Syndrome de domestication
Le syndrome de domestication fait référence à un ensemble de traits morphologiques, physiologiques et comportementaux héréditaires présents dans un ensemble d'espèces domestiquées mais absents chez leurs variantes sauvages respectives.
Ce syndrome, pouvant atteindre plantes et animaux, présente une nature phénotypique variable, mais reste considérablement apparent si on considère que les espèces partageant cet ensemble de caractères sont relativement éloignées phylogénétiquement.
Histoire
Les remarques de Darwin (1868)
Dans son ouvrage The Variation of Animals and Plants under Domestication [1], publié le , Charles Darwin met en évidence la variabilité observable chez les différentes espèces domestiquées.
Il remarque sans doute que la sélection humaine sur des traits qu'il considère avantageux pourrait expliquer cette diversité, mais en partie seulement. Il attribue majoritairement cette caractéristique à des phénomènes environnementaux; le fait d'avoir des régimes alimentaires particuliers aurait un effet sur le phénotype des organismes considérés. D'autre part, il affirme qu'une grande partie de cette variation phénotypique peut s'expliquer par une “sélection inconsciente” pendant des milliers d'années dans les débuts de la domestication.
Néanmoins, il est plus remarquable de noter que Darwin décrit aussi, parmi cette grande variabilité de formes, une série de caractères analogues pas observables chez les ancêtres sauvages des espèces étudiées.
L'expérience de Belyaev (1959 à de nos jours)
L’œuvre de Darwin inspira de nombreux scientifiques: Un siècle plus tard (1959), le généticien Dmitri Belyaev commence une célèbre expérience d'élevage sélectif de renard roux à fourrure argentée (Vulpes vulpes), dont le but est de comprendre quelles sont les premières étapes de la domestication qui mènent aux changements physiologiques, morphologiques et comportementaux décrits par ce syndrome [2].
Contrairement à Darwin, Belyaev considérait que les patrons de changement observables chez ces organismes étaient le fruit de changements génétiques forgés par sélection [2]. Son hypothèse se basait sur le fait que les individus étaient prioritairement sélectionnés par rapport à leur degré de docilité, et non pas par rapport à leur taille ou autres caractéristiques [2]. Des changements dans les voies génétiques régulant le comportement, issus d'une sélection humaine, auraient eu un impact très considérable sur le développement d'autres structures [2], donnant naissance aux traits caractéristiques du syndrome de domestication.
Ce phénomène sera par la suite décrit chez d'autres organismes, notamment les plantes utilisés en agriculture [3].
Le mécanisme d'acquisition de ces caractères reste encore mal compris, mais il est connu que des modifications dans les patrons développementaux et dans l'expression des gènes sont à la base de ce fait remarquable [4]. D'autre part, il a été démontré que l'acquisition de ces caractères chez les animaux s'est faite non pas séquentiellement où chaque trait apparait de manière indépendante, mais semi-conjointement [5],[6],[7].
Traits phénotypiques
Chez les espèces domestiques, de nombreuses études suggèrent de nombreux traits phénotypiques communs.

On peut distinguer trois catégories principales chez les animaux. Ces traits incluent :
- Caractères morphologiques: fourrure à couleurs variables, oreilles pendantes, queue en tire-bouchon, réduction de la taille du cerveau et réduction de la taille des dents. Ces caractères sont pédomorphiques.
- Caractères physiologiques : œstrus plus fréquents et non saisoniers, changements de concentration dans de nombreux neurotransmetteurs et altérations dans les niveaux d'hormone corticotrope.
- Caractères comportementaux : Accroissement de la soumission et de la docilité, prolongement des comportements juvéniles et diminution de la peur envers l'homme.
Des traits communs ont tout de même été identifiés chez des plantes :
- Graines : perte de la dissémination, diminution de la phase de dormance, augmentation de la taille et du nombre[9].
- Fruits : Augmentation de la taille et du nombre[10].


Mécanismes biologiques
L'effet épigénétique
Aucun “gène de domestication” n'a encore été découvert. La complexité du processus suggère que plusieurs gènes sont responsables des changements phénotypiques [4].
L'épigénétique peut expliquer, dans certains cas, des variations phénotypiques prononcées dans des périodes de temps courtes, pouvant avoir un effet sur la physiologie et le comportement [11]. Des différences de méthylation de certains gènes dans le thalamus/hypothalamus pourraient expliquer la perte de la peur envers l'humain et une augmentation de la docilité chez les espèces domestiquées [12]. Ces caractéristiques ayant des fondements physiologiques, la concentration d'hormones libérées par les systèmes glandulaires serait régulée vers le bas [13].
Il a été démontré non seulement que la méthylation de l'ADN diffère entre variantes domestiques et sauvages, mais que ces méthylations sont héréditaires et transmises à la descendance, ce qui explique que des comportements convergents en lien avec une perte d'agressivité existent chez des espèces domestiquées différentes [13]. La domestication étant un phénomène où les individus étaient originairement sélectionnés par leurs aptitudes comportementales, la méthylation de certains gènes aurait persisté dans leur progéniture aboutissant à des phénotypes avantageux pour l'exploitation humaine.
Malgré tout, l'importance de l'épigénétique est contestée: Des études expérimentales menées avec de nombreuses espèces démontrent qu'une sélection individuelle de la soumission mène souvent à l'obtention du reste des traits morphologiques cités ci-dessus [14],[2]. Cela suggère une origine commune des caractères comportementaux, physiologiques et morphologiques non explicables par la seule méthylation de l'ADN.
L'hypothèse des cellules de la crète neurale
Une première hypothèse pour expliquer l'expression simultanée de ces traits chez oiseaux et mammifères passe par un détournement des voies de migration des cellules de la crête neurale : ce processus étant contrôlé par de nombreux gènes, des variations d'expression de ces derniers semblent être à la base de ce phénomène [4].
Ces cellules se déplacent durant l'ontogenèse, donnant naissance aux précurseurs cellulaires de nombreux tissus [15],[16],[17],[18]; mélanocytes, l'os et le cartilage formant le crâne et les odontoblastes entre autres [19]. Compte tenu de ces informations, des changements de comportement dans la migration des cellules de la crête neurale auraient ainsi un effet direct ou indirect sur les structures dérivant de celles-ci, et donc un effet sur la couleur du pelage, le cerveau ou les dents, structures atteintes par le syndrome de domestication [4].
Les changements morphologiques chez les espèces domestiques sont donc le résultat de défauts dans la production des cellules de la crête neurale dans les premières étapes du développement. En effet, ces défauts seraient à l'origine d'une production insuffisante de mélanocytes, chondrocytes du squelette facial, odontoblastes et chondrocytes de l'oreille externe dérivés des cellules de la crête neurale, qui expliqueraient respectivement les variations de couleur, la réduction de la taille de la mâchoire et du museau, la réduction de la taille des dents et les oreilles pendantes [4].
Cette hypothèse illustre tout de même comment ces caractères apparaissent en même temps lors du processus de domestication et non pas séquentiellement et de façon indépendante comme les études de Belyaev et son équipe l'avaient démontré [5],[6],[7] ; elle témoigne aussi d'une origine commune des symptômes.
Quant aux différences physiologiques et comportementales, le même principe est applicable étant donné que les hormones impliquées (surtout l'épinéphrine) sont sécrétées par les glandes surrénales dérivées elles aussi des cellules de la crête neurale. Comme on l'a vu, les concentrations étant hypo-régulées et les glandes souvent réduites chez les variétés domestiques [20], une sous-production de cellules de la crête neural pourrait être l'explication basale du phénomène.
La diversité des traits phénotypiques

La domestication, qui voit ses origines dans le néolithique, reste un processus récent dans l'histoire de l'évolution. Les traits phénotypiques caractérisant les espèces domestiques étant relativement universels, la diversité de ces premiers reste importante tenant compte de l'actualité relative du processus.
D'un point de vue évolutionnaire, expliquer l'apparition d'une telle variabilité dans un intervalle de temps si restreint reste un défi. Néanmoins, des études suggèrent que les séquences répétées en tandem de certains gènes développementaux seraient source de diversité morphologique [21]. Des allèles présentant des contractions ou expansions hors norme pourraient expliquer les phénotypes extrêmes observables dans de nombreuses races et variétés, ces séquences incluant des familles de gènes développementaux telles que les gènes homéotiques, qui organisent le plan du corps d'une façon très générale.
Ces altérations de longueur dans les séquences répétées en tandem ont un effet notamment dans la différenciation des ostéoblastes, et peuvent provoquer des modifications morphologiques au niveau du crâne et des membres qui expliqueraient la diversité phénotypique des organismes domestiques observée par Darwin [21]. De plus, il a été démontré que les populations naturelles de mammifères placentaires présentent des altérations de longueur de séquences répétées au sein des gènes développementaux beaucoup plus modestes. De façon générale, leur degré de variation morphologique reste ainsi plus restreinte par rapport à leurs cousins domestiquées [21].
L'importance des facteurs de transcription

Chez plantes et animaux, les facteurs de transcription jouent un rôle indispensable pendant le développement et donc indirectement dans l'obtention des caractères relatifs au syndrome de domestication.
Chez les plantes, les gènes codant des facteurs de transcription MADS-box ont un rôle essentiel pendant le développement, c'est pourquoi ils ont souvent été comparés aux gènes Hox des animaux [23]. En effet, ils participent à la formation des phénotypes observables chez les variétés domestiques : Ils sont surexprimés pendant les premières phases du développement du fruit domestiqué [24] et pourraient en partie expliquer leur augmentation de taille relative par rapport aux variétés sauvages ; c'est un cas d'hétérométrie.
Des régulateurs négatifs de division cellulaire, tels que le gène Fruitweight2.2 (fw2.2) participent aussi à ce phénomène et sont responsables d'écarts de taille de 30 % [25].
D'autres gènes jouant un rôle important dans la formation des traits caractéristiques d'espèces domestiques ont été identifiés : shattering4 contrôle si la graine se détache de la plante pour participer à la dissémination des semences comme dans les variantes sauvages ou reste attachée à la plante comme dans les variantes domestiquées [26]. Ce gène codant un facteur de transcription, le changement d'un seul acide aminé dans le domaine de liaison à l'ADN est suffisant pour passer du caractère sauvage au caractère sélectionné [26], la récolte étant relativement plus simple si les graines restent attachés à la plante.
Syndrome de domestication : l'exemple du chien
La domestication est un phénomène anthropique qui touche une grande variété d’animaux appartenant à presque toutes les classes, soient les mammifères, les reptiles, les amphibiens, les oiseaux, les poissons et les insectes. Ces derniers ont été, au cours des 11 000 dernières années, utilisés par l’humain pour diverses raisons dont l’élevage destiné à la consommation, le travail, les produits dérivés et comme animaux de compagnie. Les animaux domestiqués occupent donc une grande place autant dans l’économie que dans la société et la religion[27]. Cette domestication a notamment permis de modeler les chiens que nous connaissons aujourd’hui par la transformation des loups sauvages tant au niveau comportemental qu’anatomique.
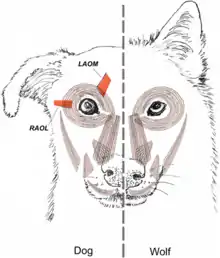
L’article « Evolution of facial muscle anatomy in dogs » par Kaminski, Waller, Diogo, Hartstone-Rose et Burrows publié en 2019[28], démontre d’ailleurs que l’anatomie faciale des chiens aurait été façonnée par la domestication principalement pour faciliter la communication avec les humains. La méthode utilisée pour la réalisation de l'étude a été séparée en deux catégories, soient en données anatomiques et en données comportementales. En ce qui concerne les données anatomiques, 4 loups sauvages (Canis lupus) et 6 chiens domestiques (Canis familiaris) ont été utilisés pour faire une dissection faciale comparative. Les résultats ont démontré que les principales distinctions anatomiques faciales entre les chiens domestiques et les loups sauvages se trouvent au niveau de la musculature entourant l'œil, soit la présence chez les chiens et l’absence chez les loups du levator anguli oculi medialis (LAOM), un muscle permettant de hausser le coin intérieur des sourcils indépendamment du plissement de relaxation des yeux. En effet, chez les loups, ce muscle n’est pas complètement formé et est plutôt simplement représenté par un tendon ou un amas de fibres situé au même endroit que le LAOM chez les chiens. L’autre différence est la présence et le niveau de développement du retractor anguli oculi lateralis (RAOL), un muscle qui permet de tirer latéralement le coin de la paupière vers les oreilles. Ce muscle, quoique présent chez plusieurs loups, est beaucoup plus mince et fragile que chez les chiens. D’autre part, bien qu’il soit présent chez tous les chiens domestiques, il est toutefois exceptionnellement absent chez le husky de Sibérie, une race de chien dont la domestication s’est faite plus tardivement et qui se rapproche donc davantage des loups. Au niveau des données comportementales, 9 loups et 27 chiens ont été individuellement filmés pendant 2 minutes chacun par un étranger se tenant face à eux à une distance d'environ 2 à 5 m afin de quantifier la fréquence et l'intensité d'un mouvement facial spécifique autour de l'œil permis par le LAOM, soit le relèvement intérieur du sourcil appelé AU101. Il a été démontré que ce mouvement est réalisé plus fréquemment et à une intensité plus élevée chez les chiens domestiques que chez les loups. Un mouvement similaire aurait pu être exprimé chez un ancêtre du chien domestique et aurait été, de manière consciente ou non, préféré et donc sélectionné au cours des 33 000 années séparant les chiens des loups et menant ainsi au développement de muscles distincts entre ces deux espèces. Une première hypothèse expliquant cette préférence est que l’exposition de la sclérotique blanche des yeux du chien par un mouvement AU101 exagéré est attrayant pour les humains parce qu’il aide à donner une direction au regard et que cela représenterait une adaptation pour la communication sociale et les interactions coopératives. Une seconde et plus probable hypothèse serait que le mouvement AU101 ressemble à l’expression faciale produite par les humains quand ils sont tristes et entraîne l’expression de caractéristiques pédomorphiques telles que des yeux plus gros donnant une apparence plus juvénile engendrant ainsi une réponse de soins et présentant un avantage sélectif. En effet, le regard mutuel et les caractéristiques énumérées ci-dessus entrainent une libération d’ocytocine qui occupe un rôle primordial dans l’affiliation, l’attachement et l’expression du comportement maternel. Les résultats obtenus confirment donc que la domestication aurait permis l’expression de mouvements et le développement de certains muscles chez les chiens domestiques en comparaison aux loups sauvages par des pressions de sélection visant à faciliter la communication.
Une étude réalisée par Waller et al. en 2013[29] traite justement de l’avantage sélectif dont bénéficient les chiens par leurs expressions faciales pédomorphiques. L’étude tentait de retracer la sélection humaine évolutive en déterminant si les humains choisissaient et adoptaient un chien en fonction des contractions musculaires faciales tels que des mouvements AU101 donnant une apparence plus juvénile par l’élévation du sourcil. Pour ce faire, les expérimentateurs ont désigné une race modèle, soit les « bulls » pour éviter toute variance de résultats. Ainsi, ils ont filmé 29 chiens appartenant aux races molossoïdes, Staffordshire Bull Terriers, Mastiffs et races mixtes de bull pendant 2 minutes chacun en face d’un expérimentateur. Ils ont ensuite mesuré le nombre et la fréquence de mouvements faciaux produits par les chiens à l’aide d’un système codant la gestuelle faciale (DogFACS) pour voir s’il y avait un lien entre le nombre de mouvements AU101 et l’adoption. Ils ont également observé si le fait d’agiter la queue avait un impact sur les chances d’être adopté. Les résultats ont démontré que les chiens qui produisaient davantage de mouvement AU101 étaient adoptés plus rapidement probablement parce que ce mouvement donne l’impression que les yeux sont plus volumineux. Ceci démontre donc que les chiens auraient utilisé ce mouvement comme avantage sélectif pour manipuler la préférence des humains envers les traits pédomorphiques. Chez les humains, surtout lorsque de la tristesse est ressentie, on retrouve l’équivalent du mouvement d’élévation du sourcil présent chez les chiens et ce dernier est nommée AU1. Ce mouvement favoriserait ainsi d’autant plus l’intention de s’investir autant chez les enfants humains que chez les chiens présentant ces caractères. À l’inverse, les résultats ont montré que l’agitation de la queue n’était pas un facteur favorisant l’adoption au même titre AU101, bien que l’action de remuer la queue soit un indicateur de bon tempérament. Cela témoigne donc de la force de sélection présente dans la domestication et engendrée par la préférence chez les humains pour le pédomorphisme au détriment de traits de tempérament réels. Cet article démontre donc que le mouvement AU101, entrainant l’expression de traits juvéniles chez les chiens domestiques, est apparu en réponse à la sélection indirecte humaine.
Voir aussi
Articles connexes
Références
- (en) Charles Darwin, The Variation of Animals and Plants Under Domestication, O. Judd, (lire en ligne)
- « Early Canid Domestication: The Farm-Fox Experiment » American Scientist », sur www.americanscientist.org (consulté le )
- (en) Geoffrey Peter Chapman, Grass Evolution and Domestication, Cambridge University Press, , 390 p. (ISBN 978-0-521-41654-2, lire en ligne)
- (en) Adam S. Wilkins, Richard W. Wrangham et W. Tecumseh Fitch, « The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and », Genetics, vol. 197, , p. 795-808 (ISSN 0016-6731 et 1943-2631, PMID 25024034, PMCID 4096361, DOI 10.1534/genetics.114.165423, lire en ligne, consulté le )
- (en) Belyaev, D. K, « Domestication of animals », Science Journal (U.K.), no 5, , p. 47-52
- (en) L. N. Trut, I. Z. Plyusnina et I. N. Oskina, « An Experiment on Fox Domestication and Debatable Issues of Evolution of the Dog », Russian Journal of Genetics, vol. 40, , p. 644-655 (ISSN 1022-7954 et 1608-3369, DOI 10.1023/B:RUGE.0000033312.92773.c1, lire en ligne, consulté le )
- (en) Lyudmila Trut, Irina Oskina et Anastasiya Kharlamova, « Animal evolution during domestication: the domesticated fox as a model », BioEssays, vol. 31, , p. 349-360 (ISSN 1521-1878, PMID 19260016, PMCID 2763232, DOI 10.1002/bies.200800070, lire en ligne, consulté le )
- (en) Adam S. Wilkins, Richard W. Wrangham et W. Tecumseh Fitch, « The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and », Genetics, vol. 197, , p. 795-808 (ISSN 0016-6731 et 1943-2631, PMID 25024034, PMCID 4096361, DOI 10.1534/genetics.114.165423, lire en ligne, consulté le )
- (en) Daniel Zohary, Maria Hopf et Ehud Weiss, Domestication of plants in the Old World : the origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin, Oxford, OUP Oxford, , 243 p. (ISBN 978-0-19-954906-1, lire en ligne)
- John F. Doebley, Brandon S. Gaut et Bruce D. Smith, « The Molecular Genetics of Crop Domestication », Cell, vol. 127, , p. 1309-1321 (DOI 10.1016/j.cell.2006.12.006, lire en ligne, consulté le )
- By Eva Jablonka et Gal Raz, « Transgenerational Epigenetic Inheritance: Prevalence, Mechanisms, and Implications for the Study of Heredity and Evolution », The Quarterly Review of Biology, vol. 84, , p. 131-176 (DOI 10.1086/598822, lire en ligne, consulté le )
- (en) Daniel Nätt, Carl-Johan Rubin, Dominic Wright et Martin Johnsson, « Heritable genome-wide variation of gene expression and promoter methylation between wild and domesticated chickens », BMC Genomics, vol. 13, , p. 59 (ISSN 1471-2164, PMID 22305654, PMCID 3297523, DOI 10.1186/1471-2164-13-59, lire en ligne, consulté le )
- Christine Künzl et Norbert Sachser, « The Behavioral Endocrinology of Domestication: A Comparison between the Domestic Guinea Pig (Cavia apereaf.porcellus) and Its Wild Ancestor, the Cavy (Cavia aperea) », Hormones and Behavior, vol. 35, , p. 28-37 (DOI 10.1006/hbeh.1998.1493, lire en ligne, consulté le )
- (en) DK Belyaev, Domestication, plant and animal, Encyclopaedia Britannica,
- (en) Bruce M. Carlson, Human Embryology and Developmental Biology, Elsevier Health Sciences, , 506 p. (ISBN 978-1-4557-2794-0 et 1-4557-2794-6, lire en ligne)
- (en) Brian K. Hall, The Neural Crest in Development and Evolution, Springer Science & Business Media, , 313 p. (ISBN 978-0-387-98702-6, lire en ligne)
- « Gilbert, Developmental Biology », sur www.sdbonline.org (consulté le )
- (en) Paul Trainor, Neural Crest Cells : Evolution, Development and Disease, Academic Press, , 488 p. (ISBN 978-0-12-404586-6, lire en ligne)
- Xiao Huang et Jean-Pierre Saint-Jeannet, « Induction of the neural crest and the opportunities of life on the edge », Developmental Biology, vol. 275, , p. 1-11 (DOI 10.1016/j.ydbio.2004.07.033, lire en ligne, consulté le )
- « Effects of domestication on the adrenal cortisol production in silver foxes during embryonic development », sur agris.fao.org (consulté le )
- (en) John W. Fondon et Harold R. Garner, « Molecular origins of rapid and continuous morphological evolution », Proceedings of the National Academy of Sciences, vol. 101, , p. 18058-18063 (ISSN 0027-8424 et 1091-6490, PMID 15596718, PMCID 539791, DOI 10.1073/pnas.0408118101, lire en ligne, consulté le )
- (en) Jiping Liu, Bin Cong et Steven D. Tanksley, « Generation and Analysis of an Artificial Gene Dosage Series in Tomato to Study the Mechanisms by Which the Cloned Quantitative Trait Locus fw2.2 Controls Fruit Size », Plant Physiology, vol. 132, , p. 292-299 (ISSN 1532-2548, PMID 12746534, PMCID 166974, DOI 10.1104/pp.102.018143, lire en ligne, consulté le )
- (en) Medard Ng et Martin F. Yanofsky, « Function and evolution of the plant MADS-box gene family », Nature Reviews Genetics, vol. 2, , p. 186-195 (ISSN 1471-0056, DOI 10.1038/35056041, lire en ligne, consulté le )
- (en) Bart J. Janssen, Kate Thodey, Robert J. Schaffer et Rob Alba, « Global gene expression analysis of apple fruit development from the floral bud to ripe fruit », BMC Plant Biology, vol. 8, , p. 16 (ISSN 1471-2229, PMID 18279528, PMCID 2287172, DOI 10.1186/1471-2229-8-16, lire en ligne, consulté le )
- (en) Bin Cong, Jiping Liu et Steven D. Tanksley, « Natural alleles at a tomato fruit size quantitative trait locus differ by heterochronic regulatory mutations », Proceedings of the National Academy of Sciences, vol. 99, , p. 13606-13611 (ISSN 0027-8424 et 1091-6490, PMID 12370431, PMCID 129721, DOI 10.1073/pnas.172520999, lire en ligne, consulté le )
- (en) Changbao Li, Ailing Zhou et Tao Sang, « Rice Domestication by Reducing Shattering », Science, vol. 311, , p. 1936-1939 (ISSN 0036-8075 et 1095-9203, PMID 16527928, DOI 10.1126/science.1123604, lire en ligne, consulté le )
- Melinda A. Zeder, « The Domestication of Animals », Journal of Anthropological Research, vol. 68, no 2, , p. 161–190 (ISSN 0091-7710 et 2153-3806, DOI 10.3998/jar.0521004.0068.201, lire en ligne, consulté le )
- Juliane Kaminski, Bridget M. Waller, Rui Diogo et Adam Hartstone-Rose, « Evolution of facial muscle anatomy in dogs », Proceedings of the National Academy of Sciences, vol. 116, no 29, , p. 14677–14681 (ISSN 0027-8424 et 1091-6490, DOI 10.1073/pnas.1820653116, lire en ligne, consulté le )
- Bridget M. Waller, Kate Peirce, Cátia C. Caeiro et Linda Scheider, « Paedomorphic Facial Expressions Give Dogs a Selective Advantage », PLoS ONE, vol. 8, no 12, , e82686 (ISSN 1932-6203, DOI 10.1371/journal.pone.0082686, lire en ligne, consulté le )
 Portail de la zoologie
Portail de la zoologie