Solanaceae
Solanacées

| Règne | Plantae |
|---|---|
| Sous-règne | Tracheobionta |
| Division | Magnoliophyta |
| Classe | Magnoliopsida |
| Sous-classe | Asteridae |
| Ordre | Solanales |
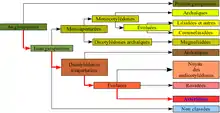
| Clade | Angiospermes |
|---|---|
| Clade | Dicotylédones vraies |
| Clade | Noyau des Dicotylédones vraies |
| Clade | Astéridées |
| Clade | Lamiidées |
| Ordre | Solanales |
Les Solanacées (Solanaceae Juss.) sont une famille de plantes dicotylédones (Magnoliopsida) appartenant à l'ordre des Solanales[2], dont le nom vient du genre Solanum. Ce sont des plantes herbacées, des arbustes, des arbres ou des lianes avec des feuilles alternes, simples et sans stipules. La famille comprend une centaine de genres pour environ 2 700 espèces[3] et occupe une grande diversité d'habitat, de morphologie et d'écologie. Cette famille cosmopolite est présente partout dans le monde à l'exception de l'Antarctique. Mais c'est en Amérique du Sud et en Amérique centrale que l'on rencontre le plus grand nombre d'espèces. Cette famille comprend des espèces alimentaires d'une grande importance économique telles que la pomme de terre (Solanum tuberosum), la tomate (Solanum lycopersicum), l'aubergine (Solanum melongena) et les piments (Capsicum). De nombreuses plantes ornementales très populaires appartiennent aux Solanacées : Petunia, Schizanthus, Salpiglossis et Datura. Certaines espèces, riches en alcaloïdes, sont mondialement connues pour leurs usages médicinaux, leurs effets psychotropes ou pour leur toxicité : belladone, morelle, brugmansia, datura, mandragore, tabac.
Étymologie
Le mot Solanacées vient du latin sōlānum qui désignait une morelle, probablement la morelle noire (Solanum nigrum L.). Sōlānum dérive du latin solari (« soulager »), référence aux feuilles émollientes ou aux propriétés narcotiques de la plante, ou de latin sōl (soleil), peut-être en raison de la forme de la fleur[4].
Paul-Victor Fournier y ajoute une référence au mot latin solanus, « vent d'est », qui, selon lui, signifie peut-être une origine orientale[5].
Répartition
Bien que la famille des Solanacées soit présente sur tous les continents, la plus grande richesse d'espèces se rencontre en Amérique centrale et en Amérique du Sud. Deux autres centres de diversité sont l'Australie et l'Afrique. Les Solanacées occupent une grande variété d'écosystèmes, des déserts aux forêts tropicales, et on les trouve souvent aussi dans la végétation secondaire colonisant les zones perturbées.
Description
Appareil végétatif
Les Solanacées sont des plantes herbacées, des sous-arbrisseaux, des arbustes, des arbres ou des lianes. Elles peuvent être annuelles, bisannuelles ou pérennes, d'un port érigé ou retombant. Elles peuvent être pourvues de tubercules souterrains. Elles ne présentent pas de canaux laticifères, ni latex, ni de suc coloré. Elles peuvent présenter une rosette de feuilles basale ou terminale ou ni l'une ni l'autre.
Les feuilles sont généralement alternes ou alternées à opposées (c'est-à-dire, alternes à la base de la plante et opposées vers l'inflorescence). La consistance des feuilles peut être molle ou coriace, ou transformées en épines. Elles sont alternes, entières à profondément lobées et sans stipules. En général, les feuilles sont pétiolées ou subsessiles, rarement sessiles. Elles sont fréquemment inodores, parfois aromatiques voire fétides. Les feuilles peuvent être simples ou composées ; dans ce dernier cas, elles peuvent être trifoliées ou pennatifides. Les nervations des feuilles sont réticulées et elles ne présentent pas de méristème basal. Les feuilles sont en général dorsiventrales, sans cavités excrétrices. Les stomates se trouvent en général confinés sur une face des feuilles, rarement sur les deux à la fois.
Appareil reproducteur
.jpg.webp)
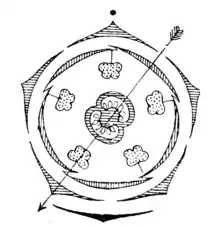
Les fleurs généralement hermaphrodites présentent un pistil et des étamines. Il existe des espèces monoïques, andromonoïques ou dioïques (par exemple, certains Solanum ou Symonanthus). La pollinisation est assurée par les insectes (entomophilie). Les fleurs peuvent être solitaires ou réunies en cymes terminales ou axillaires. Les fleurs, de taille moyenne, sont odorantes (comme Nicotiana), fétides (Anthocercis) ou inodores. Les fleurs sont actinomorphes (régulières), légèrement zygomorphes ou nettement zygomorphes (comme les fleurs avec corolle bilabiée dans le genre Schizanthus). Les irrégularités dans la symétrie de la fleur peuvent être dues à l'androcée, au périanthe ou aux deux à la fois. Les pétales sont soudés entre eux à des hauteurs variables pour donner la forme d'une étoile ou d'une trompette à la fleur. Comme pour les pétales, les sépales sont soudés à des hauteurs variables qui forment un tube qui présente cinq lobes au sommet. Les fleurs dans leur grande majorité présentent un périanthe différencié en calice et corolle (avec cinq sépales et cinq pétales, respectivement), un androcée avec cinq étamines et deux carpelles unis formant un gynécée avec un ovaire supère (on dit alors qu'elles sont pentamères et tétracycliques). Habituellement elles présentent un disque hypogyne. Le calice est gamosépale (puisque les sépales sont unis entre eux formant un tube), avec les cinq (parfois quatre ou six) segments égaux entre eux, il est pentalobé, avec les lobes plus courts que le tube, il est persistant et peut être fréquemment accrescent. La corolle habituellement présente cinq pétales qui se trouvent également unis entre eux formant un tube (corolle gamopétale). La corolle peut être campanulée, rotacée, infundibuliforme (en forme d’entonnoir) ou tubulaire. Le gynécée est bi-carpellaire (rarement 3- ou 5-loculaire), d'ovaire supère et présente deux locules. Les locules peuvent être secondairement divisés par des faux septum, comme dans le cas des Nicandreae et Datureae. Le gynécées est situé en position oblique par rapport au plan médian de la fleur. Elles présentent un unique style et un seul stigmate simple ou bilobé. Chaque locule contient 1 à 50 ovules anatropes ou hémianatropes de placentation axillaire. Le développement du sac embryonnaire peut être de type Polygonum ou du type Allium. Les noyaux polaires du sac embryonnaire peut être fusionné avant la fécondation. Ils présentent 3 antipodes, habituellement éphémère, ou persistantes comme dans le cas de Atropa. Les capsules sont normalement septicides ou rarement loculicide ou valvulaire.
Le fruit peut être charnu, c'est une baie (tomate), ou sec, c'est alors une capsule (Datura). Les graines sont habituellement endospermées oléagineuses (rarement amylacées), sans poil apparent. L'embryon, qui peut être droit ou courbe, présente deux cotylédons. Le formule chromosomique varie de x=7 à x=12. Beaucoup d'espèces sont polyploïdes.
Variabilité morphologique
En dépit de la description précédente, les Solanacées présentent une grande variabilité morphologique. Les exemples de la diversité de la famille sont :
- Le nombre de carpelles formant le gynécée :
- En général les Solanacées présentent un gynécée formée de deux carpelles. Toutefois, il y a des genres avec un gynécée monocarpellaire (Melananthus), ou avec 3 à 4 carpelles comme dans le genre Capsicum, ou 3 à 5 carpelles, comme dans le cas de Nicandra, certaines espèces de Jaborosa et de Trianaea. Enfin il existe au moins une espèce chez laquelle le gynécée possède 4 carpelles : Iochroma umbellatum.
- Le nombre de locules dans l'ovaire :
- Habituellement le nombre de locules est égal au nombre de carpelles. Toutefois, il existe des espèces chez lesquelles ces nombres ne sont pas égaux à cause de l'existence d'un faux septum (paroi interne qui subdivise chaque locule), comme le datura et certains membres de la tribu Lycieae (les genres Grabowskia et Vassobia).
- Les types d'ovules et leur nombre :
- Chez les Solanacées, les ovules sont en général anatropes. Toutefois, il y a des genres avec des ovules anacampilotropiques (exemple chez Phrodus, Grabowskia ou Vassobia), hémitropiques (Cestrum) hémicampilotropiques (Capsicum, Schizanthus et Lycium). Selon le nombre d'ovules par locules, ceux-ci sont en général nombreux, parfois peu (deux paires dans chaque locule chez Grabowskia, un pour chaque locule chez les Lycium) et, exceptionnellement, on trouve un seul ovule, comme pour Melenthus.
- Le type de fruit :
- Les fruits chez les Solanacées sont dans leur grande majorité des baies ou des capsules (dont des pyxides) et, moins fréquemment, des drupes. Les baies sont typiques des sous-familles des Cestroideae, des Solanoideae (à l'exception des genres Datura, Oryctus, Grabowskia et de la tribu des Hyoscyameae) et la tribu des Juanulloideae (à l'exception de Markea). Les capsules sont caractéristiques des sous-familles des Cestroideae (à l'exception des Cestrum) et des Schizanthoideae, les tribus des Salpiglossideae, des Anthocercidoideae et le genre Datura. La tribu des Hyoscyameae présente des pyxides. Les drupes sont typiques dans la tribu des Lycieae et chez les Iochrominae.
Composition chimique
Certaines espèces sont utilisées dans l'alimentation humaine (pomme de terre, tomate, aubergine, piment) tandis que d'autres sont très toxiques, particulièrement les Solanacées vireuses. Parmi ces dernières, la belladone, le datura, mandragore, de même que le tabac, ont été utilisées depuis des siècles pour leur propriétés psychotropes, qu'elles doivent à leur contenu en alcaloïdes, atropine, hyoscyamine et scopolamine chez les Solanacées vireuses, nicotine chez le tabac.
Toutes les Solanacées contiennent une quantité toxique de solanine, un alcaloïde stéroïde. Les baies mûres sont les parties les moins toxiques de ces plantes, mais leur ingestion peut être mortelle dans certaines circonstances. On trouve également de la solanine dans les germes et les taches vertes de la pomme de terre. Les espèces alimentaires contiennent aussi des alcaloïdes dans les feuilles, les tiges et les racines, mais les parties comestibles (tubercules de la pomme de terre, fruits des aubergines et piments) en sont dépourvues.
Alcaloïdes

Les alcaloïdes sont des bases azotées, produites par les plantes comme métabolites secondaires. Ils présentent une action physiologique sur les animaux certaine même à faible dose. Les tropanes sont les alcaloïdes les plus connues que l'on trouve chez les Solanacées. Les plantes qui contiennent ces substances ont été utilisées durant des siècles comme poisons. Cependant, malgré leur effet toxique reconnu, beaucoup de ces substances présentent des propriétés pharmaceutiques utiles. Les Solanacées sont caractérisées pour compter de nombreuses espèces contenant différents types d'alcaloïdes plus ou moins actifs ou toxiques, comme la scopolamine, l'atropine, l'hyoscyamine et la nicotine. On les trouve dans des plantes telles que la jusquiame blanche (Hyoscyamus albus), la belladone (Atropa belladonna), la stramoine (Datura stramonium), la mandragore (Mandragora autumnalis), le tabac. Quelques-uns des principaux types d'alcaloïdes de la famille des Solanacées sont les suivants:
- La solanine : glycoalcaloïde toxique de goût amer qui correspond à la formule brute C45H73NO15. Elle est composée d'un alcaloïde, la solanidine, et d'un oligoholoside. Elle se trouve naturellement dans les feuilles, les fruits et les tubercules de plusieurs Solanaceae, tels que la pomme de terre et la tomate. On considère que sa production est une stratégie adaptative des plantes en tant que mécanisme de défense contre les herbivores. L'empoisonnement à la solanine se caractérise par des troubles gastro-intestinaux (diarrhée, vomissements, douleurs abdominales) et neurologiques (maux de tête et des hallucinations). La dose toxique est de 2 à 5 mg par kilogramme de poids corporel. Les symptômes se manifestent de 8 à 12 heures après l'ingestion. La concentration de glycoalcaloïde dans la pomme de terre, par exemple, varie considérablement selon les conditions environnementales au cours de la culture, la durée de stockage et de la variété considérée. La concentration de glycoalcaloïde, en moyenne, est de 0,075 mg par gramme de pomme de terre.
- Les alcaloïdes tropaniques : composés organiques dérivés du tropane, un composé azoté bicyclique de formule brute C8H15N. Ce type d'alcaloïdes comprend, entre autres, l'atropine, la cocaïne. Ces alcaloïdes sont observés chez plusieurs espèces de Solanacées, comme la mandragore (Mandragora autumnalis), la jusquiame noire (Hyoscyamus niger), la belladone (Atropa belladonna) et la stramoine (Datura stramonium). L'atropine, mélange racémique de L- et R-hyoscyamine, provient de la belladone. Elle est utilisée pour dilater la pupille au cours des examens oculaires. En fait, le jus de ses baies a été utilisé par les courtisanes italiennes à la Renaissance pour exagérer la taille de leurs yeux en dilatant leurs pupilles. L'atropine est aussi un stimulant du système nerveux central. Il est utilisé comme un traitement pour l'empoisonnement au gaz neurotoxique ou aux insecticides organophosphorés. La scopolamine (à partir de Hyoscyamus muticus et Scopolia atropioides) est un autre tropane utilisé à petite dose pour traiter les nausées. La scopolamine et l'hyoscyamine sont des tropanes largement utilisés en pharmacologie et en médecine en raison de leurs effets sur le système nerveux parasympathique. Ces alcaloïdes ne peuvent être remplacés par une autre classe de composés pour lesquels la demande se poursuit. C'est une des raisons pour le développement d'un champ fertile de la recherche dans le métabolisme des alcaloïdes, des enzymes impliquées et les gènes qui les produisent. 6-hyoscyamine β-hydroxylase, par exemple, est l'enzyme qui catalyse l'hydroxylation de l'hyoscyamine conduisant à la production de la scopolamine, à la fin de la voie de biosynthèse des tropanes. Cette enzyme a été isolée et le gène correspondant a été cloné à partir de trois espèces, Hyoscyamus niger, Atropa belladonna et Brugmansia candida.
- La nicotine (nomenclature IUPAC (S)-3-(1-méthyl-2-pyrrolidinyl)pyridine) est un alcaloïde qui est produit en grandes quantités dans les plants de tabac (Nicotiana tabacum), mais il est aussi présent dans une moindre concentration chez d'autres espèces de Solanacées telles que le piment, la pomme de terre, la tomate . Leur fonction dans ces plantes est d'agir comme un moyen de défense contre les herbivores, étant un excellent neurotoxique, en particulier contre les insectes. En fait, la nicotine a été utilisée pendant de nombreuses années comme insecticide, actuellement remplacé par des molécules synthétiques dérivées de leur structure. À faibles concentrations, la nicotine agit comme un stimulant pour les mammifères, ce qui est la cause de la dépendance des fumeurs.
- La capsaïcine (nomenclature IUPAC 8-méthyle N-vanillyle 6-nonénamide) est un alcaloïde qui n'est pas structurellement apparenté à la nicotine ou avec les tropanes. On le trouve dans les espèces du genre Capsicum, lesquels comprennent les piments. C'est le composé actif qui détermine la saveur piquante de cette espèce. Le composé n'est pas particulièrement toxique pour l'être humain. Toutefois, il stimule les récepteurs spécifiques à la douleur chez la majorité des mammifères, particulièrement ceux qui sont relatifs avec la perception de la chaleur dans la muqueuse orale et d'autres tissus épithéliaux. En contact avec ces muqueuses, la capsaïcine provoque une sensation de brûlure qui n'est sans rappeler celle d'une brûlure réelle par le feu.
Classification et taxinomie
Cette famille de plantes a été décrite en 1789 par le botaniste français Antoine-Laurent de Jussieu.
La famille des Solanacées comprend environ 2 700 espèces[3].
Le Angiosperm Phylogeny Website [] accepte 102 genres, en incluant les familles des Duckeodendracées et des Nolanacées. L'espèce Duckeodendron cestroides est un grand arbre originaire des régions tropicales d'Amérique du Sud (Brésil). Les genres Coeloneurum, Espadaea, Goetzea, Henoonia sont des petits arbres ou des arbustes des grandes Antilles et d'Amérique du Sud. Les familles des Duckéodendracées et des Goetzéacées ne sont plus valides.
Avec plus de 1 500 espèces, le genre Solanum est le genre le plus important des Solanaceae et l'un des dix genres les plus riches en espèces chez les plantes à fleurs.
Synonymes
Selon GRIN (2 janvier 2020)[1] :
- Atropaceae Martinov
- Cestraceae Schltdl.
- Duckeodendraceae Kuhlm.
- Goetzeaceae Miers ex Airy Shaw
- Nolanaceae Bercht. & J. Presl, nom. cons.
- Salpiglossidaceae Hutch.
- Sclerophylacaceae Miers
Sous-familles
La famille est répartie en 7 ou 8 sous-familles selon les auteurs.
Liste des sous-familles
Selon Angiosperm Phylogeny Website (17 mars 2004)[6] :
- sous-famille des Cestroideae - Cestroidées avec 8 genres et 195 espèces répandues sur le continent américain.
- sous-famille des Goetzeoideae - Goetzéoidées avec 4 genres et 7 espèces originaires des Antilles.
- sous-famille des Nicotianoideae - Nicotianoidées avec 8 genres et 125 espèces originaires principalement d'Australie mais aussi du continent américain et d'Afrique.
- sous-famille des Petunioideae - Pétunioidées avec 13 genres et 160 espèces originaires d'Amérique centrale et du sud.
- sous-famille des Schizanthoideae - Schizanthoidées avec 1 genre et 12 espèces originaires du Chili.
- sous-famille des Schwenckioideae - Schwenckioidées avec 4 genres et 31 espèces originaires d'Amérique du Sud.
- sous-famille des Solanoideae - Solanoidées avec 61 genres et 1 725 espèces largement répandues dans le monde entier.
Selon Angiosperm Phylogeny Website (8 Jul 2010)[7] :
- sous-famille des Browallioideae
- sous-famille des Duckeodendron
- sous-famille des Goetzeoideae
- sous-famille des Nicotianoideae
- sous-famille des Petunioideae
- sous-famille des Schizanthoideae
- sous-famille des Schwenckioideae
- sous-famille des Solanoideae
Caractéristiques des sous-familles et tribus
Ce qui suit présente un résumé complet de la taxinomie des Solanacées, y compris sous-familles, tribus genres. Elle est fondée sur des études remontant à 2007, sur la systématique moléculaire de la famille[2],[3],[8],[9].
Cestroideae (syn.:Browallioideae)
La sous-famille des Cestroideae est caractérisée par la présence de fibres péricycliques, des androcées avec 4 ou 5 étamines, souvent didynames. Les formules chromosomiques de base varient de x = 7 à x = 13. La sous-famille comprend 8 genres (divisée en 3 tribus) et environ 195 espèces réparties en Amérique. Le genre Cestrum est le plus important en nombre d'espèces puisqu'il représente 175 des 195 espèces de la sous-famille . La tribu des Cestreae a la particularité d'inclure les taxons dont les chromosomes sont particulièrement long (de 7.21 à 11.51 µm de long), alors que le reste de la famille a généralement des chromosomes courts (1,5 à 3,52 µm chez les Nicotianoideae).
- Tribu des Browallieae Hunz. (1995)
- Browallia L. (1754), genre de 6 espèces néotropicales présentes jusqu'en Arizona (États-Unis).
- Streptosolen Miers (1850), genre monotypique natif des Andes.
- Tribu des Cestreae Don (1838). Comprend 3 genres de plantes ligneuses, généralement arbustives.
- Cestrum L. (1753). Comprend près de 175 espèces néotropicales.
- Sessea Ruíz & Pav. (1794). Genre comprenant 16 espèces andines.
- Vestia Willd. (1809), genre monotypique du Chili.
- Tribu des Salpiglossideae (Benth.) Hunz. cette tribu comprend 2 genres et 6 espèces endémiques d'Argentine et du Chili.
- Reyesia Gay (1840), comprend 4 espèces, présentes en Argentine et au Chili.
- Salpiglossis Ruíz & Pav. (1794), avec deux espèces de l'Amérique du Sud australe.
Goetzeoideae
La sous-famille des Goetzeoideae est caractérisée par une fructification en drupes et des graines avec des embryons courbes et des cotylédons grands et charnus. Le nombre chromosomique est x=13. Elle comprend 4 genres et 5 espèces répartis dans les Grandes Antilles. À partir de données moléculaires, certains auteurs classent à l'intérieur de cette sous-famille les genres monotypiques Tsoala Bosser & D'Arcy (1992), endémique de Madagascar, et Metternichia du sud-est du Brésil. Goetzeaceae Airy Shaw est considéré comme un synonyme de cette sous-famille[10].
Petunioideae
La systématique moléculaire montre que la sous-famille des Petunioideae est un clade frère des sous-familles avec un nombre chromosomique x=12 (Solanoideae et Nicotianoideae). Les espèces de la sous-famille des Petunioideae contiennent des calystégines, des alcaloïdes tropaniques. L'androcée est formé de 4 étamines (rarement 5). Le nombre chromosomique de cette famille peut être x=7, 8, 9 ou 11. Elle comprend 13 genres et 160 espèces, réparties en Amérique centrale et en Amérique du Sud. D'après les données moléculaires, certains auteurs considèrent que les genres monotypiques de Patagonie Benthamiella, Combera et Pantacantha forment le clade des Benthamielleae' qui devrait être inclus dans la sous-famille des Goetzeoideae.
- Benthamiella Speg. (1883), comprend 12 espèces natives de Patagonie ;
- Bouchetia Dunal (1852) comprend 3 espèces néotropicales ;
- Brunfelsia L. (1753), genre avec environ 45 espèces néotropicales ;
- Combera Sandw. (1936), genre deux espèces de Patagonie ;
- Fabiana Ruíz & Pav. (1794), genre comprenant 15 espèces natives des Andes ;
- Hunzikeria D'Arcy (1976), comprend 3 espèces du sud-ouest des États-Unis et du Mexique ;
- Latua Phil. (1858), avec une seule espèce du sud du Chili ;
- Leptoglossis Benth. (1845), genre avec 7 espèces de l'ouest de l'Amérique du Sud ;
- Nierembergia Ruiz & Pav. (1794), comprend 21 espèces sud-américaines ;
- Pantacantha Speg. (1902), genre monotypique de la Patagonie ;
- Calibrachoa Cerv. ex La Llave & Lex., avec 32 espèces néotropicales. Ce genre considérés, d'après les données morphologiques, comme appartenant au genre Petunia. L'analyse moléculaire et cytogénétique indique qu'il s'agit d'un genre distinct. En effet, si Calibrachoa possède un nombre chromosomique x=9, pour Petunia x=7[11],[12] ;
- Petunia (Juss.) Wijsman (1803), avec 18 espèces sud-américaines ;
- Plowmania Hunz. & Subils (1986), genre monotypique du Mexique et du Guatemala.
Schizanthoideae
La sous-famille des Schizanthoideae comprend des plantes herbacées annuelles ou bisannuelles, avec des alcaloïdes tropaniques, sans fibres péricycliques, avec des poils et des grains de pollen caractéristiques. Les fleurs sont zygomorphes. L'androcée présente deux étamines et 3 staminodes, la déhiscence des anthères est explosive. L'embryon est courbe. Le nombre chromosomique est x=10. Schizanthus est un genre assez atypique au sein des Solanacées, pour ses fleurs zygomorphes et pour son nombre chromosomique. Les données morphologiques et moléculaires montrent que Schizanthus est un genre frère du reste des Solanaceae et qui a divergé précocement du reste de la famille, probablement au Crétacé supérieur ou au Tertiaire inférieur, il y a 50 millions d'années[8],[9]. La grande diversification des types de fleurs dans le genre Schizanthus est le produit de l'adaptation des espèces de ce genre à différents groupes de pollinisateurs existant dans les écosystèmes méditerranéen, montagneux et désertique du Chili et des régions adjacentes de l'Argentine[13].
- Schizanthus Ruiz et Pav. (1794), comprend 12 espèces originaires du Chili.
Schwenckioideae
La sous-famille des Schwenckioideae comprend des plantes herbacées annuelles, avec des fibres péricycliques, les fleurs sont zygomorphes, l'androcée comporte 4 étamines, didynames ou avec 3 staminodes, l'embryon est droit et court. Le nombre chromosomique est x=12. Elle comprend 4 genres et 30 espèces réparties en Amérique du Sud ;
- Heteranthia Nees & Mart. (1823), une seule espèce du Brésil ;
- Melananthus Walp. (1850), comprend 5 espèces du Brésil, de Cuba et du Guatemala ;
- Protoschwenckia Soler. (1898), genre monotypique de Bolivie et du Brésil. Selon certaines études de systématique moléculaire, ce genre possède une position taxinomique incertaine au sein de la sous-famille.
- Schwenckia L. (1764), comprend 22 espèces néotropicales.
Nicotianoideae
La sous-famille des Nicotianoideae est subdivisée en plusieurs tribus :
- Tribu des Anthocercideae G.Don (1838). Cette tribu est endémique de l'Australie et comprend 31 espèces réparties en 7 genres. La systématique moléculaire de la tribu montre qu'elle est sœur de Nicotiana, que les genres Anthocercis, Anthotroche, Grammosolen, et Symonanthus sont monophylétiques, tandis que Cyphanthera et Duboisia ne le sont pas (Crenidium est monotypique). De plus on en a déduit que certains caractères sont dérivés à l'intérieur de la tribu, comme les étamines uniloculaires, avec opercules semi-circulaires, des fleurs ébractéolées, et des fruits en baies[14].
- Anthocercis Labill. (1806), 10 espèces, Australie
- Anthotroche Endl. (1839), 4 espèces, Australie
- Crenidium Haegi (1981), genre monotypique, Australie
- Cyphanthera Miers (1853), 9 espèces, Australie
- Duboisia R.Br. (1810), 4 espèces, Australie
- Grammosolen Haegi (1981), 2 espèces, Australie
- Symonanthus Haegi (1981), 2 espèces, Australie
- Tribu des Nicotianeae Dum. (1827)
- Nicotiana L. (1754), genre largement réparti, avec 52 espèces américaines, 23 australiennes et une africaine.
Solanoideae
La sous-famille des Solanoideae est subdivisée en plusieurs tribus et sous-tribus :
- Tribu des Capsiceae Dumort (1827)
- Capsicum L. (1753), genre comprenant près de 31 espèces néotropicales.
- Lycianthes (Dunal) Hassler (1917), près de 200 espèces réparties en Amérique et en Asie.
- Tribu des Datureae G. Don (1838) est une tribu qui comprend deux genres parfaitement différenciés tant au niveau morphologique qu'au niveau moléculaire. Brugmansia comprend des espèces arborescentes, tandis que Datura comprend des herbacées et des arbustes. Ce dernier a été divisé en 3 sections : Stramonium, Dutra et Ceratocaulis[15].
- Brugmansia Persoon (1805), genre comprenant 6 espèces des Andes. (incl.: Methysticodendron Schult. (1955))
- Datura L. (1753), avec 11 espèces néotropicales.
- Tribu des Hyoscyameae Endl. (1839)
- Anisodus Link (1825), avec 4 espèces de Chine, d'Inde de l'Himalaya.
- Atropa L. (1753), comprend 3 espèces eurasiatiques.
- Atropanthe Pascher (1909), genre monotypique originaire de Chine.
- Hyoscyamus L. (1753), comprend près de 20 espèces réparties de la Méditerranée jusqu'à la Chine.
- Physochlaina G. Don (1838), comprend 11 espèces eurasiatiques.
- Przewalskia Maxim. (1881) avec une seule espèce de Chine.
- Scopolia Jacq. (1764), genre avec une distribution disjointe, une espèce en Europe et une autre au Japon.
- Tribu des Jaboroseae Miers (1849)
- Jaborosa Juss. (1789), genre qui comprend 23 espèces sud-américaines.
- Tribu des Solandreae Miers (1849)
- Sous-tribu des Juanulloinae : elle comprend 10 genres d'arbres et d'arbustes épiphytes avec une répartition néotropicale[16]. Certains d'entre eux (Dyssochroma, Merinthopodium et Trianaea) présentent une claire dépendance avec diverses espèces de chauve-souris à la fois pour la pollinisation comme pour la dispersion des graines[17].
- Dyssochroma Miers (1849), avec 2 espèces du Sud du Brésil.
- Ectozoma Miers (1849)
- Hawkesiophyton Hunz. (1977)
- Juanulloa Ruiz et Pav. (1794), avec 11 espèces d'Amérique du Sud et d'Amérique centrale.
- Markea Rich.(1792) genre qui comprend 9 espèces d'Amérique du Sud et d'Amérique centrale.
- Merinthopodium J. Donn. Sm. (1897) inclut 3 espèces originaires d'Amérique du Sud.
- Rahowardiana D' Arcy (1973)
- Schultesianthus Hunz. (1977), genre comprenant 8 espèces néotropicales.
- Trianaea Planch. et Linden (1853) avec 6 espèces sud-américaines.
- Sous-tribu des Solandrinae, c'est une sous-tribu monotypique qui diffère de Juanulloinae par ses embryons avec cotylédons incombants et son ovaire semi-infère[16].
- Solandra Sw. (1787), comprend 10 espèces des régions néotropicales d'Amérique.
- Sous-tribu des Juanulloinae : elle comprend 10 genres d'arbres et d'arbustes épiphytes avec une répartition néotropicale[16]. Certains d'entre eux (Dyssochroma, Merinthopodium et Trianaea) présentent une claire dépendance avec diverses espèces de chauve-souris à la fois pour la pollinisation comme pour la dispersion des graines[17].
- Tribu des Lycieae Hunz. (1977). comprend 3 genres de plantes ligneuses qui croissent dans des climats arides ou semi-arides. Le genre cosmopolite Lycium est le plus ancien de la tribu et celui qui présente la plus grande variabilité morphologique[18]. Les études de systématiques moléculaires suggèrent que Grabowskia et Phrodus devraient être inclus à l'intérieur de Lycium[19] et que ce genre avec Nolana et Sclerophylax formerait un clade (Lyciina), lequel manque pour le moment de catégorie taxinomique[10]. Le fruit, une baie de couleur rouge, charnu, dispersé par les oiseaux, est le type de fruit dominant du genre Lycium. Les différents types de fruits ont évolué depuis la baie mentionnée plus tôt jusqu'à une drupe avec un nombre réduit de graines[20].
- Grabowskia Schltdl. (1832), 3 espèces sud-américaines.
- Lycium L. (1753), cosmopolite, comprend 83 espèces.
- Phrodus Miers (1849), endémique du nord du Chili, comprend 2 espèces.
- Tribu des Mandragoreae (Wettst.) Hunz. & Barboza (1995). Cette tribu monotypique n'a pas de position systématique définie selon ce que suggèrent les études de systématique moléculaire[10].
- Mandragora L. (1753), 2 espèces d’Eurasie
- Tribu des Nicandreae Wettst. (1891), il s'agit d'une tribu comportant deux genres sud-américains. Les études de systématique moléculaire montrent qu'ils n'ont aucun rapport entre eux ni avec d'autres genres de la famille, sa position taxinomique étant incertaine[10].
- Exodeconus Raf. (1838), avec 6 espèces de l'ouest de l'Amérique du Sud.
- Nicandra Adans (1763), genre avec une seule espèce dans les régions néotropicales.
- Tribu des Nolaneae Rchb. (1837)
- Nolana L. (1762) Ce sont des herbacées ou des petits arbustes dans la majorité aux feuilles succulentes, qui possèdent des fleurs très voyantes, qui vont du blanc jusqu'à divers tons de bleu, et son fruit schizocarpe. Comprend 89 espèces réparties dans l'ouest de l'Amérique du Sud.
- Tribu des Physaleae Miers (1849), c'est une grande tribu sœur de Capsiceae.
- Sous-tribu des Iochrominae (Miers) Hunz. est un clade à l'intérieur de la tribu Physaleae qui comprend 37 espèces, principalement réparties dans les Andes, assignées à 6 genres. Les membres de cette sous-tribu sont des arbustes ligneux ou des petits arbres avec des fleurs attractives tubulaires rotacées. Ils présentent, de plus, une grande diversité florale couvrant, de fait, la totalité de la variation existante dans la famille. Ses fleurs peuvent être rouges, orangées, jaunes, vertes, bleues, pourpres ou blanches. La forme de la corole peut être tubulaire à rotacée[21]
- Acnistus Schott (1829), avec une espèce néotropicale.
- Dunalia Kunth. (1818), qui comprend 5 espèces des Andes.
- Iochroma Benth. (1845), genre avec 24 espèces des Andes.
- Saracha Ruiz et Pav. (1794), qui comprend 2 espèces des Andes.
- Vassobia Rusby (1927), avec deux espèces sud-américaines.
- Eriolarynx Hunz.(2000), genre qui présente 3 espèces d'Argentine et de Bolivie.
- Sous-tribu des Physalinae (Miers) Hunz. (2000). C'est une sous-tribu monophylétique qui comprend 10 genres et inclut des herbacées ou des arbustes ligneux avec des fleurs solitaires, axillaires, jaunes, blanches ou pourpres qui sont pollinisées par les abeilles. Une fois que la pollinisation se produit, la corolle tombe et le calice se dilate jusqu'à couvrir entièrement la baie qui se développe (le calice est dit accrescent). Chez beaucoup d'espèces le calice devient jaune ou orange à maturité. Les baies, avec de nombreuses graines, sont de couleur verdâtre à jaune-orangé, fréquemment avec des reflets rouges ou pourpres[22].
- Brachistus Miers (1849), avec 3 espèces du Mexique et d'Amérique centrale.
- Chamaesaracha (A.Gray) Benth. et Hook. (1896), comprend 10 espèces du Mexique et Amérique centrale.
- Leucophysalis Rydberg (1896), comprend 3 espèces du sud-ouest des États-Unis et du Mexique.
- Margaranthus Schlecht. (1830), avec une espèce au Mexique.
- Oryctes S. Watson (1871), genre monotypique du sud-ouest des États-Unis.
- Quincula Raf. (1832) avec une seule espèce du sud-ouest des États-Unis et du Mexique.
- Physalis L. (1753), le genre le plus grand de la sous-tribu, avec 85 espèces réparties dans les régions tropicales d'Amérique et une espèce en Chine.
- Witheringia L'Heritier (1788), genre avec 15 espèces de régions néotropicales.
- Tzeltalia, genre séparé de Physalis, avec deux espèces réparties au Mexique et au Guatemala.
- Darcyanthus, genre avec une seule espèce originaire de Bolivie et du Pérou.
- Sous-tribu des Salpichroinae, est une sous-tribu de Physaleae qui comprend 16 espèces américaines distribuées en deux genres :
- Nectouxia Kunth. (1818), genre monotypique et endémique du Mexique.
- Salpichroa Miers (1845), genre avec 15 espèces des Andes d'autres régions d'Amérique du Sud.
- Sous-tribu des Withaninae, est une sous-tribu de vaste répartition, comprenant 9 genres :
- Archiphysalis Kuang (1966), avec 3 espèces de Chine et du Japon.
- Athenaea Sendtn. (1846), qui comprend 7 espèces du Brésil.
- Aureliana Sendt. (1846), avec 5 espèces d'Amérique du Sud.
- Melissia Hook. f. (1867), genre monotypique de Sainte Hélène.
- Physalisastrum Makino (1914), avec 9 espèces asiatiques.
- Tubocapsicum (Wettst.) Makino (1908), avec une seule espèce endémique de Chine.
- Withania Pauq.(1825), avec 10 espèces natives des îles Canaries, d'Afrique et du Népal.
- Cuatresia Hunz. (1977), avec 11 espèces néotropicales. Les études moléculaires montrent que ce genre, conjointement avec Deprea et Larnax possède une position taxinomique incertaine[10].
- Deprea Raf. (1838), avec 6 espèces néotropicales.
- Larnax Miers (1849), de nombreux taxinomistes les considèrent comme un synonyme de Deprea, comprend 22 espèces natives des Andes.
- Sous-tribu des Iochrominae (Miers) Hunz. est un clade à l'intérieur de la tribu Physaleae qui comprend 37 espèces, principalement réparties dans les Andes, assignées à 6 genres. Les membres de cette sous-tribu sont des arbustes ligneux ou des petits arbres avec des fleurs attractives tubulaires rotacées. Ils présentent, de plus, une grande diversité florale couvrant, de fait, la totalité de la variation existante dans la famille. Ses fleurs peuvent être rouges, orangées, jaunes, vertes, bleues, pourpres ou blanches. La forme de la corole peut être tubulaire à rotacée[21]
- Tribu des Solaneae (1852). Les genres Cyphomandra Sendtn. (1845), Discopodium Hochst. (1844), Normania Lowe (1872), Triguera Cav. (1786) Lycopersicum Mill ont été transférées à l'intérieur du genre Solanum. La sous-tribu est donc composée par deux genres[10] :
Liste des genres
Les genres avec position taxinomique incertaine (Incertae sedis) ne se trouvent toujours pas situés dans une des sous-familles reconnues de Solanaceae[10],[23]. En 2007 ce sont les suivants :
- Duckeodendron Kuhlmannb (1925), genre monospécifique d'Amazonie.
- Parabouchetia Baillon (1888)
- Pauia Deb. & Dutta (1965)
Selon Angiosperm Phylogeny Website (8 Jul 2010)[7] :
- Acnistus Schott
- Anisodus Link ex Spreng.
- Anthocercis Labill.
- Anthotroche Endl.
- Archiphysalis Kuang
- Athenaea Sendtn.
- Atrichodendron Gagnep.
- Atropa L.
- Atropanthe Pascher
- Aureliana Sendtn.
- Benthamiella Speg.
- Bouchetia Dunal
- Brachistus Miers
- Browallia L.
- Brugmansia Pers.
- Brunfelsia L.
- Calibrachoa Cerv.
- Capsicum L.
- Cestrum L.
- Chamaesaracha (A.Gray) Bentham
- Coeloneurum Radlk.
- Combera Sandwith
- Crenidium Haegi
- Cuatresia Hunz.
- Cyphanthera Miers
- Cyphomandra Mart. ex Sendtn.
- Datura L.
- Deprea Rafinesque
- Discopodium Hochst.
- Dittostigma Phil.
- Duboisia R. Brown
- Duckeodendron
- Dunalia Kunth
- Espadaea A. Richard
- Exodeconus Rafinesque
- Fabiana Ruiz & Pavon
- Goetzea Wydler
- Grabowskia Schltdl.
- Grammosolen Haegi
- Hawkesiophyton Hunz.
- Hebecladus Miers
- Henoonia Griseb.
- Hunzikeria D'Arcy
- Hyoscyamus L.
- Iochroma Bentham
- Jaborosa Juss.
- Jaltomata Schltdl.
- Juanulloa Ruiz & Pavon
- Latua Phil.
- Leptoglossis Bentham
- Leucophysalis Rydb.
- Lycianthes (Dunal) Hassl.
- Lycium L.
- Lycopersicon Mill.
- Mandragora L.
- Margaranthus Schltdl.
- Markea Rich.
- Melananthus Walp.
- Mellissia J. D. Hooker
- Metternichia J.C.Mikan
- Nectouxia Kunth
- Nicandra Adans.
- Nicotiana L.
- Nierembergia Ruiz & Pavon
- Nolana L.f.
- Nothocestrum A.Gray
- Oryctes S.Watson
- Pantacantha Speg.
- Parabouchetia Baillon
- Pauia Deb & R.M.Dutta
- Petunia Juss.
- Phrodus Miers
- Physalis L.
- Physochlaina G.Don
- Plowmania Hunz. & Subils
- Protoschwenckia Soler.
- Przewalskia Maxim.
- Quincula Rafinesque
- Rahowardiana D'Arcy
- Reyesia Gay
- Salpichroa Miers
- Salpiglossis Ruiz & Pavon
- Saracha Ruiz & Pavon
- Schizanthus Ruiz & Pavon
- Schultesianthus Hunz.
- Schwenckiopsis Dammer
- Schwenckia L.
- Sclerophylax Miers
- Scopolia Jacq.
- Sessea Ruiz & Pavon
- Sesseopsis Hassl.
- Solandra Sw.
- Solanum L.
- Streptosolen Miers
- Symonanthus Haegi
- Trianaea Planchon & Linden
- Triguera Cavanilles
- Tubocapsicum Makino
- Vassobia Rusby
- Vestia Willd.
- Withania Pauquy
- Witheringia L'Her.
Selon Tropicos (2 janvier 2020)[24] (Attention liste brute contenant possiblement des synonymes) :
- Agnistus G. Don
- Alibrexia Miers
- Alicabon Raf.
- Alkekengi Mill.
- Alona Lindl.
- Amatula Medik.
- Amphipleis Raf.
- Androcera Nutt.
- Anisodus Link ex Spreng.
- Anthocercis Labill.
- Anthotroche Endl.
- Antimion Raf.
- Apemon Raf.
- Aplocarya Lindl.
- Aquarta Cothen.
- Aquartia Jacq.
- Archihyoscyamus A.M. Lu
- Archiphysalis Kuang
- Armeniastrum Lem.
- Ascleia Raf.
- Athenaea Sendtn.
- Atropa L.
- Atropanthe Pascher
- Aureliana Sendtn.
- Bargemontia Gaudich.
- Bassova Cothen.
- Bassovia Aubl.
- Bassowia Dalla Torre & Harms
- Battata Hill
- Belenia Decne.
- Belladona Mill.
- Belladonna Scop.
- Bellinia Roem. & Schult.
- Benthamiella Speg.
- Blenocoes Raf.
- Boberella E.H.L. Krause
- Bosleria A. Nelson
- Bouchetia DC. ex Dunal
- Brachistus Miers
- Brachyhelus (Benth.) Post & Kuntze
- Brouvalia Adans.
- Brovallia L.
- Browala Cothen.
- Browallia L.
- Browallia guzmangoa S. Leiva
- Brugmansia Pers.
- Brunfelsia L.
- Brunfelsiopsis Urb.
- Calibrachoa Cerv.
- Calliphysalis Whitson
- Callomia
- Calydermos Ruiz & Pav.
- Cantalea Raf.
- Capsicophysalis Averett & M. Martínez
- Capsicum L.
- Ceratocaulos (Bernh.) Rchb.
- Cestrum L.
- Chaemaesaracha Dammer
- Chaenesthes Miers
- Chaetochilus Vahl
- Chaetochylus R. Hedw.
- Chamaesaracha (A. Gray) Benth. & Hook. f.
- Chamaesarachaea Kuntze
- Cleochroma Miers
- Cliocarpus Miers
- Codochonia Dunal
- Coeloneurum Radlk.
- Combera Sandwith
- Crabowskia G. Don
- Crenidium Haegi
- Cuatresia Hunz.
- Cyphanthera Miers
- Cyphomandra Mart. ex Sendtn.
- Darcyanthus Hunz. ex N.A. Harriman
- Dartus Lour.
- Datura L.
- Deprea Raf.
- Diamonon Raf.
- Dictyocalyx Hook. f.
- Dierbachia Spreng.
- Diplukion Raf.
- Discopodium Hochst.
- Diskion Raf.
- Dittostigma Phil.
- Dolia Lindl.
- Dolichosiphon Phil.
- Dorystigma Miers
- Duboisia R. Br.
- Duckeodendron Kuhlm.
- Dulcamara Moench
- Dunalia Kunth
- Dyssochroma Miers
- Eadesia F. Muell.
- Ectozoma Miers
- Elisia generic MILANO
- Entrecasteauxia Montrouz.
- Ephaiola Raf.
- Eplateia Raf.
- Eriolarynx (Hunz.) Hunz.
- Espadaea A. Rich.
- Eucapnia Raf.
- Evoista Raf.
- Exodeconus Raf.
- Fabiana Ruiz & Pav.
- Fontqueriella Rothm.
- Franciscea Pohl
- Fregirardia Dunal ex Delile
- Goetzea Wydler
- Goetzia Miers
- Grabowskia Schltdl.
- Grabowskya Endl.
- Grabuskia Raf.
- Grammosolen Haegi
- Gubleria Gaudich.
- Habrothamnus Endl.
- Hawkesiophyton Hunz.
- Hebecladus Miers
- Henoonia Griseb.
- Herschelia T.E. Bowdich
- Heteranthia Nees & Mart.
- Himenanthus Steud.
- Himeranthus Endl.
- Hiosciamus Neck.
- Hunzikeria D'Arcy
- Hyocyamus G. Don
- Hyoscarpus Dulac
- Hyosciamus Scop.
- Hyoscyamus L.
- Hyosicamus Hill
- Iochroma Benth.
- Jaborosa Juss.
- Jabrosa Steud.
- Jaltomata Schltdl.
- Jaltonia Steud.
- Jasminoides Duhamel
- Jochroma Wettstein in Engler & Prantl
- Juanulloa Ruiz & Pav.
- Kukolis Raf.
- Lamarkia G. Don
- Larnax Miers
- Latua Phil.
- Laureria Schltdl.
- Lehmannia Spreng.
- Leloutrea Gaudich.
- Leptofeddea Diels
- Leptofeddia Diels
- Leptoglossis Benth.
- Leptophragma Benth. ex Dunal
- Leucanthea Scheele
- Leucophysalis Rydb.
- Levana Raf.
- Lithophytum Brandegee
- Lomeria Raf.
- Lonchestigma Dunal
- Lycianthes (Dunal) Hassl.
- Lycioplesium Miers
- Lycium L.
- Lycomela Heist. ex Fabr.
- Lycopersicon Mill.
- Mandragora L.
- Manoelia Bowdich
- Marckea A. Rich.
- Marckia Post & Kuntze
- Margaranthus Schltdl.
- Markea Rich.
- Mathaea Vell.
- Mathea Steud.
- Matthissonia Raddi
- Megista Tourn.
- Meladendron Molina
- Melananthus Walp.
- Melissea Wettstein in Engler & Prantl
- Mellissia Hook. f.
- Melongena Mill.
- Merinthopodium Donn. Sm.
- Methysticodendron R.E. Schult.
- Metternichia J.G. Mikan
- Microschwenkia Benth.
- Mikroschwenkia Wettstein in Engler & Prantl
- Nectouxia Kunth
- Netouxia G. Don
- Neudorfia Adans.
- Nicandera Cothen.
- Nicandra Adans.
- Nicotia Opiz
- Nicotiana L.
- Nicotidendron Griseb.
- Nierembergia Ruiz & Pav.
- Nolana L. f.
- Normania Lowe
- Nothocestrum A. Gray
- Nycterium Vent.
- Oplukion Raf.
- Opsago Raf.
- Oryctes S. Watson
- Osteocarpus Phil.
- Pantacantha Speg.
- Panzeria J.F. Gmel.
- Papas F. M. Opiz in F. Berchtold & F. M. Opiz
- Parabouchetia Baill.
- Parqui Adans.
- Parquis Raf.
- Pauia Deb & R.M. Dutta
- Pederlea Raf.
- Pentaphiltrum Rchb.
- Perieteris Raf.
- Periloba Raf.
- Perizoma (Miers) Lindl.
- Petagnia Raf.
- Petunia Juss.
- Pheliandra Werderm.
- Phisalis Nocca
- Phrodus Miers
- Physaliastrum Makino
- Physalis L.
- Physaloides Moench
- Physochlaina G. Don
- Physoclaina Boiss.
- Pionandra Miers
- Pittunia Miers
- Plicula Raf.
- Plowmania Hunz. & Subils
- Poecilochroma Miers
- Polydiclis Miers
- Poortmannia Drake
- Portaea Sessé & Moc.
- Protoschwenkia Soler.
- Przewalskia Maxim.
- Pseudocapsicum Medik.
- Pseudodatura Zijp
- Pteroglossis Miers
- Pukanthus Raf.
- Puneeria Stocks
- Quincula Raf.
- Rahowardiana D'Arcy
- Rassovia R. Hedw.
- Rayera Gaudich.
- Reyesia Gay
- Rhopalostigma Phil.
- Saccardophytum Speg.
- Sairanthus G. Don
- Salpichroa Miers
- Salpiglossis Ruiz & Pav.
- Salpiglottis K. Koch
- Salpingoglottis K. Koch
- Saracha Ruiz & Pav.
- Sarachaea Kuntze
- Sarachra Dumort.
- Sarcophysa Miers
- Schizanthus Ruiz & Pav.
- Schraderanthus Averett
- Schuenkia Raf.
- Schultesianthus Hunz.
- Schwenckia D. Royen ex L.
- Schwenkea Cothen.
- Schwenkiopsis Dammer
- Scizanthus Pers.
- Sclerophylax Miers
- Scopolia Jacq.
- Scopolina Schult.
- Scubulon Raf.
- Sessea Ruiz & Pav.
- Sesseopsis Hassl.
- Siphaulax Raf.
- Siphonema Raf.
- Solanastrum Heist. ex Fabr.
- Solandra Sw.
- Solanocharis Bitter
- Solanopsis Börner
- Solanum L.
- Sorema Lindl.
- Sterrhymenia Griseb.
- Stimenes Raf.
- Stimomphis Raf.
- Stimoryne Raf.
- Stramonium Mill.
- Streptosolen Miers
- Streptostigma Regel
- Swartsia J.F. Gmel.
- Symonanthus Haegi
- Tabacum Gilib.
- Tabacus Moench
- Teganium Schmidel
- Teremis Raf.
- Thinogeton Benth.
- Trechonaetes Miers
- Trianaea Planch. & Linden
- Triguera Cav.
- Trinogeton Walp.
- Trompettia J. Dupin
- Trozelia Raf.
- Tsoala Bosser & D'Arcy
- Tubocapsicum (Wettst.) Makino
- Tula Adans.
- Tzeltalia E. Estrada & M. Martínez
- Ulloa Pers.
- Ulticona Raf.
- Valteta Raf.
- Vassobia Rusby
- Vateta Raf.
- Velpeaulia Gaudich.
- Vestia Willd.
- Vrolikia Spreng.
- Waddingtonia Phil.
- Wadea Raf.
- Walkeria Mill. ex Ehret
- Whitleya Sweet
- Withania Pauquy
- Witheringia L'Hér.
- Withleya Dumort.
- Zwingera Hofer
- × Eriochroma J.M.H. Shaw
- × Iozelia J.M.H. Shaw
Importance économique
Parmi la famille des Solanacées on trouve des espèces importantes pour l'alimentation humaine comme la pomme de terre (Solanum tuberosum), la tomate (Solanum lycopersicum), le piment (Capsicum annuum) et l'aubergine (Solanum melongena), mais aussi le tabac (Nicotiana tabacum).
Beaucoup de Solanaceae sont des adventices importantes dans diverses parties du monde. De nombreuses Solanacées peuvent être les hôtes de parasites et de maladies des cultures, et donc, leur présence augmente la perte de rendement ou de qualité du produit en raison de ces facteurs. Par exemple Browalia arborescens et Acnistus americana peuvent être les hôtes pour divers Thysanoptera qui endommagent la culture associée, et certaines espèces de Datura, des hôtes pour les différents types de virus qui sont ensuite transmis aux Solanacées cultivées. Certaines espèces de mauvaises herbes telles que Solanum mauritianum en Afrique du Sud, posent des problèmes écologiques et économiques si graves que certaines études envisagent le contrôle biologique à l'aide de certains insectes.
Plusieurs espèces d'arbres et d'arbustes de Solanacées sont cultivées comme plantes ornementales : Brugmansia ×candida, cultivée pour ses grandes fleurs pendantes en forme de trompette; Brunfelsia latifolia, dont les fleurs très parfumées passent du violet au blanc dans une période de trois jours. D'autres espèces arbustives cultivées pour leurs fleurs attrayantes sont Lycianthes rantonnetii aux fleurs bleu-violet; Nicotiana glauca aux fleurs jaunes, mais aussi les pétunia (Petunia ×hybrida), Lycium, Solanum, Cestrum, Calibrachoa ×hybrida, Solandra. Les pétunias (Petunia ×hybrida), Lycium, Solanum, Cestrum, Calibrachoa ×hybrida, Solandra, ainsi que le nothogenre ×Petchoa G. Boker & J. Shaw, sont des solanacée commercialisées comme plantes ornementales.
Beaucoup d'autres espèces, en particulier celles qui produisent des alcaloïdes sont utilisés en pharmacologie (Nicotiana, Hyoscyamus, et Datura).
Notes et références
- (es) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en espagnol intitulé « Solanaceae » (voir la liste des auteurs).
- USDA, Agricultural Research Service, National Plant Germplasm System. 2021. Germplasm Resources Information Network (GRIN Taxonomy). National Germplasm Resources Laboratory, Beltsville, Maryland., consulté le 2 janvier 2020
- Olmstead, R. G., J. A. Sweere, R. E. Spangler, L. Bohs, and J. D. Palmer. 1999. Phylogeny and provisional classification of the Solanaceae based on chloroplast DNA. Pp. 111-137. En: Solanaceae IV: advances in biology and utilization, M. Nee, D. E. Symon, R. N. Lester, and J. P. Jessop (eds.). The Royal Botanic Gardens, Kew Artículo en inglés
- Olmstead, R.G. and Bohs, L. 2007. A Summary of molecular systematic research in Solanaceae: 1982-2006. Acta Hort. (ISHS) 745:255-268
- François Couplan, Les plantes et leurs noms. Histoires insolites, Quae, , p. 74.
- Paul-Victor Fournier, Les quatre flores de la France : Corse comprise (Générale, Alpine, Méditerranéenne, Littorale), Paris, Lechevalier, , 1104 p. (ISBN 978-2-7205-0529-4), p. 751
- Angiosperm Phylogeny Website, consulté le 17 mars 2004
- Angiosperm Phylogeny Website, consulté le 8 Jul 2010
- Olmster, R.G. & J. Palmer. 1992. A chloroplast DNA phylogeny of the solanaceae: subfamilial relationships and character evolution. Annals Missouri Botanical Garden 79 (2):346-360. Resumen en inglés
- Martins TR, Barkman TJ (2005) Reconstruction of Solanaceae Phylogeny Using the Nuclear Gene SAMT. Systematic Botany: Vol. 30, No. 2 p. 435–447Resumen en inglés
- Olmstead, R.G. and Bohs, L. 2007. A Summary of molecular systematic research in Solanaceae: 1982-2006. Acta Hort. (ISHS) 745:255-268
- Ando, T, Kokubun, H., Marchesi, E., Suárez, E. & Basualdo, I. 2005. Phylogenetic Analysis of Petunia sensu Jussieu (Solanaceae) using Chloroplast DNA RFLP. Ann. Bot. 96(2): 289 - 297.Artículo en inglés
- Kei-Ichiro Mishiba, Toshio Ando, Masahiro Mii, Hitoshi Watanabe, Hisashi Kokubun, Goro Hashimoto, and Eduardo Marchesi. 2000. Nuclear DNA Content as an Index Character Discriminating Taxa in the Genus Petunia sensu Jussieu (Solanaceae). Ann Bot 85: 665-673. Artículo en inglés
- Perez, F., Arroyo, M., Medel, R. & M. Herskovitz. 2006. Ancestral reconstruction of flower morphology and pollination systems in Schizanthus (Solanaceae). American Journal of Botany 93(7): 1029–1038.
- (en) Vicente F. Garcia et Richard G. Olmstead, « Phylogenetics of Tribe Anthocercideae (Solanaceae) Based on ndhF and trnL/F Sequence Data », Systematic Botany, vol. 28, no 3, , p. 609-615 (ISSN 0363-6445, e-ISSN 1548-2324, OCLC 5550500276, DOI 10.1043/02-52.1, JSTOR 25063900, lire en ligne).
- Mace, E. S. C. G. Gebhardt, R. N. Lester. 1999. AFLP analysis of genetic relationships in the tribe Datureae (Solanaceae). Theoretical and Applied Genetics 99, (3-4): 634-641 Resumen en inglés
- Knapp, S., V. Persson & S. Blackmore. 1997. A Phylogenetic Conspectus of the Tribe Juanulloeae (Solanaceae). Annals of the Missouri Botanical Garden, Vol. 84, No. 1: 67-89 Résumé en anglais
- Sazima, M. ; Buzato, S.; Sazima, I. 2003. Dyssochroma viridiflorum (Solanaceae): a Reproductively Bat-dependent Epiphyte from the Atlantic Rainforest in Brazil. Annals of Botany. 92(5):725-730.
- Bernardello, L.M. 1987. Comparative Floral Morphology in Lycieae (Solanaceae). Brittonia, Vol. 39, No. 1:112-129 resumen en inglés
- Levin, R.A. & Miller, J.S. 2005. Relationships within tribe Lycieae (Solanaceae): paraphyly of Lycium and multiple origins of gender dimorphism. American Journal of Botany. 2005;92:2044-2053 resumen en inglés
- Bernardello, L. & F. Chiang-Cabrera. 1998.A cladistic study on the American species of Lycium (Solanaceae) based on morphological variation. Monographs in Systematic Botany from the Missouri Botanical Garden 68: 33–46.
- DeWitt Smith, S. & David A. Baum. 2006. Phylogenetics of the florally diverse Andean clade Iochrominae (Solanaceae). American Journal of Botany 93:1140-1153 Artículo en inglés
- Whitson, M. & P.S. Manos. 2005. Untangling Physalis (Solanaceae) from the physaloids: a two-gene phylogeny of the Physalinae. Systematic Botany 30: 216-230. Artículo en inglés
- Revisión de bibliografía realizada hasta noviembre de 2007
- Tropicos.org. Missouri Botanical Garden., consulté le 2 janvier 2020
Voir aussi
Bibliographie
- (fr) Georges Marchoux, Patrick Gognalons, Kahsay Gébré Sélassié, Virus des Solanacées : du génome viral à la protection des cultures, Paris, QUAE éditions, , 841 p. (ISBN 978-2-7592-0076-4, lire en ligne)
Liens externes
Références taxinomiques
- (fr) Référence Belles fleurs de France : Solanaceae (consulté le )
- (fr) Référence Belles fleurs de France 2 : Solanaceae (consulté le )
- (en) Référence Flora of China : Solanaceae (consulté le )
- (en) Référence Madagascar Catalogue : Solanaceae (consulté le )
- (en) Référence Flora of Chile : Solanaceae (consulté le )
- (en) Référence Angiosperm Phylogeny Website : Solanaceae () (consulté le )
- (en) Référence Jardin botanique du Missouri : Solanaceae (images de Madagascar)
- (en) Référence FloraBase (Australie-Occidentale) : classification Solanaceae (+ photos + distribution + description) (consulté le )
- (en) Référence DELTA Angio : Solanaceae Juss. (consulté le )
- (en) Référence Tree of Life Web Project : Solanaceae (consulté le )
- (en) Référence Catalogue of Life : Solanaceae Juss. (consulté le )
- (fr) Référence Tela Botanica (France métro) : Solanaceae (consulté le )
- (en) Référence Paleobiology Database : Solanaceae de Jussieu 1789 (éteint) (consulté le )
- (fr+en) Référence ITIS : Solanaceae (consulté le )
- (en) Référence World Register of Marine Species : taxon Solanaceae (+ liste espèces) (consulté le )
- (en) Référence NCBI : Solanaceae (taxons inclus) (consulté le )
- (en) Référence UICN : taxon Solanaceae (consulté le )
- (en) Référence GRIN : famille Solanaceae Juss. (+liste des genres contenant des synonymes) (consulté le )
- (en) Référence uBio : Solanaceae Juss. (consulté le )
Autres liens externes
- Solanaceae Source Site ressource sur les Solanacées au niveau international (hébergé par le Muséum d'Histoire Naturelle du Royaume-Uni) (en)
- http://didier-pol.net/8his-sol.htm
- CRB-Leg Présentation de la collection française de Solanacées à graines conservée par l'INRA
 Portail des Solanaceae
Portail des Solanaceae