Orrorin tugenensis
Orrorin tugenensis est le nom donné à une espèce éteinte d'hominines âgée d'environ 5,9 millions d'années (Miocène supérieur), définie à partir d'un ensemble de fossiles dont certains ont été officiellement découverts en octobre et novembre 2000 dans la formation de Lukeino, dans les collines de Tugen, au Kenya, par Ezra Kiptalam Cheboi. Orrorin est le nom local d'un personnage mythique, « l'Homme originel ».
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Classe | Mammalia |
| Ordre | Primates |
| Famille | Hominidae |
| Sous-famille | Homininae |
| Tribu | Hominini |
| Sous-tribu | Hominina |
Orrorin tugenensis a également été surnommé Millennium Ancestor (Ancêtre du millénaire). Il est devenu momentanément le plus ancien hominine bipède, supplantant Ardipithecus ramidus (découvert en 1993, 4,4 Ma), lui-même suivi de près par Australopithecus anamensis (environ 4 Ma). Il a cependant été évincé dans ce rôle en 2002 par Toumaï (Sahelanthropus tchadensis), âgé de 6,9 à 7,2 millions d’années, mais en suscitant un désaccord chez plusieurs paléontologues en raison d'un fémur de conformation non-hominienne resté inédit[1].
Les fossiles
La première découverte, un fragment de mâchoire, fut identifiée par Brigitte Senut et Martin Pickford du Muséum national d'histoire naturelle de Paris, en collaboration avec les musées communautaires du Kenya[2]. Douze fragments de fossiles firent surface dans trois localités dans la formation de Lukeino, collines Tugen (montagne de district de Baringo au Kenya), en octobre et ; par la suite, vint s'ajouter à la liste un treizième fossile trouvé en 1974 dans une localité proche, la formation géologique de Ngorora (Miocène moyen, 12,5 millions d’années). Le statut phylogénétique de ce dernier avait jusque-là été le centre de spéculations variées et non corroborées.
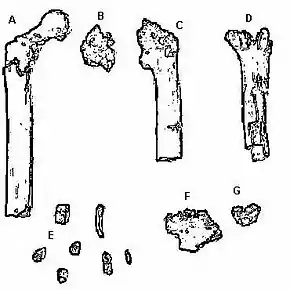
Orrorin tugenensis a donc été défini à partir de treize restes fossiles correspondant au moins à cinq individus distincts [3] et incluant :
- 1 humérus (bras) ;
- 3 fémurs, dont 1 morceau de petite taille (cuisse) ;
- 1 phalange (main) ;
- 2 mandibules ;
- 6 dents.
Découverte et « disparition »
À la suite d'une affaire de vols d'archives, le paléontologue Martin Pickford est en 1984 interdit de fouilles par les musées nationaux du Kenya, dirigé par son ancien coreligionnaire Richard Leakey, seule institution alors habilitée à délivrer des permis de fouilles dans ce pays. S'associant à Eustace Gitonga, ancien chef des expositions du Musée de Nairobi - licencié pour détournement de fonds-, Pickford obtient de celui-ci la possibilité de faire à nouveau des fouilles dans le pays : Gitonga a fondé une ONG (Community Museums of Kenya - CMK) ayant obtenu l'autorisation de délivrer des permis, et a confié à Pickford et sa compagne Senut, du Muséum d'histoire naturelle de Paris, la Kenya Paleontology Expedition (KPE), qui doit explorer les collines de Tugen, que Pickford a déjà fouillé dans les années 1970. Grâce au soutien d'Yves Coppens, co-découvreur de Lucy, sollicité par Pickford et Senut - une de ses thésardes -, le Muséum d'histoire naturelle et le Collège de France (dont Coppens est professeur émérite) financent la KPE[4].
Si la KPE a désormais un financement propre, son permis de fouilles est contesté par Richard Leakey, alors ministre de la fonction publique du Kenya ; en , Pickford est accusé d'avoir envahi un site de fouilles tenu par Meave Leakey - épouse de Richard - et arrêté pour « fouilles illégales et vol de fossile ». En , Pickford et Senut découvrent enfin le premier morceau d'Orrorin tugenensis, qu'ils dévoilent au public en décembre de la même année[4].
Les divers ossements découverts sont remis à l'ONG de Gitonga. Celui-ci les met en lieu sûr, dans un coffre de banque selon ses dires, et en limite peu à peu l'accès aux scientifiques auteurs de leurs découvertes. Souhaitant qu'un musée soit ouvert pour servir d'écrin au fossile, Gitonga obtient des fonds de l'ambassade de France et du Muséum d'histoire naturelle. Le musée est fondé à Kipsaraman, dans le massif de Tungen, mais Gitonga demande à nouveau des fonds pour créer un laboratoire pour étudier le fossile ; si le Muséum apporte en 2005 un financement pour lancer les travaux, le chantier pour le laboratoire ne sera jamais entamé, les fonds du Muséum ayant été redirigé « par les acteurs locaux à d’autres fins »[4].
Pickford et Senut n'ont pas revu les ossements depuis « 2002 ou 2003 », mais prêtent à la communauté scientifique les moulages qu'ils ont réalisés au début des années 2000 ; deux autres paléontologues, Brian Richmond et William Jungers, auteurs d’un article sur Orrorin publié dans le magazine Science en 2008, ont pu étudier les ossements dans leur coffre bancaire[4].
Description détaillée des fossiles
| Date | Localité | Code | Nature | |
|---|---|---|---|---|
| 1974 | Cheboit | KNM LU 335 | molaire M1 ou M2 gauche | Manque de l'émail sur les faces antérieure et linguale. Ligne occlusale rectangulaire, avec une légère élongation mésiodistale. Fovéa distale préservée sur cette dent seulement. |
| 2000 | Kapcheberek | BAR 349'00 | phalange - main | Courbée (pour grimper) |
| 2000 | Aragai | BAR 1215'00 | fémur proximal droit, petit fragment | Tête de fémur sphérique bien définie, qui se tourne légèrement vers l'avant.
Fovea capitis bien définie et moyennement large. Corps et col du fémur aplatis dans le sens antéro-postérieur. Petit trochanter (ou trochantin) large et médialement saillant. Sillon intertrochantérique allant d'une fosse trochantérique petite et moyennement profonde, jusqu'à un point juste au-dessus du petit trochanter. Angles col-corps estimés entre 120° et 130°. |
| 2000 | Kapsomin | BAR 1000a'00 | mandibule, fragment | Compose le holotype de O. tug. Porte toujours trois molaires inférieures, cassées et légèrement usées. |
| 2000 | Kapsomin | BAR 1002'00 | fémur proximal gauche | Le plus complet (tête fémorale et deux tiers du corps). Pas de grand trochanter. Ligne de fusion clairement apparente (jeune adulte). |
| 2000 | Kapsomin | BAR 1003'00 | fémur proximal gauche | Tête de fémur sphérique bien définie, qui se tourne légèrement vers l'avant (bipédie).
Fovea capitis bien définie et moyennement large. Corps et col du fémur aplatis dans le sens antéro-postérieur (grimper); Petit trochanter large et médialement saillant. Pas de grand trochanter. Sillon intertrochantérique allant d'une fosse trochantérique petite et moyennement profonde jusqu'à un point juste au-dessus du petit trochanter (bipédie). Angles col-corps estimés entre 120° et 130° (bipédie, contesté). Col du fémur long (bipédie). |
| 2000 | Kapsomin | BAR 1004'00 | humérus distal droit | |
| 2000 | Kapsomin | BAR 1001'00 | incisive supérieure I1 | Facette d'usure inclinée côté lingual.
Robuste, massive, mésiodistal assez large. Plus petite en taille que chez Australopithecus, et de même taille approximative que celles de Ardipithecus. Face labiale verticale et légèrement convexe dans le sens mésiodistal. Ligne cervicale de forme ovale presque circulaire. Surface linguale planaire fortement inclinée de l'apex vers le cervix. Absence de sillons linguaux clairement définis. |
| 2000 | Kapsomin | BAR 1425'00 | canine supérieure droite | Triangulaire en vue labiale, ligne cervicale peu compressée mésiodistalement et présentant un renflement, mais pas un vrai cingulum, au-dessus du cervix. Pas de crêtes linguales. Sillon mésial à peine marqué, vertical, peu profond et étroit. L'apex de la canine est pointu et presque sectoriel. |
| 2000 | Kapsomin | BAR 1390'00 | prémolaire P4 | Ligne occlusale ovoïde, comprimée (étroite) mésiodistalement, avec deux racines excentrées.
Manque de l'émail sur les faces antérieure et linguale. Épaisseur de l'émail de 3,1 mm à l'apex du paraconide. Fovéa distale large. Protoconide relevé au-dessus du métaconide. Deux crêtes partent de l'apex pointu du protoconide, côtés lingual et distal. |
| 2000 | Kapsomin | BAR 1426'00 | molaire M3 supérieure gauche | Manque de l'émail sur les faces antérieure et linguale.
Épaisseur de l'émail de 3,1 mm à l'apex du paraconide. Fovéa distale placée distobuccalement. Surface linguale verticale sans cingulum. Cuspides bunodont basses. Surface occlusale modérément plissée. Ligne occlusale trapézoïdale, presque triangulaire – métacone petit, gros protocone . Usure modérée à avancée (détails de la couronne sont imprécis). Couronne large, avec fovéa réduite. Racine assez longue. |
| 2000 | Kapsomin | BAR 1900'00 | molaire M3 supérieure droite | Manque de l'émail sur la surface mésiale, et un petit morceau d'émail sur la face distale.
Épaisseur de l'émail de 3,1 mm à l'apex du paraconide. Fovéa distale placée distobuccalement. Surface linguale verticale sans cingulum. Cuspides bunodont basses. Cuspides linguales pointues plus hautes que les cuspides buccales. Surface occlusale modérément plissée. Ligne occlusale trapézoïdale, presque triangulaire – métacone petit, gros protocone. Usure modérée à avancée (détails de la couronne sont imprécis). Couronne large, avec fovéa réduite. Racine assez longue. |
La denture présente « des affinités morphologiques et métriques » avec les deux anthropoïdes actuels. La molaire inférieure droite trouvée à Ngorora (12,5 M années) ressemble à celles du chimpanzé. Les molaires rappellent celles des hominidés plus tardifs, comme les australopithèques ou les représentants du genre Homo.
Taphonomie
La plupart des fossiles de Kapsomin sont recouverts d'une couche fine provenant d'algues et de bactéries indiquant un séjour dans l'eau avant que les corps ne soient enfouis sous les sédiments. Il y avait donc de l'eau, en quantité suffisante pour immerger tout un groupe, avant ou après sa mort.
D'autres, comme les deux fragments de mandibule, sont sévèrement craquelés ; ils ont été vraisemblablement desséchés par exposition aux éléments avant de se retrouver enterrés.
Datation
Les différents fossiles d’Orrorin n'ont pas fait l'objet de datations directes mais leur âge a pu être évalué grâce à celui de leur contexte stratigraphique, par biostratigraphie mais aussi par datations radiométriques et par magnétostratigraphie :
- le sill doléritique de Rormuch (5,62 ± 0,5 M années) passe à travers la Formation Lukeino ;
- les niveaux inférieurs des basaltes Kararaina (5,65 ± 0,13 millions d'années) recouvrent la Formation de Lukeino ;
- les niveaux supérieurs des trachytes de Kabarnet (6,2 ± 0,13 millions d'années) passent juste en dessous de la Formation de Lukeino ;
- des cristaux volcanogéniques provenant de la Formation de Lukeino ont été datés à 6,06 ± 0,13 millions d'années ;
- les fossiles d'autres espèces animales accompagnant Orrorin sont similaires à ceux trouvés dans des couches dont la datation ne fait aucun doute. L'âge commun indiqué est la fin du Miocène, 6 millions d'années.
Les fossiles trouvés sont donc âgés d'approximativement 6,2 à 5,65 millions d'années.
Plus récemment[5], des datations radiométriques (K-Ar et 40Ar-39Ar) croisées à des données magnétostratigraphiques indiquent que la formation de Lukeino, qui a livré les restes d'Orrorin tugenensis, s'est déposée entre 6 et 5,7 millions d'années. Les spécimens de Kapsomin, les plus nombreux et les plus importants de l'échantillon, seraient datés entre 5,9 et 5,8 millions d'années.
Principales caractéristiques
D'après la taille des os disponibles et surtout des fémurs, Orrorin pouvait mesurer 1,40 m pour un poids de 50 kg. On ne connaît pas le degré de variabilité du poids chez les membres du même genre et il est possible qu'elle soit importante - seuls d'autres fossiles peuvent nous renseigner sur ce point. En ne tenant compte que du matériel disponible, Orrorin aurait donc été plus grand que A.L. 288-1 (Lucy), ce qui en fait une exception parmi les hominidés primitifs.
Pickford et Senut estiment que la découverte d'Orrorin permet de prouver que celui-ci est, non seulement le plus vieux, mais également le seul ancêtre viable du genre Homo, les australopithèques n'étant alors plus qu'une branche latérale et éteinte. Ces conclusions, dont la communauté scientifique reconnaît alors le potentiel, ne sont toutefois pas consensuelles, compte tenu des fossiles obtenus, trop peu nombreux et trop fragmentaires. Les examens de Ardipithecus kadabba (découvert en 1997, daté de 5,5 millions d'années) et de Sahelanthropus tchadensis (découvert en 2001, daté de 7 millions d'années) ont émoussé les arguments concernant Orrorin, ces deux derniers fossiles ayant été considéré également comme des espèces à part entière (respectivement en 2004 et en 2002)[4].
Locomotion
La nouvelle donne qui finit par faire consensus est la morphologie d’Orrorin qui permet la bipédie occasionnelle[6]. Les caractères morphologiques observés sur les trois fémurs, autant externes (principalement : col fémoral allongé, aplati antéro-postérieurement, tubérosité glutéale bien développée qui fusionne avec la crête qui court le long de la diaphyse postérieurement et vers le bas, gouttière du muscle obturateur externe) qu'internes (répartition asymétrique du cortex du col du fémur : plus épais en partie inférieure qu'en partie supérieure, comme chez les Hominidés actuels et fossiles, alors que c'est le contraire pour les grands singes africains) indiquent qu'Orrorin était bipède fréquent et orthograde.
Ceci ne fait pas pour autant de lui un bipède permanent au sol, à la différence des hommes, ni exclusif puisque certains éléments fossilisés du squelette présentent des indices d'adaptation au grimper :
- la phalange (BAR 349'00) est recourbée de façon permanente ; les mains d’O. tugenensis étaient celles d'un hominidé arboricole ;
- la crête latérale de l'humérus distal (BAR 1004'00) est droite, comme chez les chimpanzés modernes ainsi que chez A. afarensis, et est liée à l'existence de muscles puissants : c'est sur cette crête que s'attache le muscle m. brachioradialis.
Le membre inférieur présente donc des caractères propres à un équilibre bipède de grand anthropoïde, sans être pour autant permanent au sol, tandis que le membre supérieur présente des indices nets d'arboricolisme. Cette mosaïque de caractères témoignant de différents modes de locomotion se retrouve plus tard chez les homininés comme des australopithèques qui ont acquis cependant un équilibre bipède permanent.
Selon l'article de Richmond et Jungers de la revue Science du : « L'inclinaison de la tête du fémur et la présence d'un tubercule visible en vue antérieure sur Orrorin et sur l'homme moderne alors qu'il est caché derrière chez les australopithèques confirme que la démarche d'Orrorin s'approchait plus de celle de l'homme moderne que de celle de l'australopithèque. Sur ce tubercule (le petit trochanter) s'attache en effet le muscle qui permet de lever la jambe devant le corps. »
Ce rapprochement avec l'australopithèque tend à relativiser le caractère exceptionnel de la découverte. Toutefois, Sergio Almécija, chercheur au musée américain d'histoire naturelle, corédige un article dans la revue Nature en 2013, rapprochant le fossile des grands singes du Miocène[4].
Environnement et alimentation
On a retrouvé principalement des fossiles de bovidés de petite et moyenne taille, des colobes. Des impalas indiquent des forêts clairsemées ; les colobes des espaces de forêt dense.
Orrorin vivait probablement parmi les arbres en forêt clairsemée ou en bordure de forêt humide. Son alimentation était celle d'un omnivore : fruits, amandes de noyaux, graines, baies, insectes, et occasionnellement viande.
Aucune trace d'outils n'a été trouvée près des fossiles d'Orrorin.
Références
- (en) Ewen Callaway, « Controversial femur could belong to ancient human relative », Nature, vol. 553, , p. 391–392 (DOI 10.1038/d41586-018-00972-z, lire en ligne, consulté le )
- (en) B. Senut, M. Pickford, D. Gommery, P. Mein, K. Cheboi et Y. Coppens, « First hominid from the Miocene (Lukeino Formation, Kenya) », Comptes Rendus de l'Académie de Sciences, vol. 332, pp. 137-144, 2001.
- Photos de l'ensemble des fossiles d'Orrorin tugenensis
- Bruno Meyerfeld, « Mais où est donc passé « Orrorin », ce fossile disparu ? », Le Monde, (lire en ligne)
- Y. Sawada et al., 2002. "The age of Orrorin tugenensis, an early hominid from the Tugen Hills, Kenya", C. R. Paleovol 1, 293-303
- Pickford M., Senut B., Gommery D., Treil J., 2002. « Bipedalism in Orrorin tugenensis revealed by its femora », C. R. Palevol, 1, Académie des sciences, Éditions scientifiques et médicales, pp. 191-203
Annexes
Articles connexes
Liens externes
- Orrorin tugenensis, Brigitte Senut, CNRS
- « Les grands singes fossiles africains sortent de l’ombre », Intérêt- Général.Info, AFP, , Sciences historiques.
- « Séparation entre hominidés et grands singes », Terra Nova
- (en) « Search for the First Human – A Secrets of the Dead Special » (À la recherche du premier humain), Tim Roufs, University of Minnesota Duluth, section Anthropology, Prehistoric Cultures.
- (en) Bones of contention, (expression idiomatique vaguement traduisible par « Os à problème »), Channel 4, History, version abrégée et révisée d'un article par Matthew Reynolds en 2000 pour Trench One, le magazine de Time Team Club.
- (en) « Case for ancient upright-standing ancestor gets legs », (Le vieil ancêtre bipède prend de la vitesse) Kate Wong, Scientific American.com, Science and Technologies, article.
- (en) Modern Human Origins (Origines de l'humain moderne), Orrorin tugenensis.
- (en) Orrorin Tugenensis: Pushing back the hominin line
- Mais où est donc passé Orrorin, ce fossile disparu ?
 Portail de la Préhistoire
Portail de la Préhistoire  Portail de la paléontologie
Portail de la paléontologie  Portail des primates
Portail des primates  Portail du Kenya
Portail du Kenya