Homme de Denisova
L'Homme de Denisova, ou Dénisovien, est une espèce éteinte du genre Homo, identifiée par analyse génétique en à partir d'une phalange humaine fossile datée d'environ 41 000 ans, trouvée dans la grotte de Denisova, dans les montagnes de l'Altaï en Sibérie (Russie).

— auteur incomplet —, date à préciser

L'Homme de Denisova est parfois qualifié, en tant qu'espèce, du nom binominal Homo denisovensis[1], mais son statut d'espèce à part entière ou de sous-espèce n'est pas encore fixé. Jean-Jacques Hublin estime qu'il aurait vécu durant le Paléolithique moyen en Asie orientale, de la Sibérie à l'Asie du Sud-Est. La présence de cette espèce en Extrême-Orient est à rechercher selon lui parmi des fossiles connus[2]. Les analyses de l'ADN mitochondrial du fragment de phalange ont prouvé en 2010 que les Dénisoviens étaient génétiquement distincts des Néandertaliens et des Hommes modernes[3]. L'analyse ultérieure du génome nucléaire a montré que les Dénisoviens partageaient un ancêtre commun avec les Néandertaliens, et qu'ils se sont hybridés avec les ancêtres de certains hommes modernes (3 à 5 % de l'ADN des Mélanésiens et des Aborigènes d'Australie est issu des Dénisoviens)[4],[5],[6],[7]. De même, ils auraient transmis aux Tibétains un gène permettant leur adaptation à la vie en altitude[8].
Historique
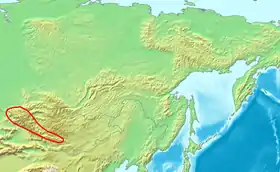

La grotte de Denisova se trouve dans le sud-ouest de la Sibérie, près de la frontière avec la Chine et la Mongolie. Elle fut nommée ainsi d'après un ermite russe, Denis, qui y vécut au XVIIIe siècle. La grotte fut explorée pour la première fois dans les années 1970 par le paléontologue russe Nikolaï Ovodov qui cherchait des fossiles de canidés[10]. En 2008, Mikhaïl Chounkov, de l'Académie des sciences de Russie, accompagné d'archéologues de l'Institut d'archéologie et d'ethnologie de Novossibirsk, explorèrent et fouillèrent la grotte. Ils trouvèrent la phalange d'auriculaire d'un humain juvénile de sexe féminin (connue comme la « femme X », en référence à la transmission matrilinéaire de l'ADNmt), puis ultérieurement un os d'orteil et deux dents dont une molaire[9]. Des objets (dont un bracelet) trouvés dans la grotte au même niveau que les fragments fossiles ont été datés par le carbone 14 entre 30 000 et 48 000 ans avant le présent[11]. Des fouilles plus récentes ont mis au jour des preuves d'une occupation intermittente remontant aux alentours de 125 000 ans[12].
Une équipe de scientifiques menée par Johannes Krause et Svante Pääbo, de l'Institut Max-Planck d'anthropologie évolutionniste à Leipzig en Allemagne, a séquencé en 2010 l'ADN mitochondrial extrait du fragment de phalange[3]. Le climat froid de la grotte a préservé l'ADN grâce à sa température moyenne annuelle de 0 °C. L'analyse génétique a révélé l'existence d'une espèce encore inconnue, l'Homme de Denisova[13].
En , une demi-mandibule humaine fossile, découverte en 1980 dans la grotte de Baishiya situé sur le plateau tibétain, près de Xiahe, dans le Gansu, en Chine, a été identifiée par une équipe internationale comme ayant appartenu à un Dénisovien. Faisant usage de techniques liées à une discipline émergente, la paléoprotéomique, cette étude s'est appuyée pour l'une des premières fois sur l'analyse des protéines conservées dans la mandibule de Xiahe, alors que l'ADN était trop dégradé pour être exploitable. La mandibule a conservé deux molaires, dont la morphologie est jugée comparable à celle des molaires trouvées dans la grotte de Denisova. Cette mandibule a un âge évalué par les séries de l'uranium à 160 000 ans[14], ce qui est quatre fois plus ancien que l'occupation humaine des hauts plateaux tibétains (par Homo sapiens) la plus ancienne connue auparavant[14],[15],[16].
Morphologie
Pendant presque dix ans, peu de choses ont été connues sur les caractéristiques morphologiques des Dénisoviens, puisque les seuls vestiges découverts étaient un os d'un doigt de la main, deux dents et un os d'orteil. L'os de doigt est exceptionnellement large et robuste, bien en dehors de la variation observée chez les humains modernes. Il appartenait à une femme, ce qui indique que les Dénisoviens étaient extrêmement robustes, peut-être même de la constitution de l'homme de Néandertal.
Une molaire exhumée dans la grotte, appartenant à un autre individu, est très grosse et archaïque d'aspect. Cette dent ne partage pas de caractéristiques morphologiques communes avec l'Homme de Néandertal ni avec l'Homme moderne[13].
Bien que découverte dans les années 1980, la demi-mandibule de Xiahe encore porteuse de deux dents a été identifiée comme dénisovienne en 2019 grâce à une analyse protéomique. Elle présente une morphologie archaïque, avec un corps très robuste, relativement bas et épais, dont la hauteur diminue légèrement vers l'arrière. Bien qu'une protubérance mentale triangulaire soit faiblement exprimée, il n'y a pas de menton développé et la symphyse est fortement en retrait[14].
Autres espèces
La stratigraphie de la grotte de Denisova montre une occupation ancienne de la grotte par des Dénisoviens et des Néandertaliens, et plus récente par des Homo sapiens. Néandertaliens et Dénisoviens semblent avoir partiellement cohabité, au moins à certaines époques, mais on ne sait pas à ce jour si Homo sapiens a fait de même ou s'il a seulement succédé aux deux premières espèces.
Phylogénie
Des chercheurs ont montré l'existence d'une forte diversité génétique chez les Dénisoviens[17]. L'analyse ADN de deux dents trouvées à un autre niveau que la phalange a révélé un degré inattendu de diversité de l'ADNmt des Dénisoviens[18].
L'analyse de l'ADN nucléaire de spécimens fossiles d'Homo sapiens, d'Homme de Néandertal et d'Homme de Denisova, menée en 2016 par une équipe de scientifiques coordonnée par Svante Pääbo de l'Institut Max-Planck d'anthropologie évolutionniste, à Leipzig, en Allemagne, a montré que l'Homme de Denisova et l'Homme de Néandertal avaient un ancêtre commun remontant à environ 450 000 ans, et que tous deux partageaient avec Homo sapiens un ancêtre commun remontant à environ 660 000 ans. Ces résultats s'appuient notamment sur l'analyse de deux spécimens fossiles de la Sima de los Huesos (Espagne), datés de 430 000 ans, attribués à l'Homme de Néandertal, et jugés postérieurs à la séparation d'avec l'Homme de Denisova[19]. Pour la première fois, les liens entre différents représentants du genre Homo ont pu ainsi être établis.
Une nouvelle étude publiée en 2020 précise que les chromosomes Y des hommes de Denisova se sont séparés il y a environ 700 000 ans d'une lignée partagée par les chromosomes Y humains néandertaliens et modernes, qui ont divergé les uns des autres autour de 370 000 ans. Les relations phylogénétiques des chromosomes Y archaïques et modernes humains diffèrent donc des relations de population déduites de leurs génomes autosomiques[20].

L'analyse comparée des génomes issus de la grotte de Denisova et des apports dénisoviens aux populations océaniennes actuelles, conduit à distinguer dénisoviens du nord (ceux de la grotte et les fossiles chinois) et du sud (actuellement absents du registre fossile)[21] :
| ancêtre commun (env. 650 ka) |
| ||||||||||||||||||
Hybridation
Denisova / Néandertal
Des analyses génétiques réalisées en 2013 comparant un Dénisovien à un Néandertalien découvert au même endroit ont révélé une hybridation locale ayant fourni 17 % du génome dénisovien, ainsi que des preuves d'une hybridation avec une autre souche archaïque du genre humain encore inconnue[18].
En 2018, l'ADN d'un fragment osseux trouvé dans la grotte de Denisova, celui d'une adolescente morte vers 13 ans il y a environ 90 000 ans[alpha 1], montre qu'elle était l'hybride d'une mère néandertalienne et d'un père dénisovien[22]. C'est la première preuve directe d'une hybridation de premier degré entre espèces humaines.
Denisova / Sapiens
D'autres analyses ont montré que des Homo sapiens et des Dénisoviens avaient eu des relations sexuelles interfécondes et avaient produit des hybrides dont les gènes se sont ensuite diffusés dans l'espèce voisine[23],[24]. L'Homme de Denisova aurait notamment contribué à hauteur de 3 à 6 % au génome des Papous et des aborigènes australiens actuels, et pour moins de 0,5 % au génome des populations d'Extrême-Orient. Les chercheurs en concluent que l'Homme de Denisova devait être relativement répandu en Asie orientale à la fin du Pléistocène[25], et que cette hybridation a dû faciliter l’adaptation des humains modernes à l'environnement des forêts équatoriales humides du sud-est asiatique[26].
Les Papous auraient acquis leurs 4 à 6 % d'ADN dénisovien de deux hybridations successives avec deux groupes distincts de Dénisoviens. La dernière hybridation aurait eu lieu il y a quelque 30 000 ans[26],[27]. Le principal flux de gènes lié aux Denisoviens s'est produit avant que les humains modernes n'atteignent Sahul, faisant de Wallacea et de Sunda des emplacements probables pour cet événement d'introgression archaïque[28].
Les Tibétains ont acquis des Dénisoviens (déjà présents sur le plateau tibétain il y a 160 000 ans) un allèle du gène EPAS1 facilitant leur adaptation à l'altitude[29].
Superarchaïques
Une étude génétique parue en février 2020, ayant analysé l'ADN fossile néandertalien (issu de spécimens de l'Altaï et de la grotte de Vindija, en Croatie) et l'ADN dénisovien (issu de la grotte de Denisova), en tire la conclusion que l'ancêtre commun de ces deux branches, appelé Néandersovien, aurait quitté l'Afrique il y a environ 750 000 ans et se serait peu après hybridé en Eurasie avec une population locale qualifiée de superarchaïque. Celle-ci serait sortie d'Afrique il y a environ 2 millions d'années, et serait donc très éloignée des Néandersoviens, ce qui n'aurait pas fait barrage à l'hybridation[30],[31].
Les Néandersoviens se seraient subdivisés entre Eurasiens de l'Ouest, ou Néandertaliens, et Eurasiens de l'Est, ou Dénisoviens, il y a environ 700 000 ans, c'est-à-dire beaucoup plus tôt que les estimations des études précédentes. Il y aurait eu par la suite une deuxième hybridation des superarchaïques avec les seuls Dénisoviens en Asie orientale[30].
Gènes dénisoviens sélectionnés chez Homo sapiens
Plusieurs études montrent qu'une partie du matériel génétique des Dénisoviens a été sélectionnée chez certaines populations d'Homo sapiens.
Le transfert de gènes dénisoviens vers les hommes modernes a laissé la plus forte fréquence d'une variante des gènes HLA (HLA-B) dans les populations d'Asie occidentale, endroit où des accouplements entre Homo sapiens et Dénisoviens se seraient produits.
En Papouasie-Nouvelle-Guinée, un gène dénisovien permettrait aux Papous de détecter des parfums très subtils.[réf. souhaitée]
Un gène dénisovien lié à l'hémoglobine permettrait aux populations himalayennes de vivre en altitude, où l'air est plus pauvre en oxygène[32]. Un variant du gène EPAS1 provenant des Dénisoviens améliore le transport d'oxygène et est présent seulement chez les Tibétains, et chez les Chinois Han dans une moindre proportion[33].
D'après une étude de l'université de Berkeley, les Inuits possèdent un variant très particulier du chromosome 1, portant deux gènes, TBX15 et WAR2. Le gène TBX15 joue un rôle dans le développement du corps et notamment dans celui du tissu adipeux brun, utilisé pour produire de la chaleur en cas de froid. À mesure que les latitudes augmentent, cette variante génomique apparait fréquente parmi les populations asiatiques, alors qu’elle est absente en Afrique, et rare en Europe. Ce gène ressemble bien plus à celui de l'homme de Denisova qu'à celui des autres populations humaines modernes. Les chercheurs émettent donc l'hypothèse que l'homme de Denisova aurait transmis une partie de son génome à Homo sapiens en Asie. Puis, au gré des migrations humaines, elle se serait répandue un peu partout sur la planète – cette séquence est retrouvée dans d'autres populations humaines, mais avec une fréquence bien moindre – et aurait connu une sélection naturelle particulièrement forte chez les Inuits[34],[35].
Comme presque toujours quand ils proviennent d'hybrides interspécifiques de mammifères, ces gènes ont été transmis par les hybrides femelles, les progénitures mâles étant le plus souvent infertiles. Suivant la règle de Haldane (en)[36], la stérilité hybride est limitée au sexe hétérogamétique. Les animaux hybrides homogamétiques (femelles chez les mammifères et les insectes, mâles chez les oiseaux et les lépidoptères[37]) sont normalement féconds.
Fossiles possiblement dénisoviens
En 2019, les seuls fossiles reconnus comme dénisoviens sont ceux de la grotte de Denisova et la mandibule de Xiahe. Mais d'autres fossiles découverts en Asie et décrits avant 2010, et considérés alors comme des Homo erectus tardifs ou des Homo sapiens archaïques (ou laissés en attente d'une dénomination), pourraient appartenir à l'Homme de Denisova, notamment :
- l'Homme de Jinniushan (Chine, Liaoning) :
- découverte : 1984 par Lu Zune,
- capacité crânienne estimée : 1 260–1 400 cm3,
- datation : entre −280 000 et −200 000 ans ;
- l'Homme de Dali (Chine, Shaanxi) :
- découverte : 1978 par Shuntang Liu,
- capacité crânienne estimée : 1 120 cm3,
- datation : −210 000 ans ;
- l'Homme de Maba (Chine, Guangdong) :
- découverte : 1958,
- capacité crânienne estimée : 1 300 cm3,
- datation : entre −150 000 et −130 000 ans.
Notes et références
Notes
- Il s'agit du fragment osseux dénommé Denisova-11. L'adolescente a été surnommée Denny.
Références
- Paolo Anagnostou et al., When Data Sharing Gets Close to 100% : What Human Paleogenetics Can Teach the Open Science Movement, 2015
- Jean-Jacques Hublin, Conférences du Collège de France, Néandertals et Dénisoviens, Dénisova, le groupe frère asiatique (3/6), novembre 2015
- Johannes Krause, Qiaomei Fu, Jeffrey M. Good, Bence Viola, Michael V. Shunkov, Anatoli P. Derevianko et Svante Pääbo, « The complete mitochondrial DNA genome of an unknown hominin from southern Siberia », Nature, 464(7290):894-7, mars 2010.
- (en-US) Carl Zimmer, « Denisovans Were Neanderthals’ Cousins, DNA Analysis Reveals », The New York Times, (lire en ligne, consulté le ).
- (en) Ewen Callaway, « First Aboriginal genome sequenced », Nature News, (DOI 10.1038/news.2011.551, lire en ligne, consulté le ).
- (en) Elizabeth Landau, « Oldest human DNA found in Spain - CNN », CNN, (lire en ligne, consulté le ).
- (en) Katherine Harmon, « New DNA Analysis Shows Ancient Humans Interbred with Denisovans », Scientific American, (lire en ligne, consulté le ).
- Pascal Depaepe, « Les Héritiéers d'Homo erectus », Sciences Humaines, , p. 44-47
- « L'homme d'Altaï, une nouvelle espèce d'hominidé ? », www.hominides.com, 24 mars 2010.
- (en) Nikolai D. Ovodov, Susan J. Crockford, Yaroslav V. Kuzmin et Thomas F. G. Higham, « A 33,000-Year-Old Incipient Dog from the Altai Mountains of Siberia: Evidence of the Earliest Domestication Disrupted by the Last Glacial Maximum », PLOS One, vol. 6, no 7, , e22821 (ISSN 1932-6203, DOI 10.1371/journal.pone.0022821, lire en ligne, consulté le ).
- avec AFP, « Un nouveau type d'hominidé découvert en Sibérie », Le Monde.fr, (lire en ligne).
- (en) Michael Marshall, « Mystery Relations », New Scientist, 5 avril 2014.
- D. Reich, R.E. Green, M. Kircher et al., « Genetic history of an archaic hominin group from Denisova Cave in Siberia », Nature, vol. 468, , p. 1053-1060 (lire en ligne)
- (en) Fahu Chen, Frido Welker, Chuan-Chou Shen et al., « A Late Middle Pleistocene Denisovan Mandible from the Tibetan Plateau », Nature, vol. 569, no 7754, (DOI 10.1038/s41586-019-1139-x, lire en ligne).
- (en-US) Carl Zimmer, « Denisovan Jawbone Discovered in a Cave in Tibet », The New York Times, (lire en ligne, consulté le )
- « L'Homme de Denisova vivait au Tibet il y a 160 000 ans, une surprise ! », sur Futura-Sciences, .
- M. Meyer, M. Kircher, M.-T. Gansauge et al., « A High-Coverage Genome Sequence from an Archaic Denisovan Individual », Science, (DOI 10.1126/science.1224344, résumé).
- (en) Elizabeth Pennisi, « More Genomes From Denisova Cave Show Mixing of Early Human Groups », Science, vol. 340, no 6134, , p. 799–799 (DOI 10.1126/science.340.6134.799, lire en ligne, consulté le ).
- (en) M. Meyer, J.-L. Arsuaga, C. de Filippo et al., « Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins », Nature, vol. 531, no 7595, , p. 504-7 (DOI 10.1038/nature17405, lire en ligne)
- (en) Martin Petr et al., « The evolutionary history of Neandertal and Denisovan Y chromosomes », bioRxiv, (DOI 10.1101/2020.03.09.983445, lire en ligne)
- Pour la Science, , p. 32.
- (en) Matthew Warren, « Mum’s a Neanderthal, Dad’s a Denisovan: First discovery of an ancient-human hybrid - Genetic analysis uncovers a direct descendant of two different groups of early humans », Nature, vol. 560, , p. 417-418 (DOI 10.1038/d41586-018-06004-0).
- Laurent Abi-Rached, M.J. Jobin, S. Kulkarni et al., « The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans », Science, vol. 334, no 6052, , p. 89-94 (DOI 10.1126/science.1209202, résumé).
- Carl Zimmer, « Toe Fossil Provides Complete Neanderthal Genome », The New York Times.com, (lire en ligne).
- La génétique arrache ses secrets de famille à un cousin des Néandertaliens.
- Pour la Science no 500 juin 2019 p. 14.
- Guy S. Jacobs et al., Multiple Deeply Divergent Denisovan Ancestries in Papuans, Cell, 2019 DOI:10.1016/j.cell.2019.02.035
- (en) Selina Carlhoff, Akin Duli, Kathrin Nägele et al., Genome of a middle Holocene hunter-gatherer from Wallacea, Nature volume 596, pages 543–547, 2021, doi.org/10.1038/s41586-021-03823-6
- (en) Chen F et al., A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau, Nature, 2019 DOI:10.1038/s41586-019-1139-x
- (en) Alan R. Rogers, Nathan S. Harris et Alan A. Achenbach, « Neanderthal-Denisovan ancestors interbred with a distantly related hominin », Science Advances, vol. 6, no 8, (DOI 10.1126/sciadv.aay5483, lire en ligne)
- « Les super-archaïques, une population fantôme d'hominidés il y a 700 000 ans », Hominidés.com, (lire en ligne)
- Lizzie Wade, « Our ancestors may have mated more than once with mysterious ancient humans », Science Mag.org, (DOI 10.1126/science.aaf9842, lire en ligne)
- E. Huerta-Sanchez, X. Jin, Asan, Z. Bianba et al., « Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA », Nature, vol. 512, , p. 194–197 (résumé).
- Science-et-vie.com, « Les Inuits auraient hérité leur résistance au froid de l'homme de Denisova », Science-et-vie.com, (lire en ligne, consulté le ).
- « Des gènes de l’homme de Denisova chez les Inuits », Sciences et Avenir, (lire en ligne, consulté le ).
- J. B. S. Haldane, « Sex ratio and unisexual sterility in hybrid animals », J. Genet., vol. 12, , p. 101–109 (DOI 10.1007/BF02983075)
- « Spéciation »(Archive • Wikiwix • Archive.is • Google • Que faire ?), sur univ-lille1 (consulté le )
Voir aussi
Bibliographie
- (en) David Reich et al., 2010, « Genetic history of an archaic hominin group from Denisova cave in Siberia », Nature, vol. 468, pp. 710-722.
- (en) H. Burbano et al., 2012, « Analysis of Human accelerated DNA regions », PLoS One, vol 7.(3), e32877.
- A. Degioanni et Silvana Condemi, « Néandertal, Sapiens et Denisova racontés par leurs gènes », Dossier Pour la Science no 76, juillet-, p. 20-25 (début de l'article en ligne).
- Jean-Jacques Hublin, « L'Homme de Denisova : Notre cousin d'Asie sort de l'ombre », Pour la science, no 506, , p. 28-36
Articles connexes
Liens externes
- « L’Homme de Denisova, entre Néandertal et Cro-Magnon ? », sur France Culture, (consulté le )
- Quand Homo sapiens peupla la planète (2/5), sur Arte (réalisateur Nigel Walk) diffusé mai 2020.
 Portail de la Préhistoire
Portail de la Préhistoire  Portail de la paléontologie
Portail de la paléontologie  Portail de l’Asie
Portail de l’Asie  Portail des primates
Portail des primates