Prométaphase
La prométaphase est la phase qui se déroule après la prophase et avant la métaphase, de la mitose et de la méiose des cellules eucaryotes. De plus, elle ne se produit pas chez toutes les cellules eucaryotes.
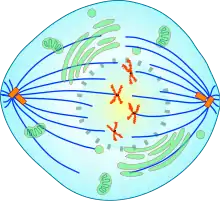
Certains auteurs considèrent la prométaphase comme une partie de la prophase plutôt qu'une phase distincte.
Elle débute avec la rupture de l’enveloppe nucléaire et se termine lorsque tous les chromosomes sont fixés sur le fuseau mitotique, juste avant la formation de la plaque équatoriale. Elle se caractérise donc par la capture des chromosomes par les fibres de ce fuseau.
Le rôle de la prométaphase s'achève quand tous les chromosomes sont correctement attachés (accrochage amphitélique). Si tel n'est pas le cas, le chromosome mal attaché envoie un signal inhibiteur et la cellule doit attendre que le problème soit résolu pour passer en métaphase. Ce phénomène de contrôle vise à minimiser les risques de mauvaise disjonction.
Notons que le terme "fibre" désigne un faisceau de plusieurs microtubules.
Mitose
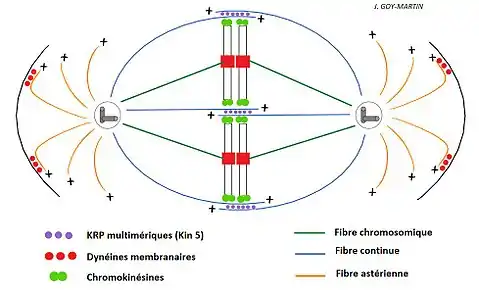
Mise en place de l’appareil mitotique achromatique
L'appareil achromatique se forme complètement sous le contrôle du complexe CDK1-Cycline B1, et est composé des deux pôles, des asters et du fuseau. Le fuseau va lui se décomposer en différentes fibres (microtubules) :
- Les fibres continues (ou polaires ou chevauchantes), liées à des Krp 5 multimériques (Kinésine 5), se chevauchent au niveau de leur extrémité + pour former des structures chevauchantes. Ces derniers, avec les moteurs protéiques Krp 5, permettent de repousser les centrosomes au deux extrémités de la cellule.
- Les fibres chromosomiques (ou kinétochoriennes) peuvent se lier aux kinétochores des chromosomes en condensation (elles représentent 80% des microtubules du fuseau). Elles permettent la traction des chromosomes vers le plan équatorial de cellule par des enchaînement de polymérisation/dépolymérisation. Chez les mammifères il y a entre 15 et 40 microtubules par kinétochore, dans l’espèce humaine on en retrouve en moyenne 20.
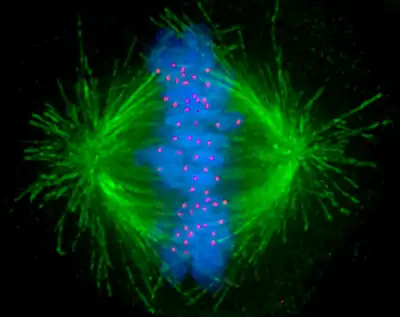
Ces différents microtubules, qui se sont développés à partir des centrosomes, envahissent progressivement l'espace laissé libre par la disparition de l'enveloppe nucléaire. Il y a un kinétochore pour chaque chromatide (donc deux par chromosome), localisé près du centromère.
Les asters sont des fibres qui ne participent pas au fuseau (dépend des auteurs). Les plus longues fibres astériennes se lient aux centrosomes et atteignent la membrane plasmique où des dynéines vont tracter les pôles (en complément des Kin5).
Devenir de l'enveloppe nucléaire
L’enveloppe nucléaire se disloque au début de la prométaphase et se trouve dispersée en petites saccules. On ne peut pas discerner (sauf les mettant en évidence) les saccules du réticulum endoplasmique de celle du noyau. Ces saccules nucléaire se répartissent à la périphérie et pourront rapidement être disponibles pour reformer l’enveloppe nucléaire à la fin de la télophase. La dissociation de l'enveloppe nucléaire est la conséquence de la phosphorylation des lamines B (présentent sur la membrane interne).
Les événements chromosomiques
Les chromosomes continuent leur condensation. Les 2 chromatides deviennent discernables mais restent étroitement accolées, grâce aux cohésines présentent tout le long du chromosome et au niveau du centromère. Les centromères eux apparaissent comme des "étranglements" des chromosomes (constriction primaire), chaque chromosome possède un centromère et deux kinétochores. Les kinétochores sont des structures complexes formées de nombreuses protéines, dont des moteurs pouvant se lier à aux microtubules kinétochoriens.
Les chromosomes vont être animés de mouvements grâce au fuseau mitotique. On retrouve des nombreuses structures qui vont moduler l'accrochage et le mouvement de ces chromosomes :
- Les fibres chromosomiques se fixent au niveau des kinétochores et tractent les chromosomes vers leur centrosome.
- Des protéines sont présentes au niveau du centromère, elles portent le nom de Complexes Passagers des Chromosomes (CPC) avec notamment la protéine Aurora B (anglais), qui peuvent décrocher les microtubules des kinétochores par phosphorylation et diminuer la force de traction polaire. Ce système intervient notamment dans le point de contrôle de la métaphase.
- Des kinésines particulières fixées sur les bras des chromosomes, les chromokinésines (ou kinésine 4-10), les tractent vers le centre en "marchant" sur les fibres continues d'un des pôles.
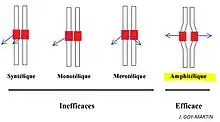
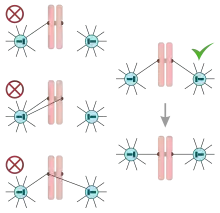
L'accrochage amphitélique des chromosomes
L'accroche amphitélique signifie que le centromère est relié, au niveau de ces kinétochores, par une fibre kinétochorienne (chromosomique) provenant des deux pôles de la cellule.
Cet accrochage amphitélique est assuré par le fuseau mitotique. Il va se mettre en place un phénomène de polymérisation/dépolymérisation des microtubules kinétochoriens (ou chromosomiques) appelé "fishing", dès la prométaphase jusqu'à la métaphase.
Si l'accrochage n'est pas amphitélique, la protéine Aurora B a pour rôle de décrocher les microtubules et ainsi de permettre aux chromokinésines, présentes au niveau des fibres continues (ou MT chevauchants) de ramener le chromosome au niveau de la plaque équatoriale.
Si l'accrochage est amphitélique, alors la protéine Aurora B devient inactive.
Méiose
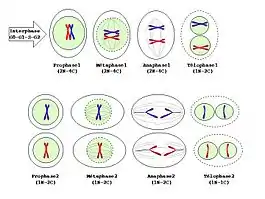
La prométaphase I de la méiose est sensiblement similaire à celle de la mitose. Dans le cas de la première division de méiose, il n'y a qu'un kinétochore par chromosome, sur le centromère toujours (de manière à séparer uniquement les chromosomes homologues); dans la deuxième division de méiose, le principe est le même que pour la mitose : un kinétochore par chromatide, de part et d'autre du centromère (de manière à séparer les chromatides).
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire