Répartition cosmopolite
Une répartition cosmopolite en biogéographie, caractérise l'aire de répartition géographique d'une catégorie d'êtres vivants si étendue qu'elle peut être rencontrée dans toutes les régions du monde. Le qualificatif s'applique également à certaines maladies. S'opposant au concept d'endémisme, ce type de répartition est parfois désigné sous le nom de pandémisme[1].
Application et limites
Catégories impliquées
Ce type de répartition concerne essentiellement des taxons, quel que soit leur rang taxonomique (espèce, genre, famille, etc.). C'est aussi le cas de certaines maladies, mais comme il s'agit dans la majorité des cas de maladies infectieuses liées à des micro-organismes ou d'autres parasites, elles ne constituent le plus souvent qu'un cas particulier du précédent.



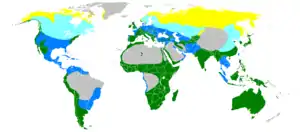
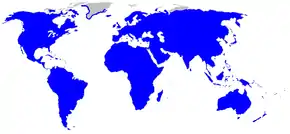 Une famille : les Rosaceae…
Une famille : les Rosaceae… un genre : les véroniques…
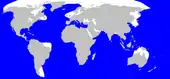
un genre : les véroniques…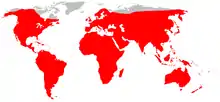 une espèce : le surmulot…
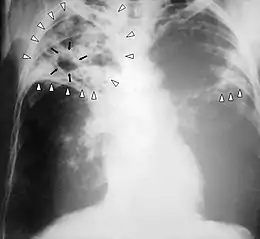
une espèce : le surmulot…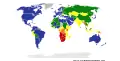 une maladie : la tuberculose en 2006.
une maladie : la tuberculose en 2006.
Répartition cosmopolite et milieux de vie
Contrairement à ce que pourraient laisser entendre certaines définitions du terme qui associent ce type de répartition à l'occupation d'un nombre minimum de continents, il existe des organismes cosmopolites non seulement dans les habitats continentaux (terrestres ou aquatiques) mais aussi dans les habitats marins.
Les cartes de répartition d'espèces ou d'autres taxons pourtant qualifiés de cosmopolites montrent que ces organismes n'occupent pas la totalité de la planète et l'espace qu'ils investissent réellement subit certaines restrictions. La première limitation est précisément liée à l'habitat, aquatique ou terrestre : la répartition des espèces marines est à l'évidence complémentaire de celle des espèces continentales.
Les contraintes climatiques comptent parmi les facteurs écologiques imposant les restrictions les plus évidentes aux aires de répartition : elles en réduisent les amplitudes latitudinale et altitudinale. Ainsi, les régions polaires, Arctique et continent Antarctique, ne sont habitées que par un très petit nombre d'organismes, et de très nombreuses espèces qualifiées de cosmopolites n'y pénètrent pas. De même, les espèces à vaste répartition sont très rarement présentes dans les déserts.
En fait, la somme des exigences écologiques spécifiques peut aboutir à des distributions paradoxales en apparence : certains organismes peuvent avoir une distribution à la fois cosmopolite et très localisée. L'exemple des organismes côtiers est particulièrement parlant à cet égard : ceux d'entre eux dont la distribution est considérée comme cosmopolite n'occupent en réalité qu'une étroite bande, linéaire, continue ou discontinue, sur les littoraux du monde entier.
La définition d'une répartition cosmopolite sous-entend donc ces limitations : un organisme cosmopolite est bien présent dans toutes les régions du monde, mais seulement là où ses exigences en matière d'habitat sont respectées. C'est sans doute parce qu'il s'agit là d'une tautologie que cette précision est si peu exprimée dans les définitions du terme.
Répartition subcosmopolite
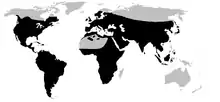
Pour des raisons historiques ou écologiques, de nombreuses catégories à très vaste répartition géographique manquent totalement dans telle ou telle partie du monde. Les crapauds vrais (Bufonidae), par exemple, outre les régions polaires et les déserts évoqués précédemment, sont absents de la zone australasienne, au sud de la ligne Wallace, et de Madagascar. Selon les auteurs, cette famille est donc considérée comme cosmopolite ou subcosmopolite (synonyme : pandémique), selon que l'accent est mis sur la très vaste répartition ou sur le caractère incomplet de cette répartition[2]. Bon nombre d'espèces catégorisées comme cosmopolites correspondent plutôt à cette catégorie.
Origine
À partir de sa région de naissance, à la mesure de ses capacités intrinsèques et des barrières écologiques que lui oppose ou non l'environnement, une espèce peut étendre son domaine et, éventuellement, devenir cosmopolite.

L'aptitude intrinsèque d'un organisme à l'expansion tient à diverses caractéristiques spécifiques : profil démographique (capacité de multiplication), moyens de dispersion permettant de parcourir de longues distances ou de franchir les barrières naturelles à un stade quelconque du cycle de vie, disposition à occuper une gamme plus ou moins diversifiée d'habitats, aptitude compétitive, etc.
Quant aux barrières naturelles, elles peuvent être de nature géographique ou écologique : océans et autres étendues d'eau pour les espèces continentales, continents pour les organismes marins, chaînes de montagnes, déserts, régions dont les conditions écologiques sont défavorables, présence d'espèces concurrentes, etc.
Mais une répartition observée, qu'elle soit contemporaine ou paléobiogéographique, est aussi la résultante d'une histoire. Au cours des temps géologiques, la répartition respective des masses continentales et océaniques et leur localisation géographique ont énormément varié (tectonique des plaques), de même que la disposition des barrières naturelles et la distribution des conditions écologiques. Des aires jadis cosmopolites plus ou moins continues ont pu se dissocier entraînant le plus souvent des processus de spéciation (hypothèse de la vicariance). L'histoire géologique de la planète et les mécanismes de l'évolution on en fait tendance à morceler les aires de répartition de grande étendue : à partir d'une espèce cosmopolite ou subcosmopolite, elles favorisent la naissance de plusieurs espèces à répartition plus restreinte.
- origine
- invasif
- potentiel
- localité isolée
Ainsi, les biogéographes recherchent-ils souvent dans les périodes où les masses continentales et océaniques étaient réunies, la Pangée[3] — pour les organismes terrestres — et la Panthalassa[4] ou la Téthys[5] — pour les taxons marins —, l'origine de la répartition cosmopolite actuelle ou ancienne de certaines catégories[1], surtout de rang taxonomique élevé (genres, familles, classes, phylums). À titre d'exemple, il existe des éléments pour affirmer qu'il n'y avait pas de restriction à la distribution de la plupart des tétrapodes sur la Pangée au Permien[6].
De nos jours, l'homme — espèce elle-même cosmopolite — contribue grandement, par ses introductions volontaires ou accidentelles, à l'expansion mondiale d'espèces à l'origine localisées. Ce phénomène pallie le défaut de capacités intrinsèques de dispersion à longue distance. Le nombre de ces espèces introduites devenues cosmopolites ou en passe de l'être, et parfois invasives, est considérable. Il s'agit tant de végétaux que d'animaux, d'espèces libres que de parasites, d'organismes terrestres que d'espèces aquatiques.
Le problème des espèces cryptiques
Cas général

Dans de nombreux cas, le constat de distributions à l'échelle mondiale pose toutefois de sérieux problèmes. D'une part la réduction, pour ne pas dire l'absence, des flux de gènes entre populations éloignées, d'autre part les nécessaires différences écologiques entre habitats homologues situés sous des climats différents, ne peuvent que favoriser la spéciation. L'hypothèse doit donc être envisagée selon laquelle un certain nombre d'espèces dites cosmopolites constitueraient en fait des complexes d'espèces cryptiques, ensembles d'organismes à la morphologie identique ou presque (morpho-espèces), mais génétiquement distincts ou non interféconds, et à distribution beaucoup plus limitée.
L'avènement et le développement récent des techniques de phylogénie moléculaire a suscité un énorme regain d'intérêt pour ces questions. Cela a surtout permis de montrer que nombre d'espèces jusqu'alors considérées comme cosmopolites constituaient effectivement de tels complexes d'espèces cryptiques, et abouti à la description de nombreuses espèces nouvelles pour la science.
À titre d'exemple, Eurythoe complanata, un ver marin de la famille des amphinomidés, a été décrit par Pallas en 1766 à partir d'exemplaires provenant des Caraïbes. De cette date au XXe siècle, une vingtaine d'espèces proches seront à leur tour décrites, puis peu à peu rassemblées en une seule sur la base de leur grande homogénéité morphologique ; ainsi, dans la seconde moitié du xxe siècle, E. complanata était considérée comme une espèce cosmopolite ou subcosmopolite, essentiellement circumtropicale, présente dans les océans Pacifique, Atlantique et Indien, ainsi qu'en Méditerranée et en Mer Rouge. Des analyses de génétique moléculaire portant sur une partie seulement des populations « mondiales » (atlantiques et est-pacifiques) ont révélé l'existence d'au moins trois espèces[7].
Des résultats du même ordre se multiplient pour un grand nombre d'organismes réputés cosmopolites dans des groupes extrêmement divers : autres polychètes[8], éponges[9], champignons[10], mousses[11], sipunculiens[12], bryozoaires[13], poissons[14], etc.
« Everything is everywhere »
On peut faire remonter à C. G. Ehrenberg, C. Darwin ou M. W. Beijerinck les premières intuitions selon lesquelles les organismes de petite taille auraient le plus souvent de vastes répartitions contrairement à la majorité des espèces de grande taille[15]. Cette opinion a pratiquement eu valeur de paradigme tout au long du XXe siècle, avant d'être plus fermement théorisée et revendiquée comme une véritable hypothèse scientifique à partir de la fin des années 1990[16]. L'hypothèse est régulièrement évoquée — et aujourd'hui controversée — sous l'expression « Everything is everywhere, but the environment selects » (« Toute chose est partout, mais l'environnement sélectionne »)[note 1] due au microbiologiste néerlandais Lourens Baas Becking en 1934[17].
L'argumentation avancée en faveur de cette hypothèse peut être ainsi résumée[16],[18],[19] :
- le pouvoir de multiplication des organismes de petite taille (<1 mm) est énorme et leurs populations sont considérables ;
- leur petite taille les rend aisément transportables, passivement, sur de très longues distances (courants aériens[20] et marins) ; il n'existe donc pas, à la surface de la Terre, de véritable barrière naturelle à leur dissémination, ce qui réduit d'autant le risque d'interruption des flux génétiques et la possibilité de diversification génétique ;
- nombre d'entre eux développent des formes de résistance ; associée au fort pouvoir de multiplication, cette possibilité réduit fortement les probabilités d'extinction locale.
Dans la mesure où elle laisse peu de place à la spéciation, un corollaire de cette théorie est que la biodiversité des micro-organismes doit être très inférieure à celle des espèces de grande taille. En d'autres termes et d'une manière schématique, alors que la répartition des organismes de grande taille, très diversifiés, participerait plutôt de l'endémisme, les micro-organismes seraient peu divers à l'échelle mondiale, mais essentiellement cosmopolites[16].
Divers indices, de nature observationnelle et plus rarement expérimentale, vont dans le sens de cette thèse. Parmi ceux-ci, les mesures et les estimations récentes de l'importance du transport transcontinental et transocéanique de microbes dans l'atmosphère, à l'occasion notamment des tempêtes de poussière et de sable : spores de champignons, bactéries, virus, pollens… sont ainsi susceptibles d'être transportés sur des milliers de kilomètres, l'importance estimée du flux annuel dans l'atmosphère étant comprise entre 1018 et 1020 micro-organismes[20],[18]. Par ailleurs, le nombre actuel d'espèces de microbes procaryotes officiellement décrites est de l'ordre de 5 000[19], ce qui est effectivement très inférieur à la diversité des animaux ou des plantes, et conforme au constat de l'hypothèse cosmopolite.
Notes et références
Notes
Références
- (fr) Cecca, F., 2009. La dimension biogéographique de l’évolution de la Vie. Comptes Rendus Palevol, 8 (2-3), 119-132. DOI:Résumé Article (620 Ko).
- (en) Pramuk, J.B., Robertson, T., Sites Jr, J.W. & Noonan, B.P., 2008. Around the world in 10 million years: biogeography of the nearly cosmopolitan true toads (Anura: Bufonidae). Global Ecology and Biogeography, 17, 72–83. DOI:Résumé « Article (778Ko) »(Archive • Wikiwix • Archive.is • Google • Que faire ?).
- (en) Cover, M.R. & Resh, V.H., 2008. Global diversity of dobsonflies, fishflies, and alderflies (Megaloptera; Insecta) and spongillaflies, nevrorthids, and osmylids (Neuroptera; Insecta) in freshwater. Hydrobiologia, 595, 409–417. DOI:Résumé Article (276Ko).
- (en) Brayard, A., Buchera, H., Escarguel, G., Fluteau, F., Bourquin, S. & Galfetti, T., 2006. The Early Triassic ammonoid recovery: Paleoclimatic significance of diversity gradients. Palaeogeography, Palaeoclimatology, Palaeoecology, 239 (3-4), 374-395. DOI:Résumé.
- (en) Camacho, A.I., 2003. Historical biogeography of Hexabathynella , a cosmopolitan genus of groundwater Syncarida (Crustacea, Bathynellacea, Parabathynellidae). Biological Journal of the Linnean Society, 78 (4), 457–466. DOI:Résumé.
- (en) Sidor, C.A., O’Keefe, F.R., Damiani, R., Steyer, J.S., Smith,R.M.H., Larsson, H.C.E., Sereno, P.C., Ide, O. & Maga, A., 2005. Permian tetrapods from the Sahara show climate-controlled endemism in Pangaea. Nature, 434, 886-889. DOI:Résumé Article (350Ko).
- (en) Barroso, R., Klautau, M., Solé-Cava, A.M., & Paiva, P.C., 2010. Eurythoe complanata (Polychaeta: Amphinomidae), the ‘cosmopolitan’ fireworm, consists of at least three cryptic species. Marine Biology, 157 (1), 69-80. DOI:Résumé.
- (en) Bleidorn, C., Kruse, I., Albrecht, S., & Bartolomaeus, T., 2006. Mitochondrial sequence data expose the putative cosmopolitan polychaete Scoloplos armiger (Annelida, Orbiniidae) as a species complex. BMC Evolutionary Biology, 6, 47 DOI: Article en ligne.
- (en) Xavier, J.R., Rachello-Dolmen, P.G., Parra-Velandia, F., Schönberg, C.H.L., Breeuwer, J.A.J. & van Soest, R.W.M., 2010. Molecular evidence of cryptic speciation in the “cosmopolitan” excavating sponge Cliona celata (Porifera, Clionaidae). Molecular Phylogenetics and Evolution, 56 (1), 13-20. DOI:Résumé.
- (en) Pringle1, A., Baker, D.M., Platt, J.L., Wares, J.P., Latgé, J.P. & Taylor, J.W., 2005. Cryptic speciation in the cosmopolitan and clonal human pathogenic fungus Aspergillus fumigatus. Evolution, 59 (9), 1886–1899. DOI:Résumé Article (184Ko).
- (en) Fernandez, C.C., Shevock, J.R., Glazer, A.N. & Thompson, J.N., 2006. Cryptic species within the cosmopolitan desiccation-tolerant moss Grimmia laevigata. PNAS ͉ 103 (3) ͉ 637– 642. DOI:Résumé Article (416Ko).
- (en) Kawauchi, G.Y. & Giribet, G., 2010. Are there true cosmopolitan sipunculan worms? A genetic variation study within Phascolosoma perlucens (Sipuncula, Phascolosomatidae). Marine Biology, 157 (7), 1417–1431. DOI:Résumé.
- (en) Nikulina, E.A., Hanel, R. & Schafer, P., 2007. Cryptic speciation and paraphyly in the cosmopolitan bryozoan Electra pilosa—Impact of the Tethys closing on species evolution. Molecular Phylogenetics and Evolution, 45, 765–776. DOI:Résumé.
- (en) Castro, G.M., Heras, S., Cousseau, M.B. & Rolda, M.I., 2008. Assessing species validity of Mugil platanus Günther, 1880 in relation to Mugil cephalus Linnaeus, 1758 (Actinopterygii). Italian Journal of Zoology, 75 (3), 319-325. DOI:Résumé.
- (en) Finlay, B.J. & Fenchel, T., 2004. Cosmopolitan metapopulations of free-living microbial eukaryotes. Protist, 155 (2), 237-244. DOI:Résumé Article (1,5Mo).
- (en) Fenchel, T. & Finlay, B.J., 2004. The ubiquity of small species: Patterns of local and global diversity. BioScience, 54 (8), 777–784. Résumé.
- (en) Whitfield, J., 2005. Biogeography:Is Everything Everywhere?. Science, 310, n°5750, 960-961. DOI:Résumé Article (450Ko).
- (en) Logares, R.E., 2006. Does the global microbiota consist of a few cosmopolitan species? Ecología Austral, 16, 85-90. Article.
- (en) Papke, R.T. & Ward, D.M., 2004. The importance of physical isolation to microbial diversification. FEMS Microbiology Ecology, 48 (3), 293–303. DOI:Résumé Article (466Ko).
- (en) Kellogg, C.A. & Griffin, D.W., 2006. Aerobiology and the global transport of desert dust. Trends in Ecology and Evolution, 21 (11), 638-643. DOI:Résumé.