Résistance des plantes aux maladies
La résistance des plantes aux maladies protège les plantes contre les agents pathogènes de deux manières : par des mécanismes pré-déterminés et par des réponses du système immunitaire induites par l'infection. Par rapport à une plante sensible à la maladie, la « résistance » est souvent définie comme la réduction de la croissance du pathogène sur la plante ou dans celle-ci, tandis que l'expression « tolérance aux maladies » décrit des plantes qui présentent moins de lésions dues à la maladie malgré des niveaux similaires de croissance du pathogène. Le déclenchement d'une maladie est déterminée par une interaction triple impliquant l'agent pathogène, la plante et le milieu (interaction connue sous le nom de « triangle des maladies »).
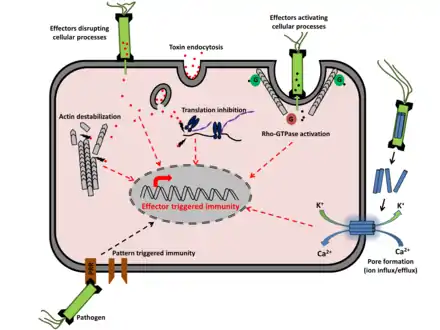
Les composés activateurs de défenses peuvent se déplacer de cellule à cellule et par voie systémique par le système vasculaire de la plante, mais les plantes n'ont pas de cellules immunitaires circulantes de sorte que la plupart des types cellulaires chez les plantes ont la capacité d'exprimer une vaste gamme de défenses antimicrobiennes.
Bien que des différences qualitatives évidentes de résistance aux maladies puissent s'observer quand on compare certaines plantes (ce qui permet de les classer comme « résistantes » ou « sensibles » après infection par la même souche pathogène à des niveaux d'inoculum pathogène similaires dans un environnement similaire), une gradation des différences quantitatives dans la résistance aux maladies est plus typiquement observée entre génotypes ou lignées de plantes. Les plantes sont presque toujours résistantes à certains pathogènes mais sensibles à d'autres ; la résistance est généralement spécifique d'une espèce ou d'une souche de pathogène.
Contexte
La résistance des plantes aux maladies est cruciale pour une production alimentaire fiable et permet des réductions significatives de l'utilisation à des fins agricoles de terres, d'eau, de carburants, et d'autres intrants. Les plantes, qu'elles appartiennent à des populations sauvages ou cultivées, sont porteuses de résistances inhérentes aux maladies, mais on connait de nombreux exemples de maladies de plantes aux effets dévastateurs (cf. Grande famine en Irlande, chancre du châtaignier), ainsi que des maladies récurrentes graves (cf. pyriculariose du riz, nématode à kystes du soja, chancre bactérien des agrumes). Cependant, la lutte contre les maladies protège raisonnablement la plupart des cultures. Elle est obtenue par l'utilisation de cultivars qui ont été sélectionnés pour leur bon niveau de résistance à de nombreuses maladies, et par des pratiques culturales telles que la rotation des cultures, l'emploi de semences saines (exemptes de pathogènes), le contrôle du calendrier et de la densité de plantation, la maîtrise de l'humidité du sol et l'emploi de pesticides. En prenant en compte de grandes régions et la plupart des espèces cultivées, on estime que les maladies réduisent généralement le rendement des plantes de 10 % chaque année dans les secteurs les plus développés, mais la perte de rendement dépasse souvent 20 % dans les secteurs moins développés.
Mécanismes communs
Systèmes immunitaires des plantes
Les systèmes immunitaires des plantes montrent des similitudes mécanistes avec ceux des insectes et des mammifères, mais présentent aussi de nombreuses caractéristiques spécifiques aux plantes.
Les plantes peuvent ressentir la présence de pathogènes et les effets de l'infection par des mécanismes différents de ceux des animaux[4].
Comme dans la plupart des réponses cellulaires à l'environnement, les défenses sont activées lorsque les protéines réceptrices détectent directement ou indirectement la présence d'agents pathogènes et déclenchent des réponses telles que l'ouverture du canal ionique, l'explosion respiratoire, des changements de redox cellulaire, des cascades de protéines kinases, et beaucoup d'autres réponses qui activent directement des modifications cellulaires (telles que le renforcement de la paroi cellulaire ou la production de composés antimicrobiens), ou activent des changements dans l'expression des gènes qui ensuite augmentent les réactions de défense des plantes.
Deux grands types de systèmes de détection des pathogènes sont observés dans les systèmes immunitaires des plantes : l'immunité de base, déclenchée par des motifs moléculaires associés aux pathogènes (PAMP), ou immunité PTI, et l'immunité déclenchée par les effecteurs (immunité ETI)[5],[6].
Les deux systèmes détectent différents types de molécules d'agents pathogènes, et ont tendance à utiliser différentes classes de protéines récepteurs des plantes pour activer les défenses anti-microbiennes. Bien que divers exemples concrets de mécanismes de détection plantes-parasites maintenant connus soient contraires à une classification claire comme PTI ou ETI[7], les études de nombreuses interactions plante-pathogène vont dans le sens du maintien des concepts PTI / ETI.
Mécanismes de signalisation
Facteurs de transcription et réponse hormonale
On a identifié de nombreux gènes, ou protéines, qui assurent la médiation de la transduction du signal de défense des plantes[8]. Le cytosquelette et la dynamique du trafic vésiculaire aident à cibler les réactions de défense des plantes asymétriquement dans les cellules végétales, vers le point de l'attaque pathogène.
Mécanismes des hormones et facteurs de transcription
La plus grande partie de l'activité de la plante est régulée par des hormones de signalisation telles que[9] :
- l'acide salicylique (croissance, développement, transport),
- l'acide jasmonique (réponse au stress, inhibition de la croissance, développement floral),
- l'éthylène (régulation de la croissance, signalisation, etc.)
Les messages de signalisation peuvent aussi être régulés par signalisation ionique, dégradation, ou contre-réaction. Les réponses de transcription de ces facteurs induisent généralement une réponse du gène primaire, qui déclenche une réponse du gène secondaire, et finalement l'interaction diaphonique des deux provoque une résistance systémique acquise (RSA contrôlée par l'acide salicylique) ou une résistance systémique induite (RSI contrôlée par l'acide jasmonique et l'éthylène) chez la plante[10]. On pense que ces réponses sont régulées par des impulsions de transcription, qui peuvent être causées par un changement dans les réactions d'oxydo-réduction qui dictent ces impulsions dans un délai de 24 à 48 heures. Ces impulsions sont mutuellement exclusives au sein d'une même cellule, autorisant l'activation des facteurs de transcription avant l'induction de la RSA[9].
Ces hormones sont impliquées dans le processus de « priming » (préparation des défenses des végétaux à la suite de leurs interactions avec des microorganismes de l’environnement), phase rapide aérienne (quelques heures) qui leur permettent de résister à des attaques ultérieures d'agents pathogènes. Ce processus de communication inter-plantes via des composés organiques volatils reste encore mal connu. Il met en œuvre l'élicitation de la phase suivante, un déploiement de la défense via des signaux relais vasculaires (émission d'hormones de défense qui se compte en jours ou en semaines)[11],[12].
Sélection des plantes pour la résistance aux maladies
Les sélectionneurs concentrent une part importante de leurs efforts sur la sélection et le développement de lignées de plantes résistantes aux maladies. Les maladies des plantes peuvent aussi être partiellement maîtrisées par le recours à des pesticides et par des pratiques culturales telles que la rotation des cultures, travail du sol, densité de plantation, achat de semences exemptes de maladies et nettoyage des équipements, mais les variétés végétales ayant une résistance inhérente (déterminée génétiquement) aux maladies sont généralement le premier choix pour lutter contrer les maladies. La sélection pour la résistance de la maladie est aussi ancienne que la domestication des plantes, mais elle exige un effort permanent. Les raisons en sont que les populations d'agents pathogènes acquièrent souvent une virulence accrue par sélection naturelle, que de nouveaux agents pathogènes peuvent être introduits dans une zone donnée, que les méthodes de culture peuvent augmenter l'incidence des maladies au fil du temps, que des changements dans la pratique de la culture peuvent favoriser de nouvelles maladies et que l'amélioration des plantes pour d'autres traits peut perturber la résistance aux maladies qui était présente dans les variétés plus anciennes. Une lignée de plantes ayant une résistance acceptable contre un agent pathogène peut aussi manquer de résistance face à d'autres pathogènes.
Gamme d'hôtes
Il existe des milliers d'espèces de micro-organismes pathogènes des plantes, mais seule une petite minorité d'entre eux ont la capacité d'infecter un large éventail d'espèces végétales. La plupart des agents pathogènes présentent plutôt un haut degré de spécificité de l'hôte. On dit souvent que les espèces de plantes non-hôtes « expriment une résistance non-hôte ». On parle de « résistance hôte » lorsqu'une espèce de plante est sensible à un pathogène, mais que certaines souches de cette espèce végétale résistent à certaines souches du pathogène. Il peut y avoir un chevauchement dans les causes de résistance hôte et de résistance non-hôte. La gamme d'hôtes d'un pathogène peut changer très soudainement si, par exemple, la capacité à synthétiser une toxine ou un effecteur spécifique de l'hôte est acquise par réarrangement génétique / mutation, ou par transfert horizontal de gènes d'un organisme apparenté ou relativement non-apparenté.
Épidémies et biologie des populations
Dans les populations sauvages, les plantes se caractérisent souvent par une importante diversité du génotype et des populations dispersées (croissance en mélange avec de nombreuses autres espèces végétales). Elles ont également subi des millions d’années de coévolution avec les pathogènes des plantes. De ce fait, tant que des pathogènes nouveaux ne sont pas introduits en provenance d’autres parties du monde, les populations naturelles de plantes ne présentent généralement qu’une faible incidence d’épidémies graves.
Dans les systèmes agricoles, l’homme cultive souvent une seule espèce de plante à haute densité, avec de nombreuses parcelles de cette même espèce dans une région donnée, et avec une diversité génétique significativement réduite tant à l’intérieur des parcelles qu’entre parcelles. En outre, le déplacement rapide des personnes et des marchandises sur de longues distances augmente le risque d’introduction de pathogènes contre lesquels les plantes n’ont pas été sélectionnées pour la résistance. Le changement climatique peut modifier la répartition géographique des espèces pathogènes et faire que certaines maladies deviennent un problème dans des zones où elles étaient précédemment moins importantes. Ces facteurs rendent l'agriculture moderne particulièrement sujette à des épidémies.
Les solutions usuelles de ces problèmes comprennent la sélection constante pour la résistance aux maladies, l’emploi de pesticides pour supprimer les épidémies potentielles récurrentes, des contrôles aux frontières et des restrictions à l’importation des plantes, le maintien d’une diversité génétique substantielle au sein du pool génique des plantes cultivées (cf. Diversité des plantes cultivées), et une surveillance constante des problèmes de maladies pour faciliter le déclenchement précoce de réponses appropriées. Certaines espèces de pathogènes sont connues pour avoir une plus grande capacité que d'autres à surmonter la résistance des plantes, souvent à cause de leur aptitude à évoluer rapidement et à se disperser largement[13].
Notes et références
- PAMPs-triggered immunity ou immunité déclenchée par les PAMPs.
- Effector-triggered immunity ou immunité déclenchée par les effecteurs (en).
- Luciano Paolozzi, Jean-Claude Liébart, Matthieu Arlat, Michel Dion, Harivony Rakotoarivonina, Introduction à la microbiologie. Microbiologie fondamentale et appliquée, Dunod, (lire en ligne), p. 183
- (en) Jade Boyd, « A bit touchy: Plants' insect defenses activated by touch. Rice University », .
- (en) J.D. Jones et J.L. Dangl, « The plant immune system », Nature, vol. 444, , p. 323–329.
- (en) P. Dodds et J. Rathjen, « Plant immunity: towards an integrated view of plant–pathogen interactions », Nature Reviews Genetics, vol. 11, , p. 539.
- (en) B. Thomma, T. Nurnberger et M. Joosten, « Of PAMPs and Effectors: The Blurred PTI-ETI Dichotomy », The Plant Cell, vol. 23, , p. 4–15.
- (en) KE Hammond-Kosack et JE Parker, « Deciphering plant-pathogen communication: fresh perspectives for molecular resistance breeding », Current Opinion in Biotechnology, Elsevier, vol. 14, no 2, , p. 177-193.
- (en) J. W. Moore, « Transcription Dynamics in Plant Immunity », The Plant Cell, vol. 23, no 8, , p. 2809–2820 (DOI 10.1105/tpc.111.087346).
- (en) Jean-Pierre Métraux, Christiane Nawrath, Thierry Genoud, « Systemic acquired resistance », Euphytica, vol. 124, no 2, , p. 237–243 (DOI 10.1023/A:101569070)
- (en) Deepak G. Panpatte, Yogeshvari K. Jhala, Rajababu V. Vyas, Harsha N. Shelat, Microorganisms for Green Revolution, Springer, (lire en ligne), p. 316
- (en) M. Heil, J. Ton, « Long-distance signalling in plant defence », Trends Plant Sci., vol. 13, no 6, , p. 264-272 (DOI 10.1016/j.tplants.2008.03.005)
- (en) Bruce A. McDonald et Celeste Linde, « Pathogen population genetics, evolutionary potential, and durable resistance », Annual Review of Phytopathology, vol. 40, , p. 349-379 (DOI 10.1146/annurev.phyto.40.120501.101443).
Voir aussi
Articles connexes
Bibliographie
- (en) J.A. Lucas, Plant Defence." Chapter 9 in Plant Pathology and Plant Pathogens, Blackwell Science, , 3e éd. (ISBN 0-632-03046-1)
- (en) K. Hammond-Kosack et J.D.G. Jones, Responses to plant pathogens.In: Buchanan, Gruissem et Jones, eds. Biochemistry and Molecular Biology of Plants., Rockville, MD., Amer. Soc. Plant Biol., (ISBN 0-943088-39-9)
- (en) P. Dodds et J. Rathjen, Plant immunity: towards an integrated view of plant–pathogen interactions., Nature Reviews Genetics 11:539, 2010..
- (en) G. Schumann, Plant Diseases: Their Biology and Social Impact., St. Paul, MN, APS Press, (ISBN 0-89054-116-7)
Liens externes
 Portail de l’agriculture et l’agronomie
Portail de l’agriculture et l’agronomie  Portail de la botanique
Portail de la botanique  Portail de la protection des cultures
Portail de la protection des cultures