Métaécosystème
En écologie, le concept de métaécosystème désigne un ensemble d’écosystèmes reliés entre eux par des flux spatiaux d’énergie, de matière, et d’organismes. Il permet de comprendre la dynamique et le fonctionnement des écosystèmes, de l’échelle locale à l’échelle globale, en associant des notions d’écologie des communautés et d'écologie du paysage[1].
Naissance du concept
Au cours des dernières décennies, la recherche en écologie s’est caractérisée par un intérêt grandissant pour les processus agissant à grande échelle spatiale, au-delà de l’échelle locale[1]. En effet, les systèmes écologiques étant ouverts et interconnectés par des flux variés, il est rare que les processus écologiques soient purement locaux[2].
C’est ainsi que sont apparus les concepts de métapopulation et de métacommunauté, formalisés respectivement par Richard Levins en 1969, et David Sloan Wilson en 1992[3]. Le concept de métacommunauté a permis de mieux comprendre le fonctionnement et la dynamique des écosystèmes à grande échelle[4]. Toutefois, il n’inclut que les compartiments biotiques de l’environnement (différentes espèces interagissant les unes avec les autres), alors que beaucoup de propriétés des communautés ne peuvent être expliquées qu’en tenant compte des contraintes abiotiques de l’environnement[1],[4].
En 1935, lorsque Arthur George Tansley définit la notion d’écosystème, il souhaite apporter une vision holistique de la nature, dans laquelle les organismes sont en relation avec leur environnement abiotique et non pas indépendants de celui-ci[4]. Malgré cela, l’écologie des écosystèmes et l’écologie des communautés ont évolué comme deux disciplines distinctes[1],[4]. Ce n’est que récemment qu’a émergé en écologie un nouveau paradigme, désigné par Naeem et al., en 2002, sous le nom de « Biodiversity–Ecosystem Function Paradigm » (BEFP), visant à rassembler ces deux disciplines[4]. Son émergence aurait été motivée par la volonté de mieux comprendre et prédire les conséquences sur les écosystèmes de la perte de biodiversité et de la perturbation des cycles biogéochimiques dues aux activités humaines[4].
C’est dans ce contexte qu’en 2003, Loreau et al. proposent le concept de métaécosystème. Il s’agit d’une extension du concept de métacommunauté, auquel est ajouté l’effet des flux de ressources[1],[5]. L’objectif de Loreau et al. était alors de développer « un cadre théorique pour une écologie spatiale des écosystèmes qui intègre les perspectives de l’écologie des communautés et de l’écologie du paysage »[1].
Intérêts de l’approche métaécosystémique
L’étude des métaécosystèmes présente un réel intérêt en gestion et conservation de la nature[5],[3]. Les écosystèmes sont aujourd’hui soumis à des pressions d’origine anthropique de plus en plus fortes : changements climatiques, dégradation et fragmentation des habitats, surexploitation d’espèces, dispersion d’espèces envahissantes[3]. Pour prédire et gérer au mieux les conséquences de ces perturbations, il est important d’étudier les écosystèmes de façon intégrative, et de comprendre les potentielles rétroactions entre flux de ressources, communautés végétales et animales, et cycles biogéochimiques[5].
L’approche métaécosystémique pourrait ainsi permettre de mieux comprendre les changements de distribution d’espèces en réponse aux changements climatiques et à la modification des habitats[3]. D’autre part, l’étude de la connectivité entre écosystèmes présente un intérêt pour prédire et gérer la dispersion d’espèces envahissantes et la diffusion de contaminants dans l’environnement[3].
Théorie des métaécosystèmes
Structure spatiale
Les caractéristiques biotiques et abiotiques de l’environnement varient dans l’espace. Cette hétérogénéité influence la distribution spatiale des organismes et des ressources créant ainsi dans l’environnement une mosaïque d’écosystèmes différents[3],[6]. Un métaécosystème est donc constitué d’un ensemble d’habitats, formant des patchs, reliés entre eux par des flux d’organismes et de ressources, et entourés d’une matrice d’autres habitats avec lesquels ils n'interagissent pas ou peu[3],[5].
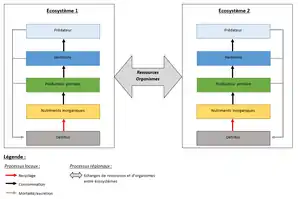
Processus locaux et régionaux
La dynamique des métaécosystèmes dépend de processus locaux et régionaux et de la façon dont ces derniers interagissent entre eux[6],[3]. Les processus locaux correspondent aux interactions trophiques (autotrophie, herbivorie, prédation) et non-trophiques (recyclage des nutriments), qui ont lieu au sein même des écosystèmes. Les processus régionaux, quant à eux, sont les flux d’organismes et de ressources reliant les différents écosystèmes entre eux[6].
Types de flux
Un métaécosystème est structuré par un ensemble de flux. Leur perturbation peut avoir des répercussions positives ou négatives, plus ou moins fortes, sur les différents compartiments qui le composent[7] et sur des écosystèmes éloignés avec lesquels ils sont connectés[5],[7].
Flux de ressources et d'organismes
Au sein des métaécosystèmes sont observables deux grandes catégories de flux : la dispersion d’organismes, et les flux de ressources[7].
Les flux de ressources regroupent les flux de nutriments inorganiques et les flux de détritus (nécromasse et produits de l'excrétion des êtres vivants). Ils peuvent être générés par des mécanismes physiques passifs (vent, gravité, courants…) entre écosystèmes similaires ou différents, proches ou éloignés[7],[5]. Par exemple, un flux de nutriments d’origine détritique est créé lorsque la litière d’origine terrestre tombe dans une rivière où elle sera décomposée par des organismes détritivores aquatiques[7]. Des flux de ressources peuvent également être générés de façon indirecte par certains types de déplacements d’organismes. C’est le cas des migrations saisonnières (cf. exemple 1), des migrations liées au cycle de vie (cf. exemple 2), et des déplacements liés à l’acquisition de nourriture (cf. exemple 3)[5].
- Exemple 1 : les troupeaux d’herbivores migrant à travers la savane déplacent les ressources qu’ils consomment en les excrétant le long de leur route de migration.
- Exemple 2 : lorsque les poissons anadromes retournent vivre dans les rivières à la fin de leur cycle de vie, et qu’ils y meurent, la carcasse qu’ils laissent derrière eux constitue un apport de ressources dans le compartiment détritique.
- Exemple 3 : les oiseaux de mer effectuant chaque jour des aller-retours entre la mer et le continent génèrent des flux de nutriments de la mer, où ils puisent leur nourriture, vers le continent, où ils défèquent et alimentent le compartiment détritique.
À la différence des flux de ressources, la dispersion a nécessairement lieu entre deux écosystèmes relativement similaires, donc éloignés. En effet, il y a dispersion lorsqu’un individu quitte son lieu de naissance, colonise un nouveau territoire, et s’y reproduit. Par conséquent, l’individu doit souvent se déplacer sur de longues distances pour trouver un habitat dont les caractéristiques sont similaires à celles de son habitat d’origine. De plus, contrairement aux autres types de déplacements d’organismes cités précédemment, la dispersion ne génère qu’un flux négligeable de ressources entre l’écosystème de départ et celui d’arrivée[5].
Flux d'informations
En plus des flux d’organismes et de ressources, Little et al. (2020)[2] proposent d’ajouter aux modèles de métaécosystèmes les flux d’informations. Ceux-ci peuvent influencer la dynamique des métaécosystèmes en modifiant le comportement, l’histoire de vie, et le développement des individus, ainsi que les mouvements d’individus et de matière entre écosystèmes.
Différents types d’informations pourraient ainsi être pris en compte dans les modèles de métaécosystèmes. Par exemple, l'information lumineuse peut être incluse dans l'étude de la dispersion des oiseaux migrateurs. En effet, "Les deux-tiers migrent de nuit" et leur sens de l'orientation est "perturbé par l'exposition à l'éclairage nocturne". De même, l'information sonore a un impact sur les flux de dispersion des organismes marins car dérangent leur capacités d'écholocation et doit être pris en compte dans les modèles de migration.
Modèles et dynamique
La prise en compte des ressources, par l’approche métaécosystémique, dans les modèles de dynamique des patchs ou de dynamique source-puits, modifie les prédictions établies à partir des modèles de métacommunauté et de métapopulation.
Modèle de patch-dynamique
Dans le cas du modèle de dynamique des patchs (où chaque patch matérialise un environnement pouvant contenir une communauté), Gravel et ses collaborateurs ont démontré que la direction du flux net de ressources pouvait favoriser ou défavoriser la coexistence de plusieurs espèces à l’échelle régionale[8]. Un flux net de ressources des patchs occupés vers les patchs vides favorise la coexistence par facilitation, exercée par les espèces top-compétitrices. À l’inverse, un flux net de ressources des patchs vides vers les patchs occupés entraîne à long terme l’installation d’une espèce meilleure compétitrice que les autres dans l’ensemble des patchs, par exclusion compétitive. Les flux de ressources pouvant donc modifier la structure des métacommunautés, il est nécessaire de les prendre en compte.
Dynamique source-puits
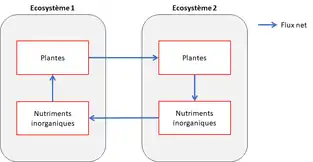
Le premier modèle de métaécosystème décrit par Loreau et ses collaborateurs[1] reprend un modèle classique en étude des métapopulations, le modèle de dynamique source-puits, en y intégrant les flux de ressources. Il considère un métaécosystème fermé et donne deux échelles de dynamique source-puits. Prenons l’échelle des échanges entre écosystèmes tout d’abord, avec des échanges entre compartiments identiques. Il dit alors que les compartiments au taux de croissance positif (sources) sont compensés par les compartiments au taux de croissance négatif (puits). À l’échelle d’un écosystème ensuite, les compartiments peuvent être des sources car à l’origine de flux sortants, ou bien des puits, car à l’origine de flux entrants. La combinaison de ces deux échelles de dynamique source-puits définit un sens de circulation des nutriments à l'échelle du méta-écosystème[1].
Plus tard, Gravel et ses collaborateurs ont également repris le modèle source-puits. Pour eux, une source est un patch dont la fertilité est plus grande que celle d'un patch puits. La fertilité est une propriété d’un patch isolé, qui indique une concentration de nutriments inorganiques présente dans un patch, en l’absence de consommateurs[9].
Ce modèle indique qu’il est fructueux que deux patchs, occupés par des producteurs primaires, interagissent. Pour cela il faut considérer deux patchs, l’un occupé par des producteurs primaires et l’autre vide. Les flux de matière inorganique (nutriments) et de matière organique (détritus) sont observés. Un patch occupé va recevoir les nutriments en excès de l’autre patch (puisque non consommés). Les flux de détritus quant à eux iront en direction du patch vide, où ils seront recyclés, lessivés et perdus. En revanche, si le patch qui reçoit les détritus est occupé par des producteurs primaires, les détritus seront recyclés et consommés et la perte de détritus est alors évitée[9].
Les modèles de Loreau et de Gravel suggèrent alors d’intégrer des notions d’écologie des écosystèmes au modèle de dynamique source-puits. En effet, un patch peut être un puits ou une source, selon le niveau de fertilité des patchs qui l’entourent. Cela pourrait conditionner la réponse à une perturbation ou à de la fragmentation[1],[9].
Structure spatiale et stabilité
La dynamique d'un métaécosystème est intimement liée à sa structure spatiale ainsi qu'à la connectivité entre les écosystèmes qui le composent[10]. La connectivité écologique est importante pour l’étude de la stabilité des écosystèmes. Cette notion peut être évaluée en ajoutant un facteur de dispersion d’organismes et de matière dans les modèles de métacommunauté.
C’est l’hétérogénéité environnementale qui va être le moteur de la stabilité du métaécosystème par la dynamique source-puits[11]. En cas de dispersion, la stabilité ne dépend plus du coefficient d’interaction de chaque écosystème isolé mais bien du coefficient d’interaction moyen sur l’ensemble des écosystèmes. Ainsi, la dispersion favorise l'homogénéisation des métaécosystèmes car elle redistribue la matière et les organismes, ayant pour effet de stabiliser la dynamique. À noter que lorsque les conditions environnementales des écosystèmes sont relativement similaires, l’effet de la dispersion est négligeable[10].
L'effet positif ou négatif de la dispersion sur la stabilité du système dépend de la nature du compartiment diffuseur : les flux de nutriments et de détritus ont tendance à déstabiliser tandis que les flux de producteurs primaires et de consommateurs n’ont pas d’effet, ou un effet stabilisant. Quand un seul flux est considéré à la fois, la stabilité du métaécosystème est plus importante quand les flux sont importants. Cependant lorsque la totalité des flux du métaécosystème est intégrée, la stabilité est plus importante quand chaque flux a une plus faible intensité[11].
L’étude de la connectivité des métaécosystèmes et des métacommunautés, qui peut empêcher ou faciliter la dispersion à l’échelle régionale, peut s’avérer primordiale dans la conservation et la gestion des espèces envahissantes par exemple[3].
Références
- (en) Michel Loreau, Nicolas Mouquet et Robert D. Holt, « Meta-ecosystems: a theoretical framework for a spatial ecosystem ecology », Ecology Letters, vol. 6, no 8, , p. 673–679 (ISSN 1461-023X et 1461-0248, DOI 10.1046/j.1461-0248.2003.00483.x, lire en ligne, consulté le )
- (en) Chelsea J. Little, Matteo Rizzuto, Thomas M. Luhring et Julia D. Monk, « Filling the Information Gap in Meta-Ecosystem Ecology », EcoEvoRxiv (prépublication), (DOI 10.32942/osf.io/hc83u, lire en ligne, consulté le )
- (en) Schiesari et al, « Towards an applied metaecology », Perspectives in Ecology and Conservation, vol. 17, no 4, , p. 172–181 (ISSN 2530-0644, DOI 10.1016/j.pecon.2019.11.001, lire en ligne, consulté le )
- (en) Shahid Naeem, « Ecosystem Consequences of Biodiversity Loss: The Evolution of a Paradigm », Ecology, vol. 83, no 6, , p. 1537–1552 (ISSN 1939-9170, DOI 10.1890/0012-9658(2002)083[1537:ECOBLT]2.0.CO;2, lire en ligne, consulté le )
- Isabelle Gounand, Eric Harvey, Chelsea J. Little et Florian Altermatt, « Meta-Ecosystems 2.0: Rooting the Theory into the Field », Trends in Ecology & Evolution, vol. 33, no 1, , p. 36–46 (ISSN 0169-5347, DOI 10.1016/j.tree.2017.10.006, lire en ligne, consulté le )
- Justin N. Marleau, Frédéric Guichard et Michel Loreau, « Meta-ecosystem dynamics and functioning on finite spatial networks », Proceedings of the Royal Society B: Biological Sciences, vol. 281, no 1777, , p. 20132094 (PMID 24403323, PMCID PMC3896007, DOI 10.1098/rspb.2013.2094, lire en ligne, consulté le )
- (en) Judith Sitters, Carla L. Atkinson, Nils Guelzow et Patrick Kelly, « Spatial stoichiometry: cross-ecosystem material flows and their impact on recipient ecosystems and organisms », Oikos, vol. 124, no 7, , p. 920–930 (DOI 10.1111/oik.02392, lire en ligne, consulté le )
- (en) Dominique Gravel, Nicolas Mouquet, Michel Loreau et Frédéric Guichard, « Patch Dynamics, Persistence, and Species Coexistence in Metaecosystems », The American Naturalist, vol. 176, no 3, , p. 289–302 (ISSN 0003-0147 et 1537-5323, DOI 10.1086/655426, lire en ligne, consulté le )
- (en) Dominique Gravel, Frédéric Guichard, Michel Loreau et Nicolas Mouquet, « Source and sink dynamics in meta-ecosystems », Ecology, vol. 91, no 7, , p. 2172–2184 (ISSN 1939-9170, DOI 10.1890/09-0843.1, lire en ligne, consulté le )
- (en) Dominique Gravel, François Massol et Mathew A. Leibold, « Stability and complexity in model meta-ecosystems », Nature Communications, vol. 7, no 1, , p. 12457 (ISSN 2041-1723, PMID 27555100, PMCID PMC4999499, DOI 10.1038/ncomms12457, lire en ligne, consulté le )
- (en) Isabelle Gounand, Nicolas Mouquet, Elsa Canard et Frédéric Guichard, « The Paradox of Enrichment in Metaecosystems », The American Naturalist, vol. 184, no 6, , p. 752–763 (ISSN 0003-0147 et 1537-5323, DOI 10.1086/678406, lire en ligne, consulté le )
Annexes
Articles connexes
 Portail de l’écologie
Portail de l’écologie