Réplication de l'ADN
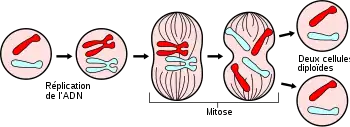
La réplication de l'ADN, appelée aussi duplication de l'ADN, est le processus au cours duquel l'ADN est synthétisé grâce à l'ADN polymérase. Ce mécanisme permet d'obtenir, à partir d'une molécule d'ADN, deux molécules identiques à la molécule initiale, en vue de leur distribution aux deux cellules filles pendant la mitose.
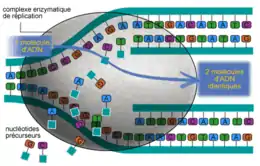
Puisque les deux chaînes de l'ADN parental se séparent et que deux nouvelles molécules sont formées, on qualifie ce processus de semi-conservatif.
L'ADN a l'importante propriété de pouvoir être reproduit à l'identique, ce qui permet à l'information génétique de se transmettre d'une cellule mère aux cellules filles lors de la division cellulaire. La molécule d'ADN s'ouvre comme une fermeture éclair (ou tirette — par rupture des liaisons hydrogène entre bases appariées de liaisons faibles) libérant deux brins complémentaires. Chaque brin solitaire catalyse alors la synthèse de sa moitié manquante, intégrant, selon la règle de complémentarité des bases, des nucléotides qui sont dispersés dans le noyau. Ainsi, chaque nouvelle molécule est identique à la molécule d'ADN initiale.
La réplication va commencer à des endroits précis : les origines de réplication. Des protéines, les facteurs d'initiation de la réplication vont reconnaître ces endroits. Le rôle de ces facteurs d'initiation est de faciliter la fixation d'autres protéines qui elles aussi vont se fixer à ces origines de réplication. Ces protéines permettent l'ouverture des deux brins de l'ADN et ainsi « faire apparaître » des fourches de réplication.
Un grand nombre de protéines interviennent dans le mécanisme moléculaire de la réplication de l’ADN formant le complexe enzymatique de réplication, appelé réplisome[1],[2]. Les enzymes et protéines intervenant dans la réplication de l’ADN sont homologues chez les eucaryotes et chez les archées mais ont des séquences en acides aminés très différentes chez les bactéries. Toutefois, au niveau fonctionnel comme au niveau structural, on retrouve des homologies frappantes entre les protéines bactériennes et les protéines eucaryotes, ce qui indique que les mécanismes de réplication sont analogues[3],[4].
Réplication de l’ADN : semi-conservatrice et bidirectionnelle
Le brin d’ADN qui sert de matrice à la réplication est le brin parental. Le nouveau brin complémentaire au brin parental est le brin néoformé. À l’issue de la réplication, chacune des deux molécules d’ADN nouvellement formée est constituée d’un brin parental et d’un brin néoformé. On qualifie ce processus de semi-conservateur. C'est l'expérience de Meselson et Stahl en 1958 qui a permis de le démontrer[5].
La réplication de l’ADN débute à partir d’une origine de réplication et progresse dans les deux sens à partir de ce point créant ainsi deux fourches de réplication. On dit que la réplication de l’ADN est bidirectionnelle.
Œil de réplication
En biologie, le terme d'œil de réplication est utilisé pour désigner la forme représentée par la molécule d'ADN lors de sa réplication. Cette réplication se fait lorsque l'ADN est à l'état de chromatine, pendant la phase S de l’interphase. L'ADN prend normalement la forme de deux brins entortillés en double hélice, mais pendant la réplication, cette forme change énormément. En effet, la réplication commence grâce à une ou plusieurs origines de réplication qui sont des séquences de nucléotides spécifiques reconnues par des protéines de réplication. Ces protéines vont s'attacher aux origines de réplication et séparer les deux brins d'ADN ce qui va former un « œil » de réplication.
On remarque que chaque nucléofilament (c'est-à-dire molécule d'ADN) se réplique et donne deux nucléofilaments identiques, reliés au niveau du centromère. Cette réplication qui conserve un brin de la molécule d'ADN initiale dans chacune des molécules d'ADN fille est dite semi-conservative. On parle alors de chromosome constitué de deux chromatides. Pour plus de détails, consultez l'interphase.
Fourche de réplication
La fourche de réplication est une transformation de la forme de l'ADN. Lorsqu'un œil de réplication est créé d'autres enzymes interviennent, les hélicases. Ces enzymes vont se fixer aux extrémités de l’œil de réplication et la topoisomérase commence à dérouler l'ADN pour enlever les supers tours. L'hélicase va détacher les deux brins, pour pouvoir augmenter la taille de l’œil de réplication. En se faisant les hélicases vont transformer les extrémités des yeux de réplication en forme de Y, appelées fourches de réplication.
La fourche de réplication est la structure formée lorsque l’ADN se réplique, et sur laquelle l'ADN polymérase vient se fixer. L'ADN polymérase est une enzyme catalysant la formation des liaisons nucléotidiques. Le complexe enzymatique intervenant dans la réplication est appelé réplicase.
La réplication peut être divisée en trois étapes principales : l’initiation, l’élongation et la terminaison.
Initiation de la réplication
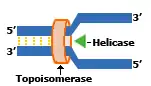
L’initiation de la réplication a lieu à l’origine de réplication. Il n’y a qu’une seule origine de réplication dans les chromosomes bactériens alors qu’il en existe plusieurs chez les eucaryotes (chez les Archaea, le nombre d’origines varie entre une et trois selon les espèces). La première étape débute par la fixation de la protéine DnaA (protéine capable de reconnaître ces origines) sur l'origine de réplication. Il s'ensuit un enroulement de l'ADN autour de la protéine qui provoque une dénaturation locale des deux brins, au niveau des 3 répétitions consécutives de 13bp. Une hélicase s'y engouffre et sépare les brins dans les deux sens en rompant les liaisons hydrogène (ou « ponts hydrogènes ») entre les deux brins de la double hélice d’ADN. Deux fourches de réplication sont ainsi créées, délimitant l'œil de réplication. D’autres protéines se lient à l’ADN simple brin (monocaténaire) ainsi formé et évitent la reformation de la double hélice. Ce sont les protéines SSB (single strand binding) chez les bactéries et les protéines RPA chez les eucaryotes et les archées. De part et d'autre des deux fourches de réplications, un gyrases (topoisomérase de classe II) enlève les supertours positifs engendrés par la formation de l'œil de réplication.
Chez les procaryotes, le complexe multienzymatique composé de l'hélicase et de la primase est appelé le primosome[6].
Élongation ou la synthèse d’ADN
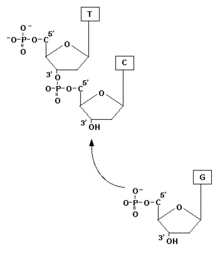
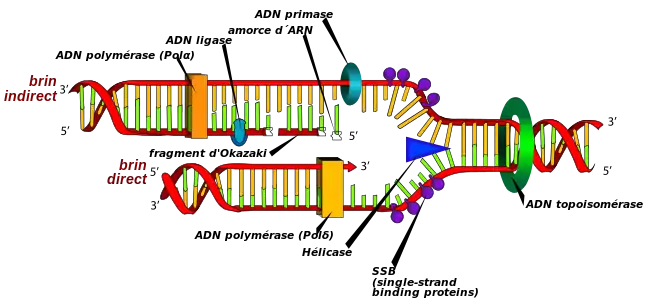
C’est au cours de cette phase qu’il y a formation du réplisome et synthèse d’ADN. L’élongation de l’ADN progresse toujours dans le sens 5' vers 3' pour le brin en création. C’est l'ADN polymérase, qui ajoute à l'extrémité 3' de la molécule en formation, des désoxyribonucléotides. Cependant, les deux brins de la double hélice d’ADN sont enroulés dans des sens opposés : ils sont antiparallèles. Il existe de ce fait des mécanismes différents selon le brin d’ADN répliqué.
Il existe ainsi un « brin direct », ou « brin précoce » (leading strand) et un « brin indirect », ou « retardé », ou « tardif » (lagging strand) :
- le « brin direct » est le brin complémentaire du brin parental orienté 3’ vers 5’ (le « brin direct » est donc orienté 5’ vers 3’). Il est donc créé de façon continue, dans le sens 5’ vers 3’ ;
- le « brin indirect » est le brin complémentaire du brin parental orienté 5' vers 3’ (le « brin indirect » est donc orienté 3’ vers 5’). Il est créé de façon discontinue, sous forme de fragments d’Okazaki, dans le sens 3’ vers 5’.
→ Chez le procaryote, les fragments d'Okazaki mesurent de 1000 à 2000 bases, et chez l'eucaryote ils sont d'environ 200 bases.
L'ADN polymérase a besoin d'une amorce pour fonctionner, parce qu'elle ne commence à synthétiser que par une extrémité 3'OH Libre. Et c'est l'amorce ARN qui va fournir cette extrémité libre. La présence ici d'ARN est expliquée par le fait que seules deux enzymes peuvent synthétiser les chaînes nucléotidiques : L'ARN Polymérase et L'ADN Polymérase. Dans le cas présent, l'ADN Polymérase ne pouvant fonctionner sans amorce, c'est L'ARN qui prend le relai pour fournir l'amorce nécessaire. La primase va en effet créer ces amorces d'ARN. Il y aura donc sur le brin retardé des jonctions ARN-ADN, qui seront par la suite éliminées par une RNase H. Des ADN polymérases particulières vont ensuite combler les lacunes laissées par l'ARN.
D'autres enzymes sont nécessaires au bon fonctionnement de la réplication. Puisque le placement des histones dans la chromatine régule l'expression génétique, des protéines chaperon séparent les histones de l'ADN avant la réplication et les remettent après aux mêmes endroits[7]. Les polymérases sont retenues à l'ADN parental par des protéines tenons et des protéines à pince coulissante. C’est le PCNA des eucaryotes et des archées, et la sous unité b des bactéries. Une hélicase brise les liaisons H entre les deux brins, et des protéines SSB se fixent à l'ADN monocaténaire par des liaisons salines, pour éviter la reformation de liaisons entre les deux brins. Sur l'ADN double brin précédant l'hélicase se fixe une topoisomérase I qui va permettre d'éviter les torsions entraînées par l'ouverture de la double-chaîne par l'hélicase (comme pour une ficelle dont on écarte les deux brins), en coupant un des brins, puis le ressoudant après déroulement. Une topoisomérase II va se fixer sur une des molécules d'ADN filles, et par la scission des deux brins de celle-ci, va permettre le démêlement des 2 ADN filles. Elle ressoude ensuite (après le passage de l'autre molécule dans l'interstice formé) la molécule lysée.
Terminaison
Cette phase correspond à l’arrêt de la réplication lorsque deux fourches de réplication se rencontrent ou lorsqu’une fourche rencontre un signal de terminaison de la réplication. Il y a « ter » : terA terD terB terC qui freinent les fourches de réplication.
Fidélité de la réplication
La fidélité de réplication est très grande et elle est en très grande partie due à l'ADN polymérase, qui incorpore les bases nucléiques en fonction de la complémentarité des appariements Watson-Crick (A-T, C-G). En effet, ce très faible taux d'erreur de l'ADN polymérase pendant la réplication est dû au fait que les ADN polymérases réplicatives disposent d'une activité de relecture qui leur permet de vérifier que le dernier nucléotide incorporé est bien le bon.
En cas d'erreur, les ADN polymérases (I et III) les corrigent au moyen de leur activité exonucléase 3' → 5'. Ceci permet à la polymérase de reculer d'un cran en éliminant le nucléotide incorrect qui est hydrolysé, pour ensuite reprendre la synthèse du brin d'ADN.
Avant cette relecture de l'ADN le taux d’erreur est d'environ 10-5 (une erreur sur cent mille bases répliquées) ce qui est très peu compte tenu du nombre de bases répliquées par seconde qui est d'environ 500 bases par seconde chez les bactéries. L'étape de relecture fait chuter le taux d'erreur de réplication à environ 10-7. Enfin, les erreurs qui ont éventuellement échappé à ce mécanisme de contrôle sont réparées ensuite dans l'ADN double brin par un mécanisme spécifique de réparation des mésappariements ou MR (mismatch repair). L'ensemble du dispositif constitue donc un système extrêmement efficace puisque le taux d'erreur chute à environ une mutation pour 10 milliards de bases répliquées.
Notes et références
- Bénédicte Michel et Giuseppe Baldacci, « Réplication », Médecine/sciences, vol. 14, no 12, , p. 1422-1427 (ISSN 0767-0974, lire en ligne)
- (en) Pomerantz R.T., O'Donnell M., « Replisome mechanics: insights into a twin DNA polymerase machine. », Trends Microbiol., vol. 15, , p. 156-164 (PMID 17350265)
- (en) Indiani C., O'Donnell M., « The replication clamp-loading machine at work in the three domains of life. », Nat. Rev. Mol. Cell. Biol., vol. 7, , p. 751-761 (PMID 16955075)
- (en) Baker T.A., Bell S.P., « Polymerases and the replisome: machines within machines. », Cell, vol. 92, , p. 295-305 (PMID 9476890)
- « Expérience de Meselson et Stahl », sur Planet-Vie (consulté le ).
- Alberts 1987
- Ransom M, Dennehey B, Tyler JK, « Chaperoning histones during DNA replication and repair », Cell, vol. 140, no 2, , p. 183-195 (PMID 20141833, DOI 10.1016/j.cell.2010.01.004)
Annexes
Bibliographie
- Cooper, Geoffrey, La Cellule, une approche moléculaire, 3e édition, 1997, Éd. De Boeck Université.
- B. M. Alberts, « Prokaryotic DNA replication mechanisms », Philos. Trans. R. Soc. Lond., B., Biol. Sci., vol. 317, no 1187, , p. 395-420 (PMID 2894677, lire en ligne [[PDF]]).
Articles connexes
Liens externes
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire