caractéristiques de cellules de l'immunité innée et de l'immunité adaptative. D'un côté, les lymphocytes T γδ peuvent être considérés comme faisant partie de l'immunité adaptative en cela qu'ils ont opéré une recombinaison V(D)J ; de l'autre, la diversité de recombinaison est relativement restreinte et s'apparente ainsi à un récepteur de reconnaissance de motifs moléculaires. Ainsi, les lymphocytes T Vγ9/Vδ2 répondent rapidement à des antigènes non peptidiques exprimés par les agents infectieux, et les lymphocytes T intraépithéliaux expriment la chaîne Vδ1 et réagissent au stress cellulaire.
Mécanismes d’action des cellules du système inné
Phagocytose
Les cellules les plus impliquées dans la phagocytose sont les macrophages, les monocytes, les polynucléaires neutrophiles et les cellules dendritiques : ce sont des « phagocytes professionnels ». La phagocytose s'adresse à des particules de grande taille, supérieures à 0,5 micron. La phagocytose dépend de l'actine.
Le mot « phagocytose » signifie littéralement « ingestion de cellule ». Les phagocytes sont des cellules pouvant engouffrer (phagocyter) diverses particules, en particulier des agents infectieux. La phagocytose s'effectue en plusieurs étapes. Tout d'abord, le phagocyte allonge des parties de sa membrane plasmique, entourant ainsi progressivement la particule jusqu'à ce que cette dernière soit à l'intérieur de la cellule. À l'intérieur de la cellule, la particule phagocytée se trouve dans un endosome qui fusionne ensuite avec le lysosome pour former le phagosome[6]. Le lysosome contient des enzymes et des acides permettant de tuer et de digérer la particule.
Les phagocytes permettent également l'élimination des cellules mortes de l'hôte, que celles-ci aient connu une mort programmée (apoptose) ou une mort cellulaire à la suite d'une infection bactérienne ou virale[2]. La phagocytose constitue ainsi un phénomène important dans le processus de cicatrisation.
| Molécules détruisant le pathogène | |
|---|---|
| Agents anti microbiens | |
| Protéases | Lysozyme, cathepsine, élastase |
| Peptides antimicrobiens | |
| Lactoferrine | Capture du fer (indispensable à la croissance des pathogènes) |
| Réactifs oxygénés ROS (Reactif oxygen species) | |
| NADPH oxydase | |
| NO synthétase | |
Reconnaissance directe
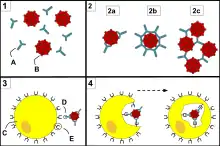
Le phagocyte reconnait le pathogène directement par ses récepteurs de reconnaissance de motifs moléculaires.
Reconnaissance indirecte ou opsonisation
Le phagocyte reconnait de façon indirecte le pathogène par une opsonine, molécule permettant le déclenchement de la phagocytose. Deux molécules jouent ce rôle :
- les immunoglobulines : les immunoglobulines se fixent sur le pathogène. Le phagocyte reconnait l'immunoglobuline par des récepteurs Fc reconnaissant la partie constante de l'immunoglobuline ;
- la fraction C3b du complément ; le phagocyte reconnait C3b par des récepteurs spécifiques.
Fonctions de la phagocytose
La phagocytose a deux fonctions :
- elle détruit le pathogène ;
- elle commence la présentation de l’antigène par la cellule dendritique au lymphocyte T CD4+. Cette présentation de l’antigène est la première étape de la réponse adaptative de l'organisme à l'infection.
Dégranulation
La dégranulation est un mécanisme de défense pour les pathogènes trop gros pour être phagocytés. Ce mécanisme est la libération de substance toxique pour le pathogène à l'extérieur de la cellule, mais avec un risque de dommage sur les cellules saines. Les neutrophiles libèrent beaucoup de molécules lors d'un contact avec un agent (lactoferrine, protéases, lysozymes, BPI). Les défenses antiparasitaires se fondent sur la production de molécules par les éosinophiles principalement (eosinophil cationic peptide ECP, eosinophil derived neurotoxin EDN, peroxydases). Dans certains cas, la dégranulation s'accompagne de la mort de la cellule immunitaire par expulsion de composants cellulaires. Cette action, dont le but n'est pas bien connu, pourrait permettre la formation de filets extra-cellulaires (neutrophil extracellulaur traps NET).
Production de cytokines et de substances vasodilatatrices
La production de la cellule dendritique n'est pas représentée ici car sa fonction principale est l'initiation de la réponse adaptative en se déplaçant du lieu de l'infection vers le ganglion lymphatique pour présenter l'antigène à la cellule T CD4+ auxiliaire.
- Par le macrophage :
- Par le mastocyte : histamine, protèases, prostaglandines, héparine, cytokines, chimiokines, facteurs de croissance
Cytotoxicité
La cytotoxicité est le mode d'action des cellules NK. Elles sont constitutivement cytotoxiques : ce sont des tueuses professionnelles. Ce sont des lymphocytes dépourvus de récepteur. Elles participent à l'élimination des cellules cancéreuses et des cellules infectées par un germe intracellulaire. Elles reconnaissent les cellules qui ne sont pas porteuses de l'antigène du soi. Les NK reconnaissent les cellules à détruire par le complexe majeur d'histocompatibilité de type I (CHM ou MCH).
Mode d'action des cellules NK
Les NK tuent de deux façons :
- par contact direct et libération du contenu de leur granules (granzymes, perforine) qui créent des pores dans la membrane cellulaire avec fuite de matériel cellulaire et la mort des cellules[11] ;
- par simulation des récepteurs de mort présent sur les cellules (death receptor Fas /Fas ligand du NK). Ce mécanisme ne serait pas le mécanisme principal[12],[13],[14].
Reconnaissance des cellules à tuer par la cellule NK
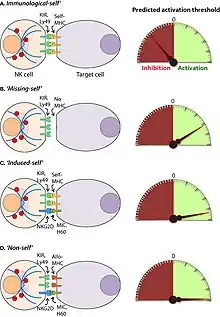
La reconnaissance des cellules à tuer par la cellule se fait par deux mécanismes :
- par activation des récepteurs des NK (NKR) la cellule NK est naturellement tueuse. Les récepteurs activateurs de la cellule NK sont en équilibre avec des récepteurs inhibiteurs de la cellule NK. Ils ne produisent aucun effet tant qu'une cellule présente des signes d’absence d'anomalies. La non activation de la cellule NK résulte d'un équilibre entre des récepteurs inhibiteurs et des récepteurs activateurs présents dans une cellule NK. L'activation est une rupture de l'équilibre naturel ;
- par activation des récepteurs Fc a faible affinité pour les immunoglobulines G.
Activation des récepteurs des NK
Dans un état de non activation, la cellule NK reconnait un ligand de son récepteur activateur sur une cellule saine et la cellule saine présente des peptides appartenant au soi sur son complexe majeur d'histocompatibilité de classe I au récepteur inhibiteur. La rupture de l'équilibre se fait de deux façons.
- En cas d'infection virale, de nombreux virus pour empêcher l'action des cellules immunitaires empêchent l'expression des complexes majeurs d'histocompatibilité de classe I[15], il n'y a donc pas de présentation de peptides viraux. Cette absence de complexe majeur d'histocompatibilité de classe débloque l'inhibition naturelle.
- En cas d'infection virale, bactérienne ou par des mycobactéries (tuberculose, listéria) ou cas de cellule cancéreuse, la cellule produit des protéines cellules de stress (MIC1, MIC2). Ces protéines vont stimuler puissamment les récepteurs activateurs de la cellule NK, aboutissant à la dégranulation de celle-ci.
Activation des récepteurs Fc a faible affinité pour les immunoglobulines G
Une cellule infectée ou tumorale va exprimer des récepteurs cellulaires qui n’existent pas dans une cellule saine. La partie variable des immunoglobulines G se fixe sur ces récepteurs et déclenche l'action cytotoxique des cellules NK.
Molécules du système immunitaire inné
Peptides anti-microbiens
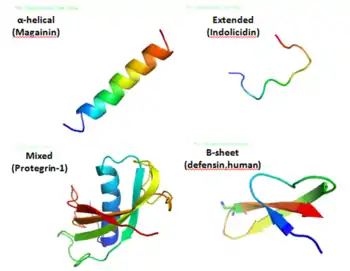
Les peptides anti-microbiens (PAM) sont des oligopeptides comprenant de 6 à 100 acides aminés. Il existe actuellement plus de 20 000 peptides anti-microbiens dans la base de données Data repository of antimicrobial peptides.
Ils constituent la première ligne de défense chimique et sont actifs contre les virus, les bactéries et les champignons[20]. La plupart des PAMs sont produits en permanence par les cellules mais quelques-uns sont produits lorsqu'il y a un agresseur spécifique comme le P9A et le P9B produits après l'injection d'Enterobacter cloaca chez un papillon[21] ou de défensine produite après l'infection de Pseudomonas aeruginosa chez la souris[22]. Les bactéries, au cours de leur destruction par les antibiotiques ou par le système immunitaire, libèrent des polysaccharides induisant une production de PAM[23].
À côté de leur rôle de stimulateur de la réaction immunitaire, ils ont aussi un rôle de modulateur de la réaction immunitaire en bloquant, par exemple, la synthèse de cytokine[24],[25],[26]. Cette régulation n’existe pas lors de l'utilisation d'antibiotique.
Protéines de la phase aiguë
Les protéines de la phase aiguë sont des protéines sécrétées par le foie circulant dans le sang de façon permanente et dont le taux varie lors de l'inflammation[27]. Les protéines de la phase aiguë comprennent la mannose binding protein (MBP), la C reactive-proteine (CRP), les facteurs du complément, les protéines de liaison au lipopolysaccharide, les inhibiteurs de réactifs oxygénés, le sérum amyloïde A, le sérum amyloïde P et l'haptoglobine.
L'augmentation de leur synthèse est déclenchée par les cytokines inflammatoires comme l'interleukine 1, l'interleukine 6, le facteur de nécrose tumorale, le facteur de croissance transformant et les interférons I produits par les macrophages et les monocytes[28],[29].
Les protéines de la phase aiguë ont différentes fonctions :
- dans la protéolyse pour les protéases,
- dans la coagulation du sang pour le fibrinogène, la prothrombine,
- dans le système du complément pour C1, C2, etc.,
- en tant que protéines de transport pour l'haptoglobine et la céruléoplasmine,
- d'autres fonctions (voir détails ci-dessous) pour la protéine C réactive, la sérum amyloïde A, l'orosomucoïde et le sérum amyloïde P.
En fait, il existe deux catégories de protéines de la phase aiguë ; celles dont le taux augmente sont nommées les protéines positives de la phase aiguë, celles dont le taux baisse sont nommées les protéines négatives de la phase aiguë[30]. En pratique clinique humaine et vétérinaire, les plus intéressantes sont les protéines positives de la phase aiguë[30].
Les trois protéines de la phase aiguë dont le taux varie le plus au cours d'une inflammation aiguë sont la protéine C réactive, le sérum amyloïde A et le sérum amyloïde P. La protéine C réactive et le sérum amyloïde A peuvent augmenter jusqu'à 100 fois leur concentration de base. La plupart des autres protéines de la phase aiguë augmentent de 2 à 10 fois leur concentration de base[30].
Protéine C réactive
La protéine C réactive est probablement une des protéines de la phase aiguë les plus utilisées en pratique médicale pour diagnostiquer une infection. La concentration sanguine normale est inférieure à 6 mg par litre (en fait, ce taux dépend des normes du laboratoire). En cas d'une infection bactérienne, son taux sanguin commence à s’élever et suit la gravité de l'infection. L'évolution du taux de protéine C réactive permet donc de suivre l'évolution de l'infection et de vérifier l'efficacité d'un traitement. Les infections bactériennes donnent des concentrations plus élevées que les infections virales. Mais toutes les infections ne donnent pas d'élévation de cette protéine, comme les infections urinaires localisées à la vessie ou les infections vaginales[30].
Une propriété majeure de la protéine C réactive est de se lier aux récepteurs de reconnaissance de motifs moléculaires calcium-dépendants permettant de fixer les pathogènes dont la membrane contient des phosphorylcholines entraînant l’activation du système du complément et le dépôt du facteur C3b du complément préparant l'opsonisation[31].
Sérum amyloïde A
La sérum amyloïde A est une apolipoprotéine jouant un rôle essentiel chez tous les mammifères. Elle est impliquée dans le recrutement cellulaire des cellules de l'immunité innée et dans la production d'enzymes. Elle est retrouvée dans le lait maternel en cas de mastite[32].
Sérum amyloïde P
Le composant amyloïde P de la pentraxine sérique ou sérum amyloïde P (SAP) est sécrété par le foie et se trouve dans le plasma à une concentration d'environ 30 mg/L. Son rôle est peu connu mais semble impliquer dans la protéolyse, l’hématose et l'activation du complément[33]. Elle est aussi impliquée dans l’opsonisation[34].
Haptoglobine
L'haptoglobine est principalement synthétisée par le foie. C'est une protéine plasmatique qui se lie avec l'hémoglobine des érythrocytes lors de la destruction de ceux-ci.
Protéine de liaison de la mannose
La protéine de liaison de la mannose calcium-dépendante est la molécule la plus efficace pour l'opsonisation. Elle se lie non seulement à la mannose mais à beaucoup d'autres sucres présents dans les parois, permettant de se lier à un très grand nombre de bactéries Gram positif, bactéries Gram négatif, virus, levures ou parasites déclenchant la voie classique du système du complément[35].
Cette molécule appartient à la famille des collectines, qui ont la propriété de reconnaître les sucres des parois des organismes étrangers qui n'ont pas le terminal galactose acide sialique[35].
Inhibiteur de réactif oxygéné
Le réactif oxygéné apparaît pendant la phagocytose des membranes de l'endosome des cellules phagocytaires avec la participation de la nicotinamide adénine dinucléotide phosphate oxydase des membranes cellulaires[36],[37].
Inhibiteurs de protéases
Dix pour cent des composants du plasma sont des inhibiteurs de protéases comme l'Alpha-2 macroglobuline.
Système du complément
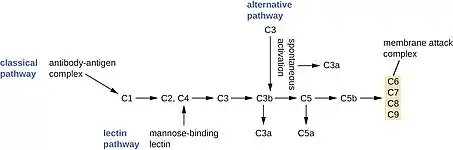
Le système du complément est le nom donné en 1930 pour définir une trentaine de protéines qui « complètent » (d'où le nom de complément) la capacité des anticorps à éliminer les pathogènes : il fait donc d'abord partie du système adaptatif. Les composants de la cascade du complément sont présents dans de nombreuses espèces en plus des mammifères, comme les plantes, les oiseaux, les poissons et quelques invertébrés[10]. Le système du complément est aussi une partie intégrante du système inné.
L'activation du complément se fait par une cascade protéolytique. La cascade du complément implique de nombreuses protéines du plasma, synthétisées dans le foie, principalement par les hépatocytes.
Il existe trois voies d'activation du complément : la voie classique (la première découverte) déclenchée par les immunoglobulines G ou M, la voie des lectines déclenchée par la protéine de liaison de la mannose, la voie alternative qui est déclenchée par l'absence de molécules du soi. Ces trois voies vont activer des serines à protéases mais ayant des activités similaires pour aboutir à :
- la formation d'une enzyme C3 convertase qui est capable de convertir la protéine C3 du complément en protéine C3a et C3b. La protéine C3a est une chemokine qui active l'inflammation. La protéine C3b favorise l'opsonisation ;
- la cascade continue pour aboutir à la formation de la protéine C5a et de la protéine C5b par action d'une C5 convertase sur la protéine C5 du complément. La protéine C5a est une chemokine qui active l'inflammation ; les protéines C3a et C5a du complément sont appelées anaphylatoxines en immunologie ;
- la protéine C5b va activer d'autres protéines du complément pour aboutir à la formation, à la surface d'un pathogène, d'un complexe protéique aboutissant à la formation de pores dans les membranes des pathogènes, le complexe d’attaque membranaire (Membrane Attack Complex ou MAC).
Les protéines du complément agissent de concert pour :
- induire le recrutement de cellules immunitaires ;
- recouvrir les pathogènes et permettre ainsi leur élimination par opsonisation ;
- recouvrir les pathogènes et induire directement des perforations conduisant à la mort directe de l'organisme pathogène ;
- éliminer les complexes immuns.
Voie classique
Un anticorps G ou M se fixe sur une bactérie. Le complément C1 clive les facteurs C4 et C2 en se fixant sur l'immunoglobuline. Les fragments C4a et C2a se fixent à la surface du microbe et forment la C3 convertase qui clive à son tour le facteur C3 en C3a et C3b :
- Le C3a diffuse et active l'inflammation ;
- Le C3b se fixe sur le microbe formant avec C4b et C2b la C5 convertase.
Voie des lectines
Cette voie est déclenchée par la fixation d'une lectine, la mannose binding lectine, sur des carbohydrates présents sur la paroi des microbes. La cascade du complément est la même à l’exception de la première étape impliquant C1.
Voie alternative
Elle met en jeu des systèmes de reconnaissance du non soi. Le facteur C3 s’hydrolyse spontanément à la surface des cellules de l'organisme ou d'un microbe. L'hydrolysation aboutit à C3a et C3b. À la surface des cellules de l'organisme, il existe des protéases qui vont dégrader le C3b, par contre sur les microbes il n'existe pas cette protéase.
L’étape finale
L'étape finale aboutit à stimuler l'inflammation par le C5a et la lyse cellulaire par formation du complexe d'attaque cellulaire (membrane attack complex, MAC). La C5 convertase clive la C5 en C5a et C5b. La C5b forme avec C6, C7 et C8 un complexe entraînant la polymérisation de C9 formant le complexe d'attaque cellulaire, un pore qui vide le pathogène de son contenu.
Interférons de type I
Attention les interférons de type I ont des propriétés différentes de l'interféron de type II aussi nommé interféron gamma.
Les interférons I alpha et I beta sont des cytokines produites lors d'une infection virale[38]. Un ARN double brin d'origine virale se fixe sur un récepteur de pattern TLR3 présent dans un phagosome intra cellulaire. Cette fixation entraîne l'activation des facteurs de transcription NFK et IRF3 déclenchant la synthèse de l'interféron alpha et béta. Les interférons alpha et beta sont de type I. Ils génèrent un état antiviral par plusieurs mécanismes :
- résistance à la réplication (multiplication) virale intracellulaire ;
- augmentation de l'expression des MHC de classe I ;
- accroissement de la présentation de l'antigène ;
- activation des cellules NK pour éliminer les cellules infectées par le virus.
Inflammation
L'inflammation est l'une des premières réponses du système immunitaire contre une infection ou une irritation. L'inflammation est induite par des facteurs chimiques relargués par les cellules abîmées et permet d'établir une barrière efficace contre la dissémination d'éventuels agents infectieux, et d'initier les processus de réparation tissulaire après l'élimination des agents pathogènes[7].
L'inflammation est induite par les cellules initialement présentes dans la plupart des tissus, comme les macrophages, les cellules dendritiques, les histiocytes, les cellules de Kupffer ou les mastocytes. Ces cellules expriment à leur surface des récepteurs de reconnaissance de motifs moléculaires. Ces récepteurs reconnaissent des molécules, appelées motifs moléculaires associés aux pathogènes, qui sont exprimées par les organismes microbiens, mais distinctes des molécules de l'organisme. Au début d'une infection, d'une brûlure, ou d'une altération des tissus, la reconnaissance de motifs moléculaires associés aux pathogènes par les récepteurs de reconnaissance de motifs moléculaires exprimés par ces cellules conduit à leur activation et à la sécrétion de différents médiateurs responsables des signes cliniques de l'inflammation (douleur, rougeur, chaleur et gonflement).
Les facteurs chimiques produits durant l'inflammation (histamine, bradykinine, sérotonine, leucotrienes et prostaglandines) augmentent la sensation de douleur, induisent localement la vasodilatation des vaisseaux sanguins et le recrutement de phagocytes, en particulier les neutrophiles[7]. Les neutrophiles peuvent également produire des facteurs solubles contribuant à la mobilisation d'autres populations de leucocytes. Les cytokines produites par les macrophages et les autres cellules du système immunitaire inné constituent un relais de la réponse immunitaire. On compte, parmi ces cytokines, le TNFα, HMGB1, et l'interleukine-1[39].
Inflammation locale
Première étape ; le franchissement de la barrière épithéliale
Que ce soit la barrière épithéliale pluristratifiée de la peau ou la barrière épithéliale faite d'une seule couche de cellules comme dans la muqueuse respiratoire, la barrière épithéliale est la première en contact avec le pathogène. Elle libère dans les tissus sous-jacents des peptides antimicrobiens et cytokines pro-inflammatoires (interleukine-1, interleukine-6, facteur de nécrose tumorale)
Deuxième étape ; la stimulation des cellules résidentes
Ces cytokines agissent sur les macrophages et les mastocytes qui libèrent des cytokines et des molécules vaso-actives.
Troisième étape ; le recrutement des cellules immunitaires sanguines
Les mastocytes libèrent des molécules vaso-actives (histamine, substance P, leucotriènes, prostaglandines) entraînant :
- une dilatation des artérioles ;
- une perméabilité accrue des capillaires permettant aux cellules du complément et des anticorps de traverser l'endothélium.
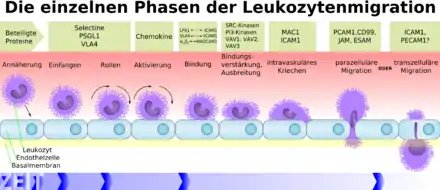
Les macrophages et les mastocytes qui libèrent des cytokines et des chimiokines pro-inflammatoire :
- augmentent les molécules d'adhésion sur les cellules endothéliales des capillaires ;
- induisent le recrutement des cellules circulantes (neutrophiles, monocytes).
La transmigration comprend 4 phases bien décrites
- Roulement (en anglais rolling) : adhérence faible entre une sélectine E capillaire et un carbohydrate de la cellule immunitaire
- Activation (activation) par interaction entre une chimiokine capillaire et une intégrine cellulaire
- Adhérence ferme (adhesion) entre l'intégrine cellulaire (LFA-1) et son ligand vasculaire (ICAM-1). La cellule est fixée sur l'endothélium
- La traversée de l'endothélium par les leucocytes ou diapédèse intervient dans un segment particulier du système circulatoire : les veinules post-capillaires.
Rôle critique des cytokines
Les trois cytokines majeures de l'inflammation sont l'interleukine-1, l'interleukine-6 et le facteur de nécrose tumorale[40],[41]. On les nomme le trio pro-inflammatoire.
| Effet local | Effet systémique |
|---|---|
| Augmentation de l’adhérence cellulaire | Fièvre |
| Augmentation de la libération du NO | Action au niveau de l'axe hypothalamo-hypophysaire avec augmentation de la libération de cortisol |
| Augmentation de la libération de l'interleukine 1, 6, 8 et du TNF | Stimulation de l'hématopoïèse (neutrophile) |
| Augmentation de la production des anticorps par les lymphocytes B | Augmentation de la libération des protéines de la phase aiguë |
Inflammation systémique
L 'Inflammation systémique implique trois organes (le foie, le système nerveux central et les glandes surrénales) et le trio pro-inflammatoire (l'interleukine-1, l'interleukine-6 et le facteur de nécrose tumorale).
Les molécules inflammatoires sensibilisent les terminaisons nerveuses. Les neurones relarguent la substance P et la CGRP des peptides qui ont des actions vaso-dilatatrices puissantes.
Résolution de l'inflammation
L'inflammation peut :
- aboutir à la guérison de l'individu ;
- donner une cicatrice ;
- être limitée dans une partie du corps et persister sous forme de granulome comme le granulome pulmonaire de la tuberculose ;
- se diffuser dans tout l'organisme sous forme de septicémie aboutissant à un choc toxique souvent mortel.
Système de reconnaissance
Les cellules effectrices du système immunitaire inné reconnaissent les agents pathogènes extérieurs par des récepteurs[3] appelés pattern recognition receptors (PRR) ou, en français, récepteurs de reconnaissance de motifs moléculaires, appelés encore récepteurs de pattern. Ces cellules effectrices peuvent reconnaître les dommages causés dans les cellules par les agents pathogènes : ce sont les signaux de danger. Le troisième mécanisme est le mécanisme de reconnaissance du soi et du non soi.
Système de reconnaissance des pathogènes
La reconnaissance d'un pathogène par le système immunitaire inné repose sur deux outils : le motif moléculaire associé aux pathogène et les récepteurs de reconnaissance de motifs moléculaires.
Motifs moléculaires associés aux pathogènes
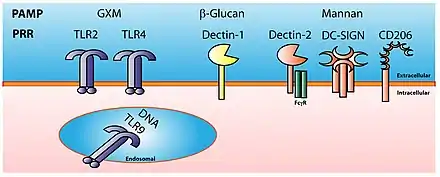
Ce sont des structures moléculaires associées aux pathogènes et qui n'existent pas chez les mammifères. Ces structures moléculaires sont indispensables aux pathogènes et sont donc peu susceptibles de mutation. Ces structures ne sont pas spécifiques d'une espèce mais d'une famille de microbes ; ces motifs moléculaires sont portés par toutes les familles de microbes, qu'ils soient ou non pathogènes pour l'organisme. Au lieu d'utiliser le terme de motifs moléculaires associés aux pathogènes (PAMP), il faudrait utiliser le terme de motifs moléculaires associés aux microbes (MAMP).
Les procaryotes sont plus faciles à distinguer que les eucaryotes (comme les parasites) par les récepteurs de reconnaissance de motifs moléculaires car un parasite est plus proche phylogénétiquement d'un être humain que d'une bactérie. L'infection parasitaire (paludisme, toxoplasmose) est donc plus difficile à combattre pour l'organisme que l'infection virale ou bactérienne. Cela explique aussi la difficulté de la mise au point de vaccin contre les parasites.
| Famille de microbes | Motifs moléculaires |
|---|---|
| Virus | ARN double brin, ADN avec motif cytosine-guanine non méthylé |
| Bactérie gram-positive | Acide téchoïque, acide lipotéchoïque, lipoprotéine, peptidoglycane |
| Bactérie gram-négative | Lipopolysaccharide, lipoprotéine, peptidoglycane |
| Mycobactérie | |
| Champignon | Glucane, manname, GKM |
Récepteurs de reconnaissance de motifs moléculaires
Les récepteurs de reconnaissance de motifs moléculaires sont codés par les cellules de la lignée germinale et sont spécifiques d'un motif moléculaire.
Récepteurs de reconnaissance de motifs moléculaires par type
| Nom du récepteur | Localisation cellulaire | Motifs moléculaires reconnus | Famille de microbes | Nom |
|---|---|---|---|---|
| Récepteurs de type Toll (TLR) | Membranaire, phagosome | Lipoprotéine, Lipopolysaccharide, ARN simple et double brin, ADN bactérien | Virus, bactérie | TLR1 à TLR11 |
| Récepteurs de type Nod (NOD) | Cytoplasme | Peptidoglycane | Bactérie | NOD1, NOD2 |
| Récepteurs hélicases (RLR) | Cytoplasme | ARN | RIG-1, MDA5 | |
| Récepteur cGAS | Cytoplasme | ADN double brin | Virus |
Système de reconnaissance des cellules lésées
L'idée que le système immunitaire puisse reconnaître les cellules lésées par un pathogène au lieu du pathogène lui-même n'est pas une théorie admise par tous les scientifiques: protéine de stress, protéine de choc thermique comme le HSP ou l'actine. L'actine est un composé intracellulaire, la présence d'actine extra-cellulaire est une indication de cellule lésée.
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Innate immune system » (voir la liste des auteurs).
- (en) Paul Grasso, Essentials of Pathology for Toxicologists, CRC Press (lire en ligne)
- (en) Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters, Molecular Biology of the Cell; Fourth Edition, New York and London, Garland Science, (ISBN 0-8153-3218-1, lire en ligne)
- (en) Hajime Kono et Kenneth L. Rock, « How dying cells alert the immune system to danger », Nature Reviews Immunology, vol. 8, no 4, , p. 279–289 (ISSN 1474-1741, PMID 18340345, PMCID PMC2763408, DOI 10.1038/nri2215, lire en ligne, consulté le )
- IMMUNOLOGY - CHAPTER ONE > INNATE (NON-SPECIFIC) IMMUNITY Gene Mayer, Ph.D. Immunology Section of Microbiology and Immunology On-line. University of South Carolina
- (en) J. K. C. Chan, C. S. Ng et P. K. Hui, « A simple guide to the terminology and application of leucocyte monoclonal antibodies », Histopathology, vol. 12, no 5, , p. 461–480 (ISSN 1365-2559, DOI 10.1111/j.1365-2559.1988.tb01967.x, lire en ligne, consulté le )
- (en) Charles Janeway, Paul Travers, Mark Walport, and Mark Shlomchik, Immunobiology, New York et London, Garland Science, , 5e éd. (ISBN 0-8153-4101-6, lire en ligne).
- (en) Viera Stvrtinová, Ján Jakubovský et Ivan Hulín, Inflammation and Fever from Pathophysiology: Principles of Disease, Computing Centre, Slovak Academy of Sciences, Academic Electronic Press, (lire en ligne)
- Rosenau W, Moon HD. Lysis of homologous cells by sensitized lymphocytes in tissue culture. J Natl Cancer Inst (1961) 27:471–83.
- Smith HJ. Antigenicity of carcinogen-induced and spontaneous tumours in inbred mice. Br J Cancer (1966) 20:831–7. doi:10.1038/bjc.1966.95
- (en) Janeway CA, Jr. et al., Immunobiology., Garland Science, , 6e éd. (ISBN 0-443-07310-4)
- Bhat R, Watzl C. Serial killing of tumor cells by human natural killer cells – enhancement by therapeutic antibodies. PLoS One (2007) 2:e326. doi:10.1371/journal.pone.0000326
- Smyth MJ, Cretney E, Kelly JM, Westwood JA, Street SE, Yagita H, et al. Activation of NK cell cytotoxicity. Mol Immunol (2005) 42:501–10. doi:10.1016/j.molimm.2004.07.034
- (en) Maria Eugenia Guicciardi et Gregory J. Gores, « Life and death by death receptors », The FASEB Journal, vol. 23, no 6, , p. 1625–1637 (ISSN 0892-6638 et 1530-6860, PMID 19141537, PMCID PMC2698650, DOI 10.1096/fj.08-111005, lire en ligne, consulté le )
- A. Ashkenazi, « Death Receptors: Signaling and Modulation », Science, vol. 281, no 5381, , p. 1305–1308 (DOI 10.1126/science.281.5381.1305, lire en ligne, consulté le )
- (en) Antony N. Antoniou et Simon J. Powis, « Pathogen evasion strategies for the major histocompatibility complex class I assembly pathway », Immunology, vol. 124, no 1, , p. 1–12 (ISSN 0019-2805 et 1365-2567, PMID 18284468, PMCID PMC2434379, DOI 10.1111/j.1365-2567.2008.02804.x, lire en ligne, consulté le )
- Norberto Walter Zwirner et Andrea Ziblat, « Regulation of NK Cell Activation and Effector Functions by the IL-12 Family of Cytokines: The Case of IL-27 », Frontiers in Immunology, vol. 8, (ISSN 1664-3224, PMID 28154569, PMCID PMC5243847, DOI 10.3389/fimmu.2017.00025, lire en ligne, consulté le )
- (en) Laurent Chiche, Jean-Marie Forel, Guillemette Thomas et Catherine Farnarier, « The Role of Natural Killer Cells in Sepsis », Journal of Biomedicine and Biotechnology, vol. 2011, , p. 1–8 (ISSN 1110-7243 et 1110-7251, PMID 21629707, PMCID PMC3100670, DOI 10.1155/2011/986491, lire en ligne, consulté le )
- (en) Markus Bosmann et Peter A. Ward, « Modulation of inflammation by interleukin-27 », Journal of Leukocyte Biology, vol. 94, no 6, , p. 1159–1165 (PMID 23904441, PMCID PMC3828601, DOI 10.1189/jlb.0213107, lire en ligne, consulté le )
- Annie Tosello-Trampont, Fionna A. Surette, Sarah E. Ewald et Young S. Hahn, « Immunoregulatory Role of NK Cells in Tissue Inflammation and Regeneration », Frontiers in Immunology, vol. 8, (ISSN 1664-3224, PMID 28373874, PMCID PMC5357635, DOI 10.3389/fimmu.2017.00301, lire en ligne, consulté le )
- (en) Katherine Radek et Richard Gallo, « Antimicrobial peptides: natural effectors of the innate immune system », Seminars in Immunopathology, vol. 29, no 1, , p. 27–43 (ISSN 1863-2297 et 1863-2300, DOI 10.1007/s00281-007-0064-5, lire en ligne, consulté le )
- (en) Dan Hultmark, Håkan Steiner, Torgny Rasmuson et Hans G. Boman, « Insect Immunity. Purification and Properties of Three Inducible Bactericidal Proteins from Hemolymph of Immunized Pupae of Hyalophora cecropia », European Journal of Biochemistry, vol. 106, no 1, , p. 7–16 (DOI 10.1111/j.1432-1033.1980.tb05991.x, lire en ligne, consulté le )
- (en) Bals R et Wang X, « Mouse Beta-Defensin 3 Is an Inducible Antimicrobial Peptide Expressed in the Epithelia of Multiple Organs », sur Infection and immunity, 1999 jul (PMID 10377137, consulté le )
- (en) R. E. W. Hancock et M. G. Scott, « The role of antimicrobial peptides in animal defenses », Proceedings of the National Academy of Sciences, vol. 97, no 16, , p. 8856–8861 (ISSN 0027-8424 et 1091-6490, PMID 10922046, PMCID PMC34023, DOI 10.1073/pnas.97.16.8856, lire en ligne, consulté le )
- (en) Thomas Birchler, Reinhart Seibl, Katja Büchner et Susanne Loeliger, « Human Toll-like receptor 2 mediates induction of the antimicrobial peptide human beta-defensin 2 in response to bacterial lipoprotein », European Journal of Immunology, vol. 31, no 11, , p. 3131–3137 (ISSN 1521-4141, DOI 10.1002/1521-4141(200111)31:113.0.CO;2-G, lire en ligne, consulté le )
- Brackett, D J et al. “A synthetic lipopolysaccharide-binding peptide based on the neutrophil-derived protein CAP37 prevents endotoxin-induced responses in conscious rats.” Infection and immunity vol. 65,7 (1997): 2803-11.
- Larrick JW, Hirata M, Balint RF, Lee J, Zhong J, Wright SC. Human CAP18: a novel antimicrobial lipopolysaccharide-binding protein. Infect Immun. 1995;63(4):1291–1297.
- (en) Michael P. Murtaugh, « Porcine cytokines », Veterinary Immunology and Immunopathology, vol. 43, nos 1-3, , p. 37–44 (DOI 10.1016/0165-2427(94)90118-X, lire en ligne, consulté le )
- (en) Bruce Beutler et Anthony Cerami, « Cachectin/tumor necrosis factor: An endogenous mediator of shock and inflammation », Immunologic Research, vol. 5, no 4, , p. 281–293 (ISSN 0257-277X et 1559-0755, DOI 10.1007/BF02935501, lire en ligne, consulté le )
- Vegad JL. A textbook of veterinary General Pathology. 2nd ed. Lucknow: Publ. International Book distributing Co; 2007. Inflammation; pp. 105–82
- Sachin Jain, Vidhi Gautam et Sania Naseem, « Acute-phase proteins: As diagnostic tool », Journal of Pharmacy and Bioallied Sciences, vol. 3, no 1, , p. 118–127 (ISSN 0976-4879, PMID 21430962, PMCID 3053509, DOI 10.4103/0975-7406.76489, lire en ligne, consulté le )
- (en) Livija Deban, Sebastien Jaillon, Cecilia Garlanda et Barbara Bottazzi, « Pentraxins in innate immunity: lessons from PTX3 », Cell and Tissue Research, vol. 343, no 1, , p. 237–249 (ISSN 0302-766X et 1432-0878, DOI 10.1007/s00441-010-1018-0, lire en ligne, consulté le )
- (en) P. D. Eckersall, F. J. Young, C. McComb et C. J. Hogarth, « Acute phase proteins in serum and milk from dairy cows with clinical mastitis », Veterinary Record, vol. 148, no 2, , p. 35–41 (ISSN 0042-4900 et 2042-7670, DOI 10.1136/vr.148.2.35, lire en ligne, consulté le )
- (en) Ebbe Toftgaard Poulsen, Kata Wolff Pedersen, Anna Maria Marzeda et Jan J. Enghild, « Serum Amyloid P Component (SAP) Interactome in Human Plasma Containing Physiological Calcium Levels », Biochemistry, vol. 56, no 6, , p. 896–902 (ISSN 0006-2960 et 1520-4995, DOI 10.1021/acs.biochem.6b01027, lire en ligne, consulté le )
- (en) Carolyn Mold, Rebecca Baca et Terry W. Du Clos, « Serum Amyloid P Component and C-Reactive Protein Opsonize Apoptotic Cells for Phagocytosis through Fcγ Receptors », Journal of Autoimmunity, vol. 19, no 3, , p. 147–154 (DOI 10.1006/jaut.2002.0615, lire en ligne, consulté le )
- IM Roitt et PJ Delves, Innate Immunity : Acute phase proteins increase in response to infection, Oxford, Blackwell, , 10e éd. (p.17).
- Forman HJ and Torres M: Reactive oxygen species and cell signaling: respiratory burst in macrophage signaling. Am J Respir Crit Care Med 166: S4-8, 2002.
- Minakami R and Sumimotoa H: Phagocytosis-coupled activation of the superoxide-producing phagocyte oxidase, a member of the NADPH oxidase (NOX) family. Int J Hematol 84: 193-198, 2006
- Le Page C, Génin P, Baines MG, Hiscott J. Interferon activation and innate immunity. Rev Immunogenet. 2000;2(3):374–386.
- (en) MT Lotze et KJ Tracey, « High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal », Nature reviews. Immunology, vol. 5, no 4, , p. 331–42 (PMID 15803152, DOI 10.1038/nri1594)
- (en) Oliver Dienz et Mercedes Rincon, « The effects of IL-6 on CD4 T cell responses », Clinical Immunology, vol. 130, no 1, , p. 27–33 (PMID 18845487, PMCID PMC2660866, DOI 10.1016/j.clim.2008.08.018, lire en ligne, consulté le )
- (en) Lauro Velazquez-Salinas, Antonio Verdugo-Rodriguez, Luis L. Rodriguez et Manuel V. Borca, « The Role of Interleukin 6 During Viral Infections », Frontiers in Microbiology, vol. 10, (ISSN 1664-302X, PMID 31134045, PMCID PMC6524401, DOI 10.3389/fmicb.2019.01057, lire en ligne, consulté le )
Voir aussi
Articles connexes
- Immunité passive
- Système immunitaire
- Système immunitaire adaptatif
- Récepteur de reconnaissance de motifs moléculaires.
- Motif moléculaire associé aux pathogènes
- Chimiokines
- Peptide antimicrobien
- Protéine de phase aiguë
- système du complément
- Motif moléculaire associé aux dégâts
- Complexe d'attaque membranaire
 Portail de la biologie
Portail de la biologie  Portail de la médecine
Portail de la médecine