Exaptation
Dans la théorie de l'évolution, l'exaptation est une adaptation sélective opportuniste, privilégiant des caractères qui sont utiles à une nouvelle fonction, pour laquelle ils n'avaient pas été initialement sélectionnés. Par exemple, les plumes des théropodes, initialement sélectionnées parce qu'elles assureraient leur thermorégulation, ont permis l'adaptation au vol. La bipédie ou la parole seraient également des exaptations[1].
L'origine du terme exaptation est attribuée aux biologistes Stephen J. Gould et Elisabeth Vrba (en), dans un article[2] expliquant comment des caractères ou des fonctions complexes peuvent apparaître et évoluer à partir de structures initiales simples.
L'exaptation est un concept qui étaye la critique de l'adaptationnisme. Il est l'une des dimensions de l'évolution qui intéresse la biologie de synthèse. Celle-ci voudrait s'en inspirer pour créer des gènes, des voies métaboliques ou des organismes nouveaux à partir de systèmes existants à modifier ou à détourner.
Confusion entre pré-adaptation, adaptation et exaptation
Entre 1901 et 1909, Lucien Cuénot élabore la théorie de la préadaptation qui rencontrera moins de succès que celle de l'exaptation « plus conforme aux standards scientifiques du XXe siècle », la formulation de Cuénot étant trop téléologique selon Gould et Vrba. Ces deux concepts se révèlent complémentaires : « la préadaptation concernerait le caractère avec sa fonction d'origine tandis que l'exaptation concernerait le caractère avec sa fonction dérivée[3] ».
Bien que les termes adaptation et exaptation soient souvent associés, ils ont des sens distincts :
- Une adaptation est le développement d’un caractère doté d'une fonction particulière qui permet à l’organisme le possédant d’améliorer sa valeur sélective (fitness).
- Une exaptation est le développement d’un caractère doté d'une fonction première qui, par la sélection naturelle, évolue au fil du temps pour jouer un tout nouveau rôle. Contrairement à l’adaptation, où la modification du phénotype (par exemple par le biais de mutations génétiques) est essentielle, l’exaptation n’amène aucun changement phénotypique.
Bien souvent, la littérature parle de l’exaptation comme d’un phénomène secondaire à l’adaptation. Toutefois, une adaptation peut également se produire à la suite d’une exaptation. On dit alors qu’un caractère subit une exaptation primaire, suivie d’une adaptation secondaire[4].
Types d'exaptation
On considère généralement deux grands types d'exaptation[4] :
- Dans le premier type, un organe non modifié est réutilisé à l'identique pour une fonction manifestement non sélectionnée à l'origine (coaptation ou exaptation par cooption). C'est ainsi le cas de certaines zones du cerveau humain ; utilisées initialement pour la reconnaissance des formes, le langage ou l'audition, elles sont aujourd'hui réutilisées pour la lecture ou la musique. C'est également probablement le cas de la bipédie[5].
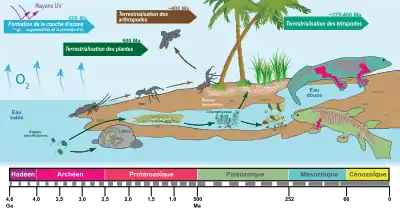
- Dans le second type, l'exaptation est une forme particulière d'adaptation naturelle, c'est-à-dire un ensemble de mutations génétiques sélectionnées par la sélection naturelle. Mais cette adaptation se fait sur la base d'un organe qui s'est initialement développé pour un tout autre usage. Le poumon des tétrapodes peut ainsi être considéré comme une exaptation : apparu chez des poissons comme une adaptation à des milieux aquatiques hypoxiques ou bien oxygénés (en lien dans ce dernier cas avec les besoins accrus en oxygène du cœur[6],[7]), il se serait secondairement modifié pour fonctionner en milieu purement terrestre.
Origines possibles de l'exaptation
Deux origines possibles de l’exaptation ont été mises de l’avant par les scientifiques Paul W. Andrews, Steven W. Gangestad et Dan Matthews[4] :
- La première possibilité suppose que le caractère issu de l’exaptation remplit en premier lieu une fonction qui se modifie au cours du temps. C’est le cas notamment de l’évolution fonctionnelle des plumes chez les oiseaux (voir exemple ci-bas). De nombreux scientifiques sont en effet d’avis que les plumes des oiseaux avaient pour fonction primaire la thermorégulation des individus (notamment grâce à la présence de barbes à la base des plumes). Ce n’est que plus tard dans l’évolution que les plumes auraient été utilisées pour le vol.
- La deuxième origine possible suppose que le caractère issu de l’exaptation soit une conséquence secondaire liée à la sélection d’un premier trait de caractère, ce dernier ayant un effet positif sur la fitness d’un individu. Au cours de l’évolution, ce caractère évolue pour acquérir de nouvelles fonctions et amener de nouveaux bénéfices. Ce type de genèse peut être illustré chez les escargots : la coquille de ces mollusques est formée de façon à y laisser un vide. Au cours de l’évolution, cet espace a graduellement été utilisé par certaines espèces pour le stockage des œufs lors de la reproduction. La modification de la fonction de cet espace illustre bel et bien un cas d’exaptation.
Quelques exemples d'exaptation
Les plumes chez les théropodes

Un exemple où l’exaptation prend tout son sens est le cas de l’apparition des plumes pour le vol chez les théropodes. Archeopteryx (première espèce de théropode à plumes découverte) a toujours été l’organisme clé permettant d’expliquer l’évolution des théropodes, notamment en raison des nombreux fossiles découverts et relativement bien conservés depuis le XIXe siècle. L'examen des dix fossiles récupérés a démontré que cette espèce avait une capacité de vol réduite en raison du diamètre plus étroit des rachis. Toutefois, un onzième spécimen découvert en Bavière en 2011[8] et spécialement bien conservé montre des plumes dotées de rachis solides. De plus, la distribution des plumes sur le dernier fossile laisse sous-entendre une grande ressemblance avec la distribution des plumes chez les oiseaux modernes. Ainsi, comment se fait-il que la morphologie alaire d’Archeopteryx soit similaire à celle des oiseaux modernes, alors même qu'Archeopteryx a une capacité de vol réduite ? Ceci laisse penser que les plumes pouvaient avoir une tout autre fonction que le vol au préalable. En effet, les théropodes auraient d’abord développé un plumage pour permettre la thermorégulation[9]. Ces plumes jouaient probablement aussi un rôle de camouflage et d'équilibrage lors des déplacements.
Les découvertes archéologiques montrent également un large éventail de coloration et de motifs pour les plumes. Ainsi, de nombreux chercheurs sont d’avis que celles-ci auraient pu servir initialement à la parade lors des périodes de reproduction. Elles auraient ensuite été réutilisées pour le vol, par sélection de leurs caractéristiques mécaniques[10].
Les choanes, les poumons et les pattes chez les tétrapodes
Chez les tétrapodes, les poumons (remplaçant les branchies) et les pattes (remplaçant les nageoires) sont apparus avant la sortie des eaux et la conquête de la terre ferme, c'est-à-dire que ces organes ont initialement été développés pour la vie en milieu aquatique :
- les choanes des poissons pulmonés et celles des tétrapodes sont apparues indépendamment par convergence évolutive résultant d'une adaptation à la vie terrestre. L'évolution phylogénétique suggère de considérer le nez respiratoire comme une exaptation du nez olfactif primaire[11].
- les poumons primitifs (sacs pulmonaires) des poissons pulmonés sont une adaptation première aux eaux pauvres des lagunes ou aux eaux bien oxygénés (en lien dans ce dernier cas avec les besoins accrus en oxygène du cœur). Ils ont été réutilisés lors des premières sorties de l'eau.
- les pattes (membres chiridiens) ayant un rôle de pagaies, avec des doigts pour agripper les proies lors d'une attaque. La fonction de marche des pattes permettant de sortir de l'eau et de conquérir la terre ferme est apparue plus tard ; il s'agit donc d'une exaptation d'une structure dérivée qui a gardé sa fonction primitive de marche et qui s'est diversifiée et perfectionnée au gré des lignées (notion de transaptation).
Les os
De plus, le développement des os est une caractéristique à la base du passage de la vie en milieu aquatique à la vie en milieu terrestre chez les vertébrés[2]. Pourquoi ces organismes possédaient-ils des os avant ce changement de mode de vie ? De nombreuses hypothèses ont été émises sur le sujet, bien qu’aucune n'ait été encore formellement prouvée. Certains scientifiques affirment que les os auraient d’abord joué un rôle dans l’osmorégulation des organismes aquatiques. D’autres adhèrent plutôt à la théorie que les os jouaient un rôle de protection contre les prédateurs, en agissant comme une armure. Finalement, certains chercheurs sont plutôt d’avis que les os avaient comme fonction première le stockage du phosphate permettant les activités musculaires. Cette théorie est particulièrement appuyée de nos jours, considérant que « chez les humains, 90 % du phosphore est présent dans la phase minérale de l’os ». Les os auraient donc rempli diverses fonctions telles que le stockage du phosphore et auraient évolué pour permettre aux organismes de se soutenir[2].
Le lait
Chez les mammifères, le lait est probablement produit par des glandes sudoripares modifiées (les glandes mammaires). Une sorte de sueur devait suinter sur le corps des synapsides (ancêtres des mammifères), qui avaient développé des œufs à coquille molle et perméable, à l'inverse des sauropsides dont les œufs sont calcifiés. À leur naissance, les jeunes se collaient à leur mère et étaient donc souvent en contact avec cette sécrétion aux propriétés protectrices et nutritives. La transformation de cette « sueur » ou sécrétion spéciale en lait a pu se faire par plusieurs exaptations successives. Aussi, le composé α-lactalbumine, grâce auquel la synthèse du lactose est possible, résulte d’une adaptation à la lactation. Des études ont démontré que la structure de cette protéine était génétiquement intimement liée à celle des lysosomes. Les lysosomes sont des organites cellulaires ayant pour fonction la lyse des cellules. Ils sont très utiles dans la défense des organismes, notamment en détruisant la paroi mucopolysaccharide des bactéries[2]. Selon certains chercheurs, la forte ressemblance des lysosomes avec α-lactalbumine laisserait supposer un nouveau cas d’exaptation.
Le clitoris de la hyène tachetée

Un autre exemple du concept d’exaptation, bien qu’il demeure encore aujourd’hui contesté scientifiquement, peut être observé chez les hyènes tachetées (Crocuta crocuta). Contrairement à la majorité des autres mammifères, les organes génitaux des femelles ne diffèrent pas d’un point de vue morphologique de ceux des mâles. Les femelles possèdent en effet un clitoris dont la taille est comparable avec celle du pénis du mâle. Les lèvres supérieures femelles peuvent aussi être facilement confondues avec le scrotum[12].
La proéminence des organes féminins est attribuée à un taux trop élevé des hormones androgènes dans le corps. Les hyènes femelles sont ainsi souvent qualifiées d’hermaphrodites et les scientifiques attribuent cette ressemblance génitale à une forme de mimétisme sexuel.
L’utilité de cette ressemblance est encore mal connue de la communauté scientifique. Selon les chercheurs, les femelles auraient au départ développé ce mimétisme pour la meeting ceremony. Bien que les hyènes aient un mode de vie majoritairement solitaire, ces animaux sont amenés à se regrouper en clans pour différentes raisons telles que la prédation. La meeting ceremony est une forme de rituel de reconnaissance effectué par les hyènes solitaires pour se regrouper, lors de laquelle chaque individu sent et lèche les parties génitales de ses confrères. Au cours de l’évolution, cette forme de mimétisme aurait évolué pour permettre aux femelles d'accéder à un statut social supérieur. Certains auteurs affirment en effet qu'une plus grande taille des organes génitaux est positivement corrélée avec l’acceptation par les pairs, ce qui peut également conférer un avantage en matière de fitness des individus[2].
Divers
L'exaptation s'oppose à la pensée d'Aristote selon laquelle la fonction crée l'organe[alpha 1]. Cette idée persiste encore aujourd'hui, bien qu'elle ait été réfutée dès le Ier siècle av. J.-C. par Lucrèce[14].
L'exaptation montre ainsi que l'évolution n'a pas de plan préétabli, mais qu'elle a plutôt tendance à « bricoler » avec l’existant. Il ne faut donc pas se poser la question du « pourquoi » mais plutôt celle du « comment ». La bipédie n'est pas apparue pour voir au-dessus des buissons, pas plus que l’œil n'est apparu pour voir[1] : « la fonction ne crée pas l'organe ». En revanche, ces aptitudes ont été développées parce qu'elles procuraient un avantage sélectif.
Notes et références
Notes
- L'expression « la fonction crée l'organe » n'est pas d'Aristote (elle a été employée par Darwin[réf. nécessaire], d'ailleurs pour la réfuter) mais l'idée, oui : « Chaque être, chaque organe a été créé par la Nature en vue d'une fin, d'une destination spéciale. » (dernière phrase de l'Histoire des animaux[13]).
Références
- Pascal Picq, La plus belle histoire du langage, Paris, Seuil, , 184 p. (ISBN 978-2-02-040667-3), p. 33-34.
- (en) Stephen Jay Gould, S.J.; Vrba, E. (1982) Exaptation - a missing term in the science of form, Paleobiology, vol. 8, p. 4-15.
- Thomas Heams, Philippe Huneman, Guillaume Lecointre et Marc Silberstein, Les mondes darwiniens: L'évolution de l'évolution, Éditions Matériologiques, (lire en ligne), p. 121
- Andrews, P.W.; Gangestad, S.W.; Matthews, D. (2002) Adaptationism – how to carry out an exaptationist program, Behavioral and brain sciences, vol. 25, p. 489-553.
- (en) Milford H. Wolpoff, Paleoanthropology, McGraw-Hill, , p. 219.
- (en) Colleen Farmer, « Did lungs and the intracardiac shunt evolve to oxygenate the heart in vertebrates », Paleobiology, vol. 23, no 3, , p. 358-372.
- (en) John N. Maina, The Biology of the Avian Respiratory System: Evolution, Development, Structure and Function, Springer, (lire en ligne), p. 102-103
- SCIENCE & VIE N°1164 - Septembre 2014
- (en) Ostrom, J.H. (1974) Archaeopteryx and the origin of flight, The quarterly review of biology, vol. 49, p. 27-47.
- (en) Foth, C.; Tischlinger, H.; Rauhut, O.W.M. (2014) New specimen of Archeopteryx provides insights into the evolution of pennaceous feathers, Nature, vol. 511, p. 79-82.
- (en) John Abramyan, Béatrice Thivichon-Prince, Joy Marion Richman, « Diversity in primary palate ontogeny of amniotes revealed with 3D imaging », Journal of Anatomy, vol. 226, no 5, , p. 420–433 (DOI 10.1111/joa.12291).
- (en) Harrison Matthews, L. (1939) Reproduction in the Spotted Hyaena, Crocuta crocuta (Erxleben), Philosophical transactions of the Royal Society of London, vol. 230, p. 1-78.
- Paul Rossi, « Promenades zoologiques dans la littérature ancienne », Bulletin de l'Association Guillaume Budé, vol. 1, no 3, , p. 269-282 (lire en ligne, consulté le ).
- De rerum natura, Livre IV, v. 834-835 : « Nil ideo quoniam naturum est in corpore ut uti / Possemus, sed quod natum est id procreat usum. » « Rien ne naît dans le corps pour qu'on puisse en user, / Mais ce qui naît d'abord crée ensuite l'usage. » Lire en ligne.
Voir aussi
Articles connexes
Liens externes
- (en) Exaptation-A missing term in the science of form, Stephen Jay Gould et Elisabeth Vrba (article de 1982).
- (en) Exaptation: How Evolution Uses What's Available, Wynne Parry, LiveScience, 16 septembre 2013.
- (en) Evolution as Opportunist, Wynne Parry, QuantaMagazine, 4 septembre 2013.
 Portail origine et évolution du vivant
Portail origine et évolution du vivant