Endophyte
Les endophytes[1] (du grec endo « dans », φυτόν « végétal » ; littéralement « à l'intérieur d'un végétal », terme défini et employé pour la première fois en 1866 par Anton de Bary[2], mais Link est le premier à décrire en 1809 ces organismes sous le terme d'« Entophytae » considérés alors comme un groupe distinct de parasites fongiques[3]) sont tous les micro-organismes (bactéries ou champignons en général) qui accomplissent tout ou partie de leur cycle de vie à l'intérieur d'une plante, de manière symbiotique (endosymbiote), c'est-à-dire avec un bénéfice mutuel pour les deux organismes ou sans conséquences négatives pour la plante-hôte[4]. L'endophyte peut vivre et se reproduire dans les espaces intercellulaires (méats des parenchymes)[5] et/ou dans certaines cellules de la plante. L'endophyte et son hôte entretiennent une interaction appelée endophytisme. Les endophytes qui font partie du microbiote des plantes (le microbiome étant appelé phytobiome) jouent chez elles un rôle assez comparable à celui du microbiote intestinal chez l'animal, rôle qui selon les études les plus récentes, a été très longtemps inconnu puis sous-estimé.
De nombreux endophytes ont une activité ou des propriétés favorisant potentiellement la croissance de la plante, ou sa résilience face à certain stress (chaleur, manque d'eau, sel, attaques parasitaires via l'effet bioinsecticide, etc.) [6],[7]. Une partie du génome d'endophyte est parfois retrouvée, intégrée, dans l'ADN de la plante-hôte[8]
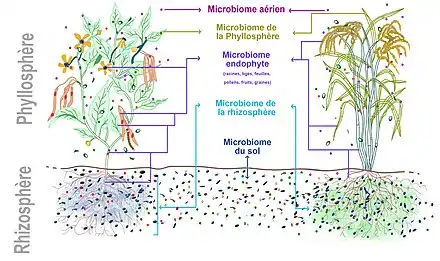
On retrouve aussi sur (voire dans) la plante des microbes plus ou moins ubiquistes et opportunistes, éventuellement pathogènes provenant de l'air et du sol.
Histoire
On connait depuis plusieurs décennies les bactéries symbiotes qui vivent dans les nodules de plantes fixatrices d'azote, mais la plupart des endophytes connus n'ont été découverts que récemment. Discrets car ne provoquant aucun symptôme de maladie, ils sont en réalité omniprésents ; retrouvés dans presque toutes les espèces de plantes étudiées à ce jour. On a d'abord cru qu'il s'agissait de champignons et/ou de bactéries pathogènes en latence dans la plante, sans y causer de dommages apparents, mais sans être bénéfique pour la plante Puis, au début des années 2000, essentiellement de nombreuses preuves scientifiques ont montré que ces endophytes concourent à améliorer la croissance et la santé des plantes (y compris cultivées), et à augmenter leur biomasse, notamment en les rendant plus résistantes aux maladies. L'endophytisme microbien est ainsi aujourd’hui considéré comme une règle générale chez les plantes[9].
Enjeux
Une partie des relations endophyte ↔ plante et de l'origine de ces relations sont encore mal comprises, mais on sait déjà que des endophytes jouent un rôle majeur pour la santé des plantes et leur bioproductivité, dont chez la plupart des plantes fourragères et herbacées économiquement importantes (par exemple, Festuca spp. ou Lolium spp.) vivent avec des champignons endophytes (Neotyphodium spp.) qui améliore notamment la capacité de ces plantes à tolérer des stress abiotiques tels que la sécheresse, ainsi que leur résistance aux insectes et mammifères herbivores[10]. Ou parmi les plantes engazonnantes, la fétuque élevée, le ray-grass vivace, la fétuque rude, la fétuque rouge gazonnante et la fétuque rouge traçante en contiennent, et ce dans tous les compartiments de la plante (hormis dans les racines qui elles vivent en symbiose avec d'autres microbes) ; les endophytes sont plus concentrés dans la gaine foliaire chez ces espèces, alors que le pâturin des prés ou l'agrostide stolonifère ne semble pas en contenir[11].
Une part des endophytes fait partie de la diversité biologique utile qui a été dégradée ou perdue par l'industrie semencière. Les semences paysannes et le patrimoine génétique endophyte qu'elles recèlent pourrait redevenir une ressource reconnue pour l'amélioration végétale dans le monde[12]
Il est possible que les pollinisateurs et/ou le type de pollinisation joue un rôle important dans la diversité biologique et donc dans l'évolution adaptative de l'endosphère[13]. Le recul des pollinisateurs serait alors aussi un enjeu encore mal évalué de ce point de vue. Selon des travaux récents, les espèces pollinisées par les insectes ont un microbiote moins diversifié que celles qui sont anémophiles (« ce qui suggère un effet de nivellement par les insectes vecteurs ; La microscopie électronique à balayage ainsi que l'hybridation fluorescente in situ couplée à la microscopie confocale à balayage laser (FISH ‐ CLSM) ont indiqué que la surface du tectum constituait la niche préférée de la colonisation bactérienne ».
Il est possible que leurs endophytes expliquent le succès de reproduction et d'adaptation ou de résilience de certaines plantes invasives[14].
Champignons endophytes
Acceptés par le système immunitaire de leur plante-hôte - probablement à la suite d'une longue coévolution - les champignons endophytes colonisent l'intérieur des tissus vivants de tout ou partie de cette plante, sans symptôme apparents de maladie[15].
Selon (Manoharachary et al. (2005)[16] il pourrait exister environ 1.5 million d’espèces de champignons endophytes, mais seules environ 75.000 d’entre elles avaient déjà été décrites en 2005 et le nombre d'espèces de champignon endophyte est encore complètement inconnu[14]. La plante nourrit et protège ces champignons. En échange, ces derniers apportent des métabolites intéressant pour la plante, car améliorant sa santé, sa compétitivité dans le milieu (par une résistance accrue aux pathogènes, aux parasites, à divers autres stress et en rendant la plante plus résiliente[17].
Transmission, reproduction & diffusion
Les modes de transmission des endophytes sont dits[18] :
- verticaux (passage direct des plantes « parents » aux propagules « enfants ») ; La transmission verticale des champignons endophytes correspond à une reproduction asexuée.
Elle se fait par des hyphes de champignon pénétrant l'hôte (exemple avecNeotyphodium). Leur taux de réussite en matière de reproduction est intimement lié à celui de leur plante hôte, ces champignons sont donc souvent ceux qui vivent en situation de mutualisme avec leur hôte. De même, les graines des arbres et des céréales, lorsqu'elles sont diffusées par le vent, l'eau, les oiseaux, mammifères, etc. hébergent déjà un complexe microbiotique de bactéries spécifiques. Ces bactéries proviennent de l'anthosphère, de la carposphère ou des cônes de gymnospermes mais aussi des tissus internes des plantes ; après une longue colonisation passant généralement du sol ou de l'eau du sol aux organes reproducteurs[15]. Les mises en cultures artificielles sur des substrats stérilisés peuvent priver la future plante d'une partie de ce patrimoine.
- horizontaux (passage d'un individu à un autre individu sans nécessité de contact ou lien direct).
La transmission horizontale est propre aux champignons endophytes à reproduction sexuée. Elle se fait via des spores qui peuvent être disséminées par le vent, ou par un vecteur (insectes en général) et pénètrent indirectement (via les stomates ou les lenticelles) ou directement. Ces champignons se propagent plutôt à la manière d'agents pathogènes infectieux (ils sont souvent apparentés à des agents phytopathogènes, mais n'en sont pas eux-mêmes, en tous cas pour leurs plantes-hôtes).
Relation des endophytes-hôte
Elles sont diverses et varient selon les plantes et les contextes :
- Des compatibilités génétiques sont nécessaires pour qu'un endophyte soit accepté par une plante[19]
- Certains endophytes sont très bénéfiques, voire nécessaires à leur plantes-hôtes. Ils améliorent l'accès de la plante aux nutriments et parfois empêchent certains organismes pathogènes de les coloniser (dans le cas de relations symbiotiques vraies) grâce à une batterie de molécules utiles ou défensives sécrétées par l'endophyte[20],[21].
Ainsi, la variété Dichanthelium lanuginosum var. thermale, qui pousse dans des domaines géothermiques du parc national de Yellowstone (États-Unis) est capable de résister à des températures élevées grace à une symbiose entre un champignon Curvularia protuberata (en) et un virus[22]. La vigne (Vitis vinifera L.)profite d'une colonisation par diverses souches de bacterium Burkholderia sp qui la rendent plus productive [23] - La colonisation de tout ou partie des tissus végétaux par des endophytes crée et entretient un «effet barrière» empêchant d'autres endophytes qui pourrait être pathogènes de s'installer, faute de place et/ou parce que les endophytes peuvent aussi sécréter des substances biocides ou répulsives à l'égard de ses concurrents, éventuellement pathogènes. L'utilisation par une plante de champignons endophytes pour se défendre est un phénomène très commun.
- Dans le cas des champinons endophytes, leur pénétration dans la plante se limite souvent aux seuls tissus racinaires, mais chez certaines espèces de plantes elle s'étend à d'autres organes (ex : « endophytes foliaires ») ; Dans tous les cas, la présence de champignons endophytes se traduit par une évapotranspiration plus importante de la plante hôte, mais des champignons mycorhiziens symbiotes aident aussi la plante à fortement augmenter son accès à l'eau du sol et aux nutriments ;
- Certains endophytes bénéfiques peuvent devenir pathogènes quand la plante, par exemple en situation de stress, ne les contrôle plus. Il semble en réalité exister une complexité probablement sous-estimée des relations d'endophytisme et d'interactions durables, avec un « continuum endophyte pathogène-saprotrophe »[24]. Ainsi des endophytes peuvent être bénéfiques dans certains écosystèmes, néfastes dans d'autres (par exemple l'intoxication par la fétuque élevée).
- Certains endophytes pathogènes passent directement de la plante à la graine (via l' infection du pollen ou du pistil au moment de la floraison[25]) ; la graine devient alors vectrice de la maladie[26],[27] et peuvent ainsi être diffusés (dont par l'homme lors du commerce à grande échelle de graines, qui peut donc contribuer à l'émergence de phytopathologies loin de leur source biogéographique.
Utilisation
Une large gamme de molécules complexes produits par des endophytes a été découverte et est en cours d'exploration, souvent avec des vertus antibiotiques, et parfois antiparasitaires voire anticancéreuses dans quelques cas quand ils sont utilisés chez les animaux, dont les humains.
Un endophyte notable du point de vue de l'intérêt médicinal a été découvert par le Dr Gary A. Strobel. C'est Pestalotiopsis microspora, un champignon endophyte vivant à l'intérieur de l'if Taxus wallachiana de l'Himalaya, qui produit le taxol [28].
Des endophytes (ou les molécules qu'ils produisent) pourraient être utilisés en agriculture, sylviculture ou production de bio- ou agrocarburants.
Inoculer des plantes cultivées avec certains endophytes peut en effet améliorer la résistance ou résilience de ces plantes face à divers stress ou maladies[10] ou parasites.
D'autres endophytes disposent d'enzymes capables de convertir la cellulose et d'autres sources de carbone, enzymes que certains aimeraient utiliser par exemple via le génie génétique ou d'autres manipulations (inoculations) par exemple pour produire un carburant "myco-diesel»[29].
Piriformospora indica est un champignon endophyte de l'ordre Sebacinales semble pouvoir coloniser les racines et entretenir une relation symbiotique avec n'importe quelle plante terrestre. P. indica s'est aussi montré capable d'augmenter le rendement des récoltes et les défenses de plusieurs variété cultivées (d'orge, tomates, maïs, etc) contre certains des agents pathogènes des racines[30],[31].
Au vu des données disponibles, on peut supposer qu'il puisse exister plusieurs milliers d'endophytes potentiellement utiles à l'humanité. Il n'y a cependant que peu de spécialistes pour les étudier et les forêts et milieux riches en biodiversité sont de plus en plus rapidement détruits ou dégradés et à grande échelle, rendant probable la disparition de nombreux endophytes potentiellement utiles pour la médecine, l'industrie, l'économie... avant même qu'ils ne soient découverts.
Quelques études portent sur les effets réels ou possibles du dérèglement climatique ou d'une augmentation des taux de CO2 sur les endophytes (dans le domaine de la forêt et de l'agriculture essentiellement).
Identification
Elle se fait via le microscope, les analyses biologiques ou génétiques.
Beaucoup d'endophytes colonisent de multiples espèces de plantes. Certains ont un ou quelques hôtes spécifiques. On suppose qu'ils ont coévolué avec leurs hôtes et son système immunitaire[32].
Les espèces endophytes déjà connues sont très diverses, et on n'en a probablement identifié, caractérisé et nommé qu'une petite partie. Une seule et même feuille peut abriter de nombreuses espèces d'endophytes, tant bactériennes que fongiques. Ils peuvent être identifiés de plusieurs manières, de plus en plus souvent par amplification (PCR) et séquençage d'un petit morceau d'ADN. Certains endophytes peuvent être mis en culture à partir d'un petit morceau de leur plante-hôte, dans un milieu de croissance approprié.
Il existe des espèces cryptiques d'endophytes.
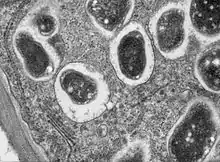
À titre d'exemple, on peut observer des endophytes dans les feuilles des herbacées les plus communes au microscope (grossissement 400 X). Ils ont l'apparence de tubes en spirale (hyphes) une fois les cellules foliaires traitées à l'éthanol et colorées au bleu d'aniline.
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Endophyte » (voir la liste des auteurs).
- Nota bene : Endophyte est masculin
- (de) Morphologie und Physiologie der Pilze, Flechten und Myxomyceten, 1866
- (de) Claudia Görke, Mykozönosen von Wurzel und Stamm von Jungbäumen unterschiedlicher Bestandsbegründungen, Borntraeger, , p. 376
- Hyde K. D. and Soytong K (2008) The fungal endophyte dilemma. Fungal Diversity ; 33:163-173
- Sandrine Gelin, Pierre Stengel, Sol : interface fragile, Quae, , p. 105
- Alibrandi P, Cardinale M, Rahman MM, Strati F, Ciná P, de Viana ML, Giamminola EM, Gallo G, Schnell S, de Filippo C, Ciaccio M, Puglia AM (2017) The seed endosphere of Anadenanthera colubrina is inhabited by a complex microbiota, including Methylobacterium spp. and Staphylococcus spp. with potential plant-growth promoting activities (L'endosphère de la graine d'Anadenanthera colubrina est habitée par un microbiote complexe, notamment Methylobacterium spp. et Staphylococcus spp. avec des activités potentielles favorisant la croissance des plantes). Plant Soil 1–19. doi: 10.1007/s11104-017-3182-4 (résumé)
- Rodriguez R.J, Redman R.S & Henson J.M (2004) The role of fungal symbioses in the adaptation of plants to high stress environments. Migration and Adaptation Strategies for Global Change ;9: 261-272
- Saar D. E., Polans N.O., Sorensen P. D. and Duvall M. R.Angiosperm (2001) DNA contamination by endophytic fungi: Detection and methods of avoidance. Plant Molecular Biology Reporter ; 19: 249-260.
- (en) George Carroll, « Fungal Endophytes in Stems and Leaves. From Latent Pathogen to Mutualistic Symbiont », Ecology, vol. 69, no 1, , p. 2-9 (DOI 10.2307/1943154)
- (en) « University of Rhode Island GreenShare Factsheets: Endopyhte-Enhanced Grasses » (consulté le )
- Omagra, Ontario (ministère de l'Agriculture) Démystifier les endophytes, consulté 2012-01-07
- Chable V (2018) Regain des semences paysannes. Pour, (2), 63-72.
- Ambika Manirajan B, Ratering S, Rusch V, Schwiertz A, Geissler-Plaum R, Cardinale M, Schnell S (2016) Bacterial microbiota associated with flower pollen is influenced by pollination type, and shows a high degree of diversity and species-specificity. Environ Microbiol 18:5161–5174. doi: 10.1111/1462-2920.13524
- Shipunov, A., Newcombe, G., Raghavendra, A. K., & Anderson, C. L. (2008). Hidden diversity of endophytic fungi in an invasive plant. American Journal of Botany, 95(9), 1096-1108.
- Zerroug A (2018) Métabolites secondaires bioactifs des champignons endophytes isolés de retama raetam Forssk (Doctoral dissertation).
- Manoharachary C., Sridhar K., Singh R., Adholeya A., Suryanarayanan T. S., Rawat S. and Johri N (2005) Fungal biodiversity: Distribution, conservation and prospecting of fungi from India. Current Science ; 89: 58-71
- Saikkonen K, Faeth S.H, Helander M and Sullivan T.J (1998) Fungal endophytes: A continuum of interactions with host plants. Annual Review of Ecology and Systematics; 29: 319-343.
- (en) Vijay C. Verma, Alan C. Gange, Advances in Endophytic Research, Springer Science & Business Media, (lire en ligne), p. 70
- Saikkonen K., Wali P. R. and Helander M (2010) Genetic compatibility determines endophyte-grass combinations | PLoS One ; 5(6):e11395. doi:10.1371/journal.pone.0011395
- Tan, R. X., & Zou, W. X. (2001). Endophytes: a rich source of functional metabolites. Natural Product Reports, 18(4), 448-459.
- Compant S, Clément C, Sessitsch A (2010) Plant growth-promoting bacteria in the rhizo- and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol Biochem 42:669–678. doi: 10.1016/j.soilbio.2009.11.024
- (en) Luis Márquez, Regina Redman, Russell Rodriguez et Marilyn J. Roossinck, « A Virus in a Fungus in a Plant: Three-Way Symbiosis Required for Thermal Tolerance », Science, vol. 315, no 5811, , p. 513–5 (DOI 10.1126/science.1136237, lire en ligne).
- Compant S, Reiter B, Sessitsch A, Nowak J, Clément C, Ait Barka E (2005) Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl Environ Microbiol 71:1685–1693. doi: 10.1128/aem.71.4.1685-1693.2005
- Arnold AE. 2007. Understanding the diversity of foliar endophytic fungi: progress, challenges, and frontiers. Fungal Biol. Rev. 21: 51-66
- Maude R.B (1996) Seedborne diseases and their control: principles and practice. CAB International (Wallingford, UK)
- Darrasse, A., Bureau, C., Samson, R., Morris, C.E., Jacques, M.A. (2007). Contamination of bean seeds by Xanthomonas axonopodis pv. phaseoli associated with low bacterial densities in the phyllosphere under field and greenhouse conditions. European Journal of Plant Patholog
- Darrasse, A., Darsonval, A., Boureau, T., Brisset, M.-N., Durand, K. and Jacques, M.-A. (2010) Transmission of Plant-Pathogenic Bacteria by Nonhost Seeds without Induction of an Associated Defense Reaction at Emergence. Applied and Environmental Microbiology,76, 6787-6796
- (en) Gary Strobel, « Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana », Microbiology, no 142, , p. 435–440
- Strobel G et al. Microbiology 154 (2008), 3319-3328; DOI 10.1099/mic.0.2008/022186-0
- (en) Ajit Varma, « Piriformospora indica, a Cultivable Plant-Growth-Promoting Root Endophyte », Applied and Environmental Microbiology, vol. 65, no 6, , p. 2741–2744 (lire en ligne)
- (en) Karl-Heinz Kogel, « The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield », Proc Natl Acad Sci(PNAS), vol. 102, no 38, , p. 13386–13391 (PMID 16174735, PMCID 1224632, DOI 10.1073/pnas.0504423102)
- Saikkonen K., Wali P., Helander M. & Faeth S. H (2004) Evolution of endophyte-plant symbioses. Trends in Plant Science 9: 275-280
Voir aussi
Articles connexes
Liens externes
- (en) « Endophytes and their promise for new medicines and products for agriculture and industry », Jewels of the Jungle (consulté le )
- Microbewiki Plant_endophyte
Bibliographie
- (en) Barret, M., Guimbaud, J. F., Darrasse, A., & Jacques, M. A. (2016) Plant microbiota affects seed transmission of phytopathogenic micro-organisms 4. Links, 2920, 238.
- (en) Cheplick, G.P. and S.H. Faeth. 2009. Ecology and Evolution of the Grass-Endophyte Symbiosis. Oxford University Press, Oxford.
- (en) Compant S, Clément C, Sessitsch A (2010) Plant growth-promoting bacteria in the rhizo- and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol Biochem 42:669–678. doi: 10.1016/j.soilbio.2009.11.024
- (en) Compant S, Mitter B, Colli-Mull JG, Gangl H, Sessitsch A (2011) Endophytes of grapevine flowers, berries, and seeds: identification of cultivable bacteria, comparison with other plant parts, and visualization of niches of colonization. Microb Ecol 62:188–197. doi: 10.1007/s00248-011-9883-y
- (en) Edwards J, Johnson C, Santos-Medellín C, Lurie E, Podishetty NK, Bhatnagar S, Eisen JA, Sundaresan V (2015) Structure, variation, and assembly of the root-associated microbiomes of rice. Proc Natl Acad Sci U S A 112:911–920. doi: 10.1073/pnas.1414592112
- (en) Ewald PW (1987) Transmission modes and evolution of the parasitism-mutualism continuum. Ann N Y Acad Sci 503:295–306
- (en) Ferreira A, Quecine MC, Lacava PT, Oda S, Azevedo JL, Araújo WL (2008) Diversity of endophytic bacteria from eucalyptus species seeds and colonization of seedlings by Pantoea agglomerans. FEMS Microbiol Lett 287:8–14. doi: 10.1111/j.1574-6968.2008.01258.x
- (en) Fürnkranz M, Lukesch B, Müller H, Huss H, Grube M, Berg G (2012) Microbial diversity inside pumpkins: microhabitat-specific communities display a high antagonistic potential against phytopathogens. Microb Ecol 63:418–428. doi: 10.1007/s00248-011-9942-4
- (en) Glassner H, Zchori-Fein E, Yaron S, Sessitsch A, Sauer U, Compant S (2017) Bacterial niches inside seeds of Cucumis melo L. Plant Soil:1–13. doi: 10.1007/s11104-017-3175-3
- (en) Hardoim PR, Hardoim CCP, van Overbeek LS, van Elsas JD (2012) Dynamics of seed-borne rice endophytes on early plant growth stages. PLoS One 7:e30438. doi: 10.1371/journal.pone.0030438
- (en) Hardoim PR, van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A, Döring M, Sessitsch A (2015) The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol Mol Biol Rev 79:293–320. doi: 10.1128/MMBR.00050-14
- (en) James EK, Gyaneshwar P, Mathan N, Barraquio WL, Reddy PM, Iannetta PP, Olivares FL, Ladha JK (2002) Infection and colonization of rice seedlings by the plant growth-promoting bacterium Herbaspirillum seropedicae Z67. Mol Plant-Microbe Interact 15:894–906. doi: 10.1094/MPMI.2002.15.9.894
- (en) Johnston-Monje D, Raizada MN (2011) Conservation and diversity of seed associated endophytes in Zea across boundaries of evolution, ethnography and ecology. PLoS One 6:e20396. doi: 10.1371/journal.pone.0020396
- (en) Klaedtke S, Jacques M-A, Raggi L, Préveaux A, Bonneau S, Negri V, Chable V, Barret M (2016) Terroir is a key driver of seed-associated microbial assemblages. Environ Microbiol 18:1792–1804. doi: 10.1111/1462-2920.12977
- (en) Lòpez-Fernàndez S, Mazzoni V, Pedrazzoli F, Pertot I, Campisano A (2017) A phloem-feeding insect transfers bacterial endophytic communities between grapevine plants. Front Microbiol 15; 8:834. 10.3389/fmicb.2017.00834
- (en) Madmony A, Chernin L, Pleban S, Peleg E, Riov E (2005) Enterobacter cloacae, an obligatory endophyte of pollen grains of Mediterranean pines. Folia Microbiol (Praha) 50:209–216
- (en) Maude RB (1996) Seedborne diseases and their control: principles and practice. CAB International, Wallingford
- (en) Mercado-Blanco J, Lugtenberg BJJ (2014) Biotechnological applications of bacterial endophytes. Curr Biotechnol 3:60–75
- (en) Miller IM (1990) Bacterial leaf nodule symbiosis. Adv Bot Res Adv Plant Pathol 17:163–234
- (en) Mitter B, Pfaffenbichler N, Flavell R, Compant S, Antonielli L, Petric A, Berninger T, Muhammad N, Sheibani-Tezerji R, Von Maltzahn G, Sessitsch A (2017) A new approach to modify plant microbiomes and traits by introducing beneficial bacteria at flowering into progeny seeds. Front Microbiol. doi: 10.3389/fmicb.2017.00011
- (en) Philippot L, Raaijmakers JM, Lemanceau P, van der Putten WH (2013) Going back to the roots: the microbial ecology of the rhizosphere. Nat Rev Microbiol 11:789–799
- (en) Podolich O, Ardanov P, Zaets I, Pirttilä A, Kozyrovska N (2015) Reviving of the endophytic bacterial community as a putative mechanism of plant resistance. Plant Soil 388(1–2):367–377
- (en) Pohjanen J, Koskimäki JJ, Pirttilä AM (2014) Interactions of meristem-associated endophytic bacteria. In: Verma VC, Gange AC (eds) Advances in endophytic research. New Delhi, Springer India, pp 103–113C
- (en) Prévost P, Lallemand P (2010) L’approche terroir : pour une démarche de recherche-formation-action http://hal.archives-ouvertes.fr/docs/00/51/25/28/PDF/Prevost_approche-terroir.pdf
- (en) Rezki S, Campion C, Iacomi-Vasilescu B, Preveaux A, Toualbia Y, Bonneau S, Briand M, Laurent E, Hunault G, Simoneau P, Jacques M-A, Barret M (2016) Differences in stability of seed-associated microbial assemblages in response to invasion by phytopathogenic microorganisms. PeerJ 4:e1923
- (en) Robinson RJ, Fraaije BA, Clark IM, Jackson RW, Hirsch PR, Mauchline TH (2016) Wheat seed embryo excision enables the creation of axenic seedlings and Koch’s postulates testing of putative bacterial endophytes. Sci Rep 6:25581
- (en) Rosenblueth M, López-López A, Martínez J, Rogel MA, Toledo I, Martínez-Romero E (2012) Seed bacterial endophytes: common genera, seed-to-seed variability and their possible role in plants. Acta Hort (ISHS) 938:39–48
- (en) Schulz B. and Boyle C (2005) The endophytic continuum. Mycological Research ; 109: 661-686.
- (en) Shade A, McManus PS, Handelsman J (2013) Unexpected diversity during community succession in the apple flower microbiome. MBio 4:e00602–e00612. doi: 10.1128/mBio.00602-12
- (en) Smith EF (1911) Bacteria in relation to plant diseases. Carnegie institution of Washington, Washington, D.C.
- (en) Truyens S, Weyens N, Cuypers A, Vangronsveld J (2015) Bacterial seed endophytes: genera, vertical transmission and interaction with plants. Environ Microbiol Rep 7:40–50. doi: 10.1111/1758-2229.12181
- (en) Ushio M, Yamasaki E, Takasu H, Nagano A, Fujinaga S, Honjo MN, Ikemoto M, Sakai S, Kudoh H (2015) Microbial communities on flower surfaces act as signatures of pollinator visitation. Sci Rep 5:8695. doi: 10.1038/srep08695
- (en) Van Oevelen S, De Wachter R, Vandamme P, Robbrecht E, Prinsen E (2002) Identification of the bacterial endosymbionts in leaf galls of Psychotria (Rubiaceae, angiosperms) and proposal of ‘Candidatus Burkholderia kirkii’ sp. nov. Int J Syst Evol Microbiol 52:2023–2027
- (en) Van Oevelen S, De Wachter R, Vandamme P, Robbrecht E, Prinsen E (2004) ‘Candidatus Burkholderia calva’ and ‘Candidatus Burkholderia nigropunctata’ as leaf gall endosymbionts of African Psychotria. Int J Syst Evol Microbiol 54:2237–2239
- (en) von Faber FC (1912) Das erbliche Zusammenleben von Bakterien und tropischen Pflanzen. Jahrb Wiss Bot 54:285–375
- (en) Rosenblueth, M., & Martinez-Romero, E. (2006). Bacterial endophytes and their interactions with hosts. Molecular Plant-Microbe Interactions, 19(8), 827-837 (Résumé).
- (en) Strobel G et al. Microbiology 154 (2008), 3319-3328; DOI 10.1099/mic.0.2008/022186-0
 Portail de la botanique
Portail de la botanique  Portail de la mycologie
Portail de la mycologie  Portail de la protection des cultures
Portail de la protection des cultures